










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.61 no.3 Palmira July/Sept. 2012
1 Centro Internacional de Agricultura Tropical (CIAT), A.A. 6713, Cali, Valle del Cauca, Colombia.
2 Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, A.A. 237, Palmira, Valle del Cauca, Colombia.
*Autor para correspondencia: j.a.polania@cgiar.org
El desarrollo de variedades adaptadas es una de las estrategias que contribuye a garantizar la seguridad alimentaria en zonas productoras de frijol con estrés por sequía. En los invernaderos de cristal del Centro Internacional de Agricultura Tropical (CIAT), Palmira, Colombia, se evaluaron las características morfológicas y fisiológicas de raíces de 21 líneas de frijol (Phaseolus vulgaris L.) en condiciones de estrés por sequía e irrigación, utilizando un sistema de tubos plásticos desarrollado por el CIAT. Las características evaluadas fueron profundización visual, longitud total de raíces y distribución de raíces a través del perfil del suelo. En campo, utilizando los mismos genotipos de frijol, se evaluaron características relacionadas con la eficiencia de uso de agua mediante la técnica de discriminación de isótopo de carbono y producción de grano. Los resultados en invernadero mostraron que la profundización visual y longitud de raíces entre 60 y 75 cm tienen una correlación significativa (r = 0.56 y 0.50 respectivamente, P < 0.001) con la biomasa aérea de la planta en condiciones de estrés por sequía. En campo, la discriminación del isótopo de carbono en grano se correlacionó (r = 0.32, P < 0.01) con la producción de grano en condiciones igualmente de estrés por sequía. Las líneas de frijol NCB 226, SER 16, SEN 56 y SEA 15 presentaron una mayor longitud de raíces bajo estrés por sequía (28, 24, 29 y 27 m/planta respectivamente) en comparación con las demás líneas evaluadas, lo que les permite mayor transpiración y movilización de fotoasimilados, que favorecen una alta producción de granos. Se estima que la profundización visual, la longitud de raíces entre 60 y 75 cm y la discriminación de isótopo de carbono en grano, son características valiosas como criterios de selección en mejoramiento por tolerancia a estrés por sequía en frijol común.
Palabra clave: Discriminación de isótopo de carbono, longitud de raíz, producción de grano, profundización de raíz.
Development of drought adapted common bean cultivars is a strategy to improve food security in bean producing areas under drought stress. Under greenhouse conditions at the International Center for Tropical Agriculture (CIAT), Palmira, Colombia, we evaluated the morphological and physiological traits of 21 common bean lines (Phaseolus vulgaris L.) that were subjected to drought stress and irrigated treatments using a soil cylinder system. The characteristics evaluated were visual rooting depth, total root length and distribution across the soil profile. In the field, using the same bean genotypes, we evaluated differences in water use efficiency (using carbon isotope discrimination) and grain yield. Under greenhouse conditions, we found that visual rooting depth and root length at soil depth of 60-75 cm correlated positively with shoot biomass (r = 0.56 and 0.50 respectively, P < 0.001) production when water supply was limiting. Under field conditions, grain yield under drought stress was positively associated with carbon isotope discrimination in grain (r = 0.32 P < 0.01). Five lines (NCB 226, SER 16, SEN 56 and SEA 15) had higher root length under drought stress (28, 24, 29 and 27 m/plant respectively) compared with the other lines evaluated, which allows greater water uptake and transpiration, and also in mobilization of photosynthate to greater production of grain. It is estimated that the visual rooting depth, root length at soil depth of 60 to 75 cm and carbon isotope discrimination in grain, are valuable traits as selection criteria in breeding for drought stress tolerance in common bean.
Key words: Carbon isotope discrimination, grain yield, root length, visual rooting depth.
El cambio climático es una amenaza progresiva y cada vez más latente para la producción de alimentos, especialmente en las regiones menos desarrolladas. Entre estas amenazas se encuentran sequías e inundaciones severas y frecuentes que favorecen la aparición de nuevas plagas y enfermedades y el aumento de las ya existentes. Los impactos más negativos de esta situación los sentirán los pequeños agricultores en sistemas de subsistencia en países en desarrollo (Morton, 2007). El frijol común (Phaseolus vulgaris L.) es la leguminosa más importante para la alimentación en el trópico de América Latina y Africa oriental y meridional. El frijol es actualmente cultivado en estas regiones por pequeños agricultores, con frecuencia en condiciones no favorables y de mínimo uso de insumos (Beebe et al., 2008), donde constituye una fuente económica de proteína y calorías para la población (Rao, 2001). No obstante los rendimientos del cultivo son afectados por varias causas, entre ellas la sequía que en ocasiones genera pérdidas entre 10% y 100% del cultivo. Cerca del 60% de las regiones productoras de frijol presentan periodos prolongados de sequía y es el segundo factor más importante de reducción en rendimiento, después de las enfermedades (Thung y Rao, 1999; Rao, 2001).
El desarrollo de cultivares adaptados a estas condiciones extremas es una opción posible para contribuir a la seguridad alimentaria de una población creciente que requiere incrementos significativos en la producción de alimentos (Polanía et al., 2009). Es un hecho cierto que el progreso en los programas de mejoramiento depende de la selección precisa de los genotipos poco frecuentes que poseen atributos nuevos o mejorados (Sorrells, 2007), lo cual significa que una caracterización fenotípica adecuada seguirá siendo uno de los pilares del mejoramiento. Existen algunos procesos que pueden ser utilizados para el mejoramiento por estrés hídrico de plantas, entre ellos: (1) mayor movimiento del agua disponible en la capa arable por la acción radicular; (2) adquisición de más carbono (biomasa) a cambio del agua transpirada por el cultivo, o eficiencia de transpiración; y (3) mayor movilización de la biomasa acumulada al producto cultivado (Condon et al., 2004).
Turner (1979) y Huang y Gao (2000) encontraron que algunas características de la planta mejoran la adaptación a condiciones de sequía, especialmente un sistema radicular vigoroso. Un mayor acceso a agua disponible garantiza altas tasas de transpiración y crecimiento.
La discriminación de isótopo de carbono es una característica que permite diferenciar por tasa de transpiración y eficiencia de uso de agua. Los isótopos son variantes de los átomos de un elemento químico; en el caso del carbono existen tres isótopos: 12C, 13C y 14C, cuyos núcleos contienen el mismo número de protones (seis), pero un número diferente de neutrones (seis, siete y ocho, respectivamente), por tanto a pesar de tener propiedades químicas semejantes, tienen una masa atómica diferente: 12, 13 y 14 (Uriarte, 2003). La mayoría del carbono natural es 12C y representa el 98.9% en la naturaleza, una pequeña parte es 13C (1.1%) (Farquhar et al., 1989). Una porción aún más pequeña es 14C, que es radiactivo e inestable, y su aplicación ha sido paleo-cronológica. Durante la fotosíntesis, las plantas realizan una discriminación prefiriendo la molécula de CO2 con el carbono más liviano 12C, por tanto la mayor proporción de carbono contenido en la biomasa vegetal es de este tipo (Farquhar et al., 1989). La discriminación de isótopo de carbono se utiliza en fisiología vegetal como indicador indirecto del cierre de estomas y uso eficiente de agua, debido a que durante periodos de estrés hídrico algunas plantas tienden a cerrar sus estomas para perder menos agua y en consecuencia a menor CO2 entrante, las plantas discriminan menos el 13C y su concentración en los azúcares aumenta (Uriarte, 2003).
La selección por baja discriminación de 13C ha sido propuesta como un método para mejorar el uso eficiente de agua en programas de mejoramiento de cultivos C3 (Khan et al., 2007). En el caso de frijol común (cultivo C3), varios genotipos evaluados en condiciones de estrés hídrico mostraron una mayor discriminación de 13C, igualmente se encontró una relación positiva entre este tipo de discriminación y la densidad de longitud de raíces (White et al., 1990). A partir de los resultados de estos estudios se generó la hipótesis que las plantas bajo estrés hídrico con capacidad para producir raíces profundas y, por tanto, acceder a un mayor volumen de agua, sufren menos por este tipo de estrés, lo que resulta en una mayor conductancia estomática y mayor discriminación de 13C; no obstante es una hipótesis que aún requiere estudios más detallados (Hall 2004; White et al., 1990; White 1993).
El objetivo principal de este trabajo fue evaluar características morfo-fisiológicas del cultivo de frijol común, como enraizamiento profundo y discriminación de isótopo de carbono y su contribución para una mayor producción de grano bajo estrés por sequía, con el fin de incorporarlas como criterios de selección en el programa de mejoramiento del cultivo.
La investigación se realizó en el Centro Internacional de Agricultura Tropical (CIAT) situado en Palmira, Valle del Cauca, Colombia, a 3° 31' N y 76° 19' W, a 965 m.s.n.m. Se utilizó un grupo de 21 genotipos meso-americanos de frijol común (P. vulgaris L.): BAT 477, Carioca, DOR 390, NCB 226, NCB 280, RCB 273, SEA 15, SEN 36, SEN 56, SER 109, SER 113, SER 118, SER 125, SER 16, SER 78, SXB 405, SXB 412, SXB 415, SXB 418 y Tío Canela 75 y G 40001 (Phaseolus acutifolius). Para evaluar las características relacionadas con enraizamiento profundo se realizó un ensayo en condiciones de invernadero y para las características de discriminación de isótopo de carbono y producción de grano se hizo un ensayo en campo.
En invernadero. El ensayo se desarrolló entre febrero y mayo del 2010. Las plantas fueron establecidas en cilindros de plástico transparente de 8 cm de diámetro y 80 cm de profundidad, insertados en tubos de PVC, en un Oxisol de Santander de Quilichao, Colombia, sin presencia de toxicidad por aluminio (pH 4.6, Saturación Al < 35%) ni compactación (densidad aparente de 1.1 g/cc). El suelo utilizado en el ensayo no fue esterilizado ni recibió aplicación de inoculantes, pero sí fue fertilizado con una dosis equivalente (kg/ha) de 80 N, 50 P, 100 K, 101 Ca, 29.4 Mg, 20 S, 2 Zn, 2 Cu, 0.1 B y 0.1 Mo. Los tratamientos se dispusieron en bloques completos al azar, con dos niveles de suministro de agua: 80% capacidad de campo (CC) (riego) y sin riego (sequía terminal) en tres repeticiones. Las plantas se desarrollaron en condiciones óptimas de riego (80% CC) durante los primeros 10 días y después de este periodo de establecimiento fueron sometidas a los tratamientos de suministro de agua. En el tratamiento de riego los cilindros fueron pesados cada dos días y se aplicó una cantidad de agua equivalente al consumo. En los del tratamiento de sequía terminal no se aplicó riego, según la metodología propuesta por Butare et al. (2011) y Polanía et al. (2009). En las raíces se midieron: profundización visual medida con cinta métrica una vez por semana; biomasa aérea de la planta; y longitud total y distribución de raíces a través del perfil del suelo en cinco profundidades (0 - 10, 10 - 20, 20 - 40, 40 - 60 y 60 - 75 cm). Las raíces fueron separadas del suelo por lavado en forma manual, limpiadas y procesadas en un escáner Epson Expression 10000 XL. La longitud de raíz por cada perfil del suelo (m/ planta) fue determinada con el analizador de imágenes WinRHIZO versión 2007.
En campo. El trabajo se realizó en el CIAT entre junio y septiembre de 2009, en un Mollisol (Aquic Hapludoll) de alta fertilidad, con una capacidad estimada de almacenamiento de agua disponible de 130 mm hasta 1 m de profundidad en el suelo –agua entre CC -0.03Mpa y punto de marchitez permanente -15Mpa–. Se utilizó un diseño de Latice de 6 x 6 (36 genotipos, incluidas las 21 líneas del ensayo anterior y 15 líneas adicionales en observación), en tres repeticiones. Cada parcela estaba representada por cuatro surcos de 3.72 m de longitud, con una distancia entre surcos de 60 cm y un espaciamiento entre plantas de 7.5 cm (15 semillas por 1 m lineal del surco). Las plantas recibieron los tratamientos de riego y estrés por sequía. Para asegurar el establecimiento de estas últimas se aplicaron tres riegos por gravedad, de aproximadamente 35 mm cada uno, el primero tres días antes de la siembra, y los dos restantes diez y veintiún días después de la siembra. En el tratamiento de riego se aplicaron en total ocho riegos con un espaciamiento máximo entre ellos de diez días.
La producción de grano como peso seco y húmedo se midió mediante la cosecha de las plantas en 3 m de dos surcos centrales de cada parcela. En cada repetición se tomó una muestra de grano la cual se secó en horno a 60 °C durante 48 h antes de ser molidas y enviadas a la Universidad de California, Davis, para análisis de 13C/12C en espectrómetro de masas que permite medir la desviación d13C de la concentración isotópica en la muestra, con respecto a una medida estándar, que es el carbono presente en carbonato cálcico de la concha de un fósil marino denominado PDB (Pee Dee Belemnite) (Farquhar et al., 1989).
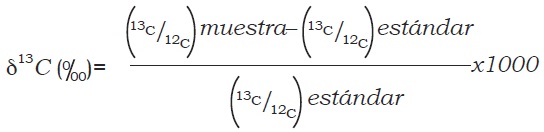
La discriminación de D 13C se calcula mediante la siguiente relación, que refleja la magnitud del fraccionamiento isotópico entre la fuente y el producto, siendo en este caso la fuente el aire, el cual tiene una composición isotópica estable de d13C de –8.
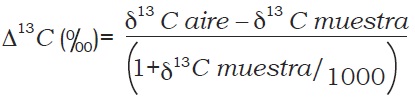
La discriminación de carbono, D, se expresa en unidades de mil (0/00), un equivalente a 10-3, que no es unidad; por tanto, la discriminación del isótopo de carbono es adimensional (Farquhar et al., 1989).
Con la ayuda del programa estadístico SAS, v 9.2 (SAS Institute, 2008) se calcularon correlaciones entre las diferentes características de las raíces y la biomasa foliar en invernadero, e igualmente entre discriminación de isótopo de carbono y rendimiento de grano. Los datos se sometieron a análisis de varianza y fueron graficados para observar la relación entre las diferentes características.
En invernadero. Durante el crecimiento y desarrollo del cultivo, se registraron temperaturas promedio máxima de 32.4 °C y mínima de 19.1 °C. El contenido de humedad al finalizar el ensayo (48 días después de la siembra) fue del 52% de la capacidad de campo. En promedio, la producción de biomasa de la parte aérea de la planta en condiciones de estrés por sequía fue de 1.67 g/planta, mientras que en condiciones óptimas de riego fue de 4.65 g/planta, lo que equivale a una reducción por estrés de 73% sobre la producción de biomasa aérea (tallo, hoja y vaina) de la planta, la cual indica la severidad y el daño por estrés debido a sequía terminal (secamiento progresivo del suelo). La longitud total de raíz en condiciones de sequía presentó una correlación positiva con la producción de biomasa aérea (P < 0.05) (Cuadro 1), lo que indica que a mayor producción de raíces bajo sequía, mayor es el vigor de la planta en términos de biomasa aérea. Trabajos previos en frijol común mostraron la existencia de relación entre las características aéreas de la planta y de la raíz y la importancia de esta última en la adaptación de frijol a sequía (White y Castillo, 1989). Por otra parte, también se observaron correlaciones positivas (P < 0.05) entre biomasa aérea y la profundización visual de las raíces en diferentes tiempos de desarrollo de la planta, tanto en condiciones óptimas de riego como de estrés por sequía (Cuadro 1).
El desarrollo radicular adecuado permite reducir y amortiguar los efectos de la deshidratación en las plantas, así, la profundización de la raíz les permite explorar un mayor volumen de suelo y extraer agua de perfiles inferiores, cuando las raíces superficiales ya han agotado el agua disponible en extractos superiores del suelo (Khan et al., 2010). Ho et al. (2005), Blum (2002), Huang y Gao (2000), Sponchiado et al. (1989), y Turner (1979) encontraron que un sistema radicular profundo y vigoroso es una característica importante de las plantas adaptadas a sequía prolongada. En el caso particular del frijol el desarrollo de raíces profundas es una característica para mejorar adaptación a sequía (Beebe et al., 2010; Polanía et al., 2009; Beebe et al., 2008; White et al., 1994; Sponchiado et al., 1989). Los genotipos que presentaron mayor profundidad de raíces, vigor de planta y desarrollo de la parte aérea fueron SER 109, SEA 15, SXB 412, SEN 56 y SER 16 (Figura 1 y Figura 2).
Los genotipos Tío Canela 75 y BAT 477 presentaron raíces profundas, pero su biomasa aérea no fue lo suficientemente desarrollada, lo que indica que la planta invierte una gran parte de la energía para la obtención de agua. Otros genotipos presentaron una baja profundización de raíces combinado con una baja producción de biomasa aérea (Figuras 1 y 2). Khan et al., (2010) y Bruce et al. (2002) consideran que los desbalances que ocasiona el estrés por sequía en las plantas pueden ser compensados por el mayor crecimiento radicular con un alto uso de foto-asimilados y una mayor absorción de agua.
En campo. Durante el crecimiento y desarrollo del cultivo se registraron, en promedio, una temperatura máxima de 32.5 °C y mínima de 19.41 °C. La precipitación total en el periodo de cultivo fue de 58.7 mm y la evaporación potencial de 462.5 mm, lo que indica que el cultivo creció en condiciones de sequía terminal. En promedio, la producción de grano en estas condiciones fue de 989 kg/ ha vs. 2516 kg/ha en condiciones óptimas de riego, equivalente a una reducción de 61% (Cuadro 2). El estrés por sequía aumentó la precocidad en la mayoría de las líneas evaluadas, ya que en estas condiciones se requirieron menos días para que la planta alcance la madurez fisiológica.
Rendimiento de grano. En ambos sistemas, riego y sequía, se observaron diferencias (P < 0.05) en producción de grano entre genotipos. La relación en rendimiento entre sistemas permite identificar las líneas mejor adaptadas a sequía y a la vez responden bien a la aplicación de riego, entre ellas: SEN 36, SEN 56, SER 16, SER 113 y NCB 226, lo que indica que la selección por tolerancia a sequía no necesariamente reduce el rendimiento en condiciones óptimas de cultivo y que el estrés hídrico no es excluyente para un buen comportamiento bajo riego. Las líneas SER 78, NCB 280, SEA 15 y SER 125 presentaron muy buena adaptación a condiciones de sequía, pero no respondieron a la aplicación de riego.
La discriminación del isótopo de carbono 13C –una variable que se relaciona con el uso eficiente de agua en plantas C3, como frijol– presentó una correlación significativa (r = 0.32**) con el rendimiento de grano en condiciones de sequía. A mayor discriminación de 13C menor es la eficiencia de uso de agua, por tanto esta correlación positiva indica que en frijol común los materiales de mayor productividad son aquellos que más transpiran y consecuentemente hacen un uso menos eficiente del agua. Estos resultados coinciden con los encontrados por White et al. (1990) quienes observaron una relación positiva entre frijol de alto rendimiento bajo estrés por sequía con una mayor discriminación de 13C.
Los genotipos NCB 226, SER 125, SXB 415, NCB 280, SER 16, SEN 36, SEN 56 y SEA 15 combinan alto rendimiento de grano con alta discriminación de 13C en grano en condiciones de sequía, por tanto, hacen uso menos eficiente de agua (Figura 3) y presentan una mayor transpiración, uso de agua e intercambio de gases para obtener mayor vigor y producción de grano. Los genotipos NCB 226, SER 16, SEN 56 y SEA 15 mostraron alta discriminación de 13C y mayor longitud de raíces entre 60 y 75 cm de profundidad en el suelo cuando crecieron en invernadero en condiciones de estrés por sequía (Figura 2); esta característica les permite extraer una mayor cantidad de agua desde los perfiles inferiores del suelo, y mantener altas tasas de transpiración e intercambio gaseoso para una mayor producción de grano. White et al. (1990) igualmente encontraron una relación positiva entre discriminación de 13C y la densidad de longitud de raíces. La hipótesis a partir de este resultado en frijol común sugiere que las plantas tolerantes a estrés hídrico tienen posibilidad para producir raíces más profundas, lo que les permite mayor acceso a agua, dando como resultado mayor conductancia estomática y mayor discriminación de 13C (Hall 2004; White et al., 1990; White 1993).
La línea SER 78 fue sobresaliente en producción de grano bajo estrés por sequía y presentó muy baja discriminación de 13C en grano, lo cual revela un uso eficiente del agua. Las líneas DOR 390, Tío Canela 75 y Carioca forman un grupo con uso eficiente de agua, menos discriminación de isótopo de carbono, pero tienen una baja producción de grano (Figura 3); en condiciones de riego esta característica no presentó correlación con la producción de grano (r = 0.04).
- En condiciones de estrés por sequía terminal en invernadero, algunas líneas de frijol común evaluadas presentaron características morfo-fisiológicas de raíces que se correlacionaron positivamente con producción de biomasa foliar, entre éstas: longitud total de raíz (cm/planta), profundización visual de raíz (cm) 35 días después de siembra, y longitud de raíz entre 40 y 60 y entre 60 y 75 cm en profundidad en el suelo.
- Los resultados permitieron caracterizar el grupo de 21 líneas de frijol evaluadas por sus expresiones fisiológicas e identificar características relacionadas con mayores rendimientos de granos. Dos líneas de grano negro y pequeño, SEN 56 y NCB 226, se caracterizaron por presentar buena profundización de raíces y buen vigor en campo, combinado con una mayor movilización de foto-asimilados para la formación de vaina y grano y por ende un mayor rendimiento en condiciones de sequía terminal. Las líneas SER 16 y SER 109, de grano rojo y pequeño, en condiciones de sequía terminal presentaron una buena profundización de raíces, lo que les permite tener alta tasa de transpiración y un buen vigor de planta, pero su producción de vainas y granos fue similar al promedio de las líneas evaluadas. La línea SER 78 presentó una buena eficiencia en el uso del agua, una característica importante en ambientes muy limitantes de este recurso. La línea SEA 15 sobresalió por la producción y profundización de raíces en condiciones de sequía, combinado con una mayor transpiración, lo cual la presenta como una opción para ser utilizada como planta padre para estas características.
- Las líneas testigo BAT 477 y Tío Canela 75 se caracterizaron por una buena producción y profundización de raíces bajo estrés por sequía terminal, combinado con una mayor eficiencia en el uso del agua (menor discriminación de isótopo de carbono), no obstante su producción de grano fue escasa, lo que evidencia que la selección sólo por vigor de raíces no es suficiente para lograr mayor adaptación a sequía en frijol común.
- Las evaluaciones en invernadero y campo demostraron la existencia de varias características fisiológicas relacionadas con tolerancia a sequía que son de utilidad en programas de mejoramiento genético de frijol; además, permitieron identificar líneas sobresalientes en estas características para ser usadas como parentales en los programas de cruzamiento, por tanto, es recomendable combinar diferentes características como movilización de fotoasimilados y vigor.
Los autores agradecen el apoyo recibido por fondos del Generation Challenge Programme y la Fundación Bill and Melinda Gates, en el proyecto Tropical Legumes I (TLI): Improving Tropical Legume Productivity for Marginal Environments in sub-Saharan Africa and South Asia. Igualmente agradecen a Miguel Grajales, Mariela Rivera y María del Pilar Hurtado por su ayuda profesional.
Beebe, S.; Rao, I. M.; Cajiao, C.; y Grajales, M. 2008. Selection for drought resistance in common bean also improves yield in phosphorus limited and favorable environments. Crop Sci. 48:582 - 592. [ Links ]
Beebe, S. E.; Rao, I. M.; Blair, M. W.; y Acosta- Gallegos, J. A. 2010. Phenotyping common beans for adaptation to drought. En: J. M. Ribaut y P. Monneveux (eds.). Drought phenotyping in crops: from theory to practice. Generation Challenge Program Special Issue on Phenotyping. p. 311 - 334. [ Links ]
Blum, A. 2002. Drought tolerance - is it a complex trait? En: Saxena, N.P. y J.C. O'Toole (eds.). Field screening for drought tolerance in crop plants with emphasis on rice. Proc. International Workshop on Field Screening for Drought Tolerance in Rice, 11-14 Dic. 2000. ICRISAT, Patancheru y Rockefeller Foundation, Nueva York. p. 17 - 22. [ Links ]
Bruce, W. B.; Edmeades, G. O.; y Barker, T. C. 2002. Molecular and physiological approaches to maize improvement for drought tolerance. J. Exp. Bot. 53:13 - 25. [ Links ]
Butare, L.; Rao, I. M.; Lepoivre, P.; Polania, J.; Cajiao, C.; Cuasquer, J. B., y Beebe, S. 2011. New sources of resistance in Phaseolus species to individual and combined aluminium toxicity and progressive soil drying stresses. Euphytica 181:385 - 404. [ Links ]
Condon, A. G.; Richards, R. A.; Rebetzke, G. J.; y Farquhar, G. D. 2004. Breeding for high wateruse efficiency. J. Exp. Bot. 55:2447 - 2460. [ Links ]
Farquhar, G. D.; Ehleringer, J. R.; y Hubick, K. T. 1989. Carbon isotope discrimination and photosynthesis, Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:503 - 37. [ Links ]
Hall, A. E. 2004. Comparative ecophysiology of cowpea, common bean and peanut. En: Physiology and biotechnology integration for plant breeding. Ed. Nguyen HT y Blum A. Marcel Dekker Inc. Nueva York. 271 - 325. [ Links ]
Ho, M. D.; Rosas, J. C.; Brown, K. M.; y Lynch, J. P. 2005. Root architecture tradeoffs for water and phosphorus acquisition. Funct. Plant Biol. 32:737 - 748. [ Links ]
Huang, B. y Gao, H. 2000. Root physiological characteristics associated with drought resistance in tall fescue cultivars. Crop Sci. 40:196 - 203. [ Links ]
Khan, H.; Link, W.; Hocking, T. J.; y Stoddard, F. L. 2007. Evaluation of physiological traits for improving drought tolerance in faba bean (Vicia faba L.). Plant Soil. 292:205 - 217. [ Links ]
Khan, H. R.; Paull, J. G.; Siddique, K. H.; Stoddard, F. L.. 2010. Faba bean breeding for drought-affected environments: a physiological and agronomic perspective. Field Crops Res. 115:279 - 286. [ Links ]
Morton, J. F. 2007. The impact of climate change on smallholder and subsistence agriculture. Proc. Nat. Acad. Sci. 104:19680 - 19685. [ Links ]
Polanía, J A.; Rao, I. M.; Beebe, S.; y García, R. 2009, Desarrollo y distribución de raíces bajo estrés por sequía en frijol común (Phaseolus vulgaris L.) en un sistema de tubos con suelo. Agron. Col. 27(1):25 - 32. [ Links ]
Rao, I. M. 2001. Role of physiology in improving crop adaptation to abiotic stresses in the tropics: The case of common bean and tropical forages. En: Pessarakli, M. (ed.). Handbook of Plant and Crop Physiology. Marcel Dekker, Inc., Nueva York. p. 583 - 613. [ Links ]
SAS Institute, 2008. Base SAS 9.2. Procedures Guide. Statistical procedures. SAS, Inst., Cary, NC, EE.UU. [ Links ]
Sorrells, M.E. 2007. Application of new knowledge, technologies, and strategies to wheat improvement. Euphytica 157:299 - 306. [ Links ]
Sponchiado, B.; White, J.; Castillo, J.; y Jones, P. 1989. Root growth of four common bean cultivars in relation to drought tolerance in environments with contrasting soil types. Exp. Agric. 25:249 - 257. [ Links ]
Thung, M. y Rao, I. M. 1999. Integrated management of abiotic stresses. En: Singh, S. (ed.). Common bean improvement in the twenty-first century. Kluwer Academic Publ., Dordrecht, Holanda. p. 331 - 370 [ Links ]
Turner, N. C. 1979. Drought resistance and adaptation to water deficits in crop plants. En: H. Mussell y R.C. Staples, (eds.). Stress physiology in crop plants. Wiley Interscience, Nueva York. p. 343 - 372. [ Links ]
Uriarte, C. A. 2003. Historia del clima de la tierra. 1a. Edición. Servicio Central de Publicaciones del Gobierno Vasco. ISBN: 84-457-2079-1. 306 p. [ Links ]
White, J. W. y Castillo, J. A. 1989. Relative effect of root and shoot genotypes and yield on common bean under drought stress. Crop Sci. 29:360 - 362. [ Links ]
White, J. W.; Castillo, J. A.; y Ehleringer, J. R. 1990. Associations between productivity, root growth and carbon isotope discrimination in Phaseolus vulgaris under water deficit. Aust. J. Plant Physiol. 17:189 - 198. [ Links ]
White, J. W. 1993. Implications of carbon isotope discrimination studies for breeding common bean under water deficits. En: Stable isotopes and plant carbon-water relations. En: Ehleringer J. R.; Hall, A. E.; y Farquhar, G. D. (eds.). Academic Press. San Diego. p. 387 - 398. [ Links ]
White, J. W.; Ochoa, M. R.; Ibarra, P. F.; y Sing, S. P. 1994. Inheritance of seed yield, maturity and seed weight of common bean (Phaseolus vulgaris) under semi-arid rainfed conditions. J. Agric. Sci. 122:265 - 273. [ Links ]
