










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.61 no.4 Palmira Oct./Dec. 2012
1Ingeniero Agrónomo. 2Química, M.Sc. 3Bióloga, Ph.D., Profesora Asociada, Dedicación Exclusiva. 4Facultad de Agronomía, Universidad Nacional de Colombia y Centro Colombiano de Genómica y Bioinformática en Ambientes Extremos–GEBIX, Bogotá, Colombia. *Autor para correspondencia: etorresr@unal.edu.co; †rgcaconc@unal.edu.co; ‡lmavellanedat@unal.edu.co
En la vereda El Bosque del Parque Nacional Natural de Los Nevados (PNNN), Colombia, se estimó la densidad poblacional de microorganismos asociados con el metabolismo de nitrógeno (N) en suelos bajo tres sistemas de uso: cultivo de papa, ganadería y zonas poco intervenidas (páramo). Indirectamente se cuantificaron microorganismos amonificantes (AMO), proteolíticos (PRO), oxidantes del amonio (BOA), oxidantes del nitrito (BON) y denitrificantes (DEN) en suelos de tres fincas ubicadas a diferente altitud y en épocas seca y lluviosa. Se realizó la prueba no-paramétrica de Kruskal-Wallis por pares, con un nivel de significancia P < 0.05, para detectar diferencias estadísticamente significativas entre las diferentes abundancias de grupos funcionales de microorganismos. Luego, se realizó análisis de componentes principales (ACP) para evaluar las relaciones entre las diferentes variables estudiadas. Los resultados mostraron diferencias para los grupos evaluados, siendo menor la densidad de microorganismos en suelos de páramo. Los PRO fueron más abundantes en todos los tipos de suelos, seguido de los DEN con mayor densidad en el ecosistema páramo. La densidad poblacional microbiana no fue afectada por la altitud, sin embargo se presentaron algunas diferencias que no mostraron un patrón único de comportamiento. Se observaron diferencias como resultado de la época climática, siendo mayor en la época seca para los grupos PRO, AMO, BOA y BON, y en época de lluvias para los DEN. Este estudio contribuye al conocimiento de las relaciones que existen entre el uso del suelo y la presencia de los microorganismos asociados con el ciclo de nitrógeno.
Palabras clave: Amonificantes, denitrificantes, oxidantes del amonio, oxidantes del nitrito, proteolíticos.
This study estimated the population density of microorganisms associated with the metabolism of nitrogen (N) in soils with three uses: potato crop, livestock and low intervention zones (high plains or páramo) in the El Bosque district of Los Nevados National Natural Park in Colombia. The following microorganisms were indirectly quantified: amonifiers (AMO), proteolytic (PRO), ammonium oxidizing (BOA), nitrite oxidizing (BON) and denitrifiers (DEN) in soils of three farms located at different altitudes and under two climatic seasons. The non-parametric Kruskal-Wallis test paired with a significance level of P < 0.05 was used to detect statistically significant differences among abundances of functional groups of microorganisms. Then a principal component analysis (PCA) was performed to evaluate the relationships among different variables. The results showed differences among all the groups, with páramo soils having the lowest density of microorganisms in the soil. The PRO microorganisms were the most abundant in all soils, followed by DEN, with a higher density in the páramo ecosystem. The microbial population density was not significantly affected by altitude but showed some differences that did not have a unique behavior pattern. Additionally, differences were seen in the seasons, being higher in the dry season for the PRO, AMO, BOA and BON groups; and in the rainy season for DEN. This study provided knowledge of the relationships between land use and the presence of microorganisms associated with the nitrogen cycle.
Key words: Ammonium oxidizing, amonifiers, denitrifiers, nitrite oxidizing, proteolytic.
La comunidad microbiana asociada con el ciclo del nitrógeno (N) representa una ventaja evolutiva al fijar N2 atmosférico y convertirlo en formas asimilables para otros organismos. El N está presente en varias formas, las cuales son transformadas a lo largo del ciclo por la acción de microorganismos amonificantes (AMO), proteolíticos (PRO), oxidantes de amonio (BOA), oxidantes de nitrito (BON) y denitrificantes (DEN), entre otros (Loomis y Connor, 2002).
El conocimiento sobre la diversidad y capacidad metabólica de los microorganismos del suelo asociados con el ciclo del N es escaso. No obstante, hay diferentes métodos para analizar las comunidades microbianas dependientes y no-dependientes de los cultivos (Nannipieri et al., 2003). Entre los primeros se encuentra la técnica del número más probable (NMP) que estima de manera indirecta la densidad de una población sin hacer conteo real de microorganismos o colonias. A pesar de que la información generada se restringe sólo a las comunidades que pueden ser cultivadas y es sensible a la densidad de inóculo (Kirk et al., 2004), es una técnica rápida y económica que genera información útil para estimar la abundancia de la comunidad microbiana en el suelo.
El tipo de uso del suelo se considera un factor que controla directa o indirectamente la estructura de comunidades de microorganismos en el suelo (Han et al., 2011). La biomasa microbiana es uno de los indicadores que mejor refleja las respuestas al cambio ambiental, incluso mejor que parámetros físicos y químicos (Kaschuk et al., 2011), y por tanto, es fundamental para la evaluación de la calidad del suelo (Acosta-Martínez et al., 2008).
El páramo se encuentra entre 3200 y 4500 m.s.n.m. y constituye un ecosistema frágil de gran importancia para el almacenamiento y la regulación hídrica (Hofstede et al., 2003). En Colombia este ecosistema abarca cerca de 30,000 km2, siendo el país con mayor área de páramos (Ramírez, 2011) y en el Parque Nacional Natural de los Nevados (PNNN) es el ecosistema más representativo con un área estimada de 38,600 ha (66.21%) (PNN de Colombia, 2011).
En los páramos colombianos se cultiva papa y se desarrollan actividades ganaderas, sin embargo, no se conoce el efecto de estas prácticas sobre las propiedades del suelo y las comunidades microbianas. Por tanto, el objetivo de este estudio fue estimar la densidad poblacional de microorganismos asociados con el metabolismo de N en suelos de la vereda El Bosque del PNNN, utilizados para cultivo de papa y ganadería, comparados con suelos de páramo con vegetación nativa. Se espera establecer posibles relaciones entre el uso del suelo y la presencia de microorganismos pertenecientes a los diferentes grupos funcionales asociados con el metabolismo de N.
Sitio y recolección de muestras
Las muestras de suelos rizosféricos para el estudio fueron tomadas en las fincas Buenos Aires (3769 m.s.n.m.), El Edén (3590 m.s.n.m.) y La Secreta (3432 m.s.n.m.) en la vereda El Bosque, municipio de Pereira, Risaralda. En cada sitio se evaluaron los usos del suelo: bosque de páramo, cultivo de papa (Solanum tuberosum) y ganadería en épocas seca y húmeda. La ganadería es doble propósito (producción de leche y carne), con énfasis en producción de leche dadas las condiciones de la zona. Los pastos que se siembran en la zona son Orchoro (Dactylis glomerata), Raygras (Lolium sp.) y Plegadera (Lachemilla sp.). El páramo presenta vegetación predominante de Cortaderia selloana, Pernettya prostrata, Buddleja sp., Lupinus albus, Dendropanax sp., Chusquea sp. En cada uno de los tipos de uso de suelo se realizaron tres muestreos, compuestos por 10 submuestras, para un total de tres usos del suelo x tres fincas x dos épocas x tres muestreos, para un total de 54 muestras. En cada muestra se determinó la abundancia de cinco grupos funcionales de microorganismos, asociados con el metabolismo de N: PRO, AMO, BOA, BON y DEN.
Abundancia de microorganismos y medios de cultivo
La abundancia de los microorganismos para los diferentes grupos funcionales se estimó mediante el método NMP (Cochran, 1950). Se utilizaron diversos medios de cultivo y fuentes de carbono y N según el grupo funcional considerado. Para PRO y BOA el medio de cultivo se preparó utilizando como base la solución salina de Winogradsky (Pochon, 1954) a razón de 50 ml/lt suplementado con una solución de oligoelementos (1 ml/lt) (Balch et al., 1977). Para los PRO se utilizó gelatina (30 g/lt) como única fuente de carbono y N, y un tiempo de incubación de 15 días. Para los BOA se utilizó carbonato de calcio (1 g/lt) y sulfato de amonio (0.5 g/lt), como fuentes de carbono y N, respectivamente, y un tiempo de incubación de 30 días (Flórez-Zapata y Uribe-Vélez, 2011). Para AMO se utilizó como medio de cultivo y única fuente de carbono y nitrógeno peptona bacteriológica (2 g/lt) y un tiempo de incubación de 24 h (IGAC, 2006). Para aislar los BON se empleó como medio de cultivo nitrito de potasio (0.006 g/lt), fosfato bipotásico (1 g/lt), sulfato de hierro heptahidratado (0.03 g/lt), cloruro de sodio (0.3 g/lt), sulfato de magnesio heptahidratado (0.1 g/lt), carbonato de calcio (1 g/lt) y cloruro de calcio (0.3 g/lt); el tiempo de incubación fue de 96 h (IGAC, 2006). Finalmente, para los DEN se utilizó como medio de cultivo una mezcla de dos soluciones, A y B, con la composición siguiente, solución A: nitrato de potasio (2 g/ lt), asparagina (2 g/lt), azul de bromotimol al 1% (10 g/lt); solución B: citrato de sodio (17 g/lt), fosfato mono potásico (2 g/lt), sulfato de magnesio (2 g/lt), cloruro de calcio hexahidratado (0.4 g/lt), cloruro férrico hexahidratado (0.1 g/lt). El tiempo de incubación fue de 72 h (IGAC, 2006) a 28°C.
Recuento de microorganismos
La presencia o ausencia de los organismos se determinó utilizando como características la licuefacción de la gelatina (PRO) y la producción de nitrito, medida con el reactivo de Griess Ilosvay (BOA) (Flórez-Zapata y Uribe- Vélez, 2011). El consumo de nitrito se determinó con el reactivo de Griess Ilosvay (BON), la producción de amonio por el reactivo de Nessler (AMO) y la producción de gas con la utilización de campanas de Durham y cambio de coloración azul (DEN) (IGAC, 2006). Los conteos fueron expresados en unidades formadoras de colonia/g de suelo seco (UFC/g).
Análisis estadístico
El análisis de datos sobre las diferentes abundancias de grupos funcionales de microorganismos se realizó mediante Anova, no obstante, debido a que los datos no soportaron el supuesto de normalidad, se realizó la prueba no-paramétrica de Kruskal-Wallis (KW) por pares con un nivel de significancia P < 0.05, para detectar diferencias estadísticamente significativas (Kruskal y Wallis, 1952). Lo anterior se realizó con el software estadístico SAS 9.0 (2009) y STATA 11.1 (2005). Luego, se hizo análisis de componentes principales (ACP) para evaluar las relaciones entre las diferentes variables estudiadas.
Se encontraron diferencias (P = 0.0017) en la abundancia de los diferentes grupos funcionales (Cuadro 1). En general, la abundancia microbial fue menor en suelo de páramo comparado con suelo en cultivo de papa y pastura, los cuales no presentaron diferencias entre sí (P = 0.289). De acuerdo con Roldán et al. (2009) esto posiblemente se debe a que en suelos de páramo las densidades para la mayoría de grupos funcionales relacionados con el ciclo del N son más bajas, ya que este uso posee una alta diversidad vegetal, ofreciendo sustratos que favorecen la riqueza de microorganismos, mas no la densidad, a diferencia de los usos con manejos intensivos. Sin embargo, la estrategia de conteos mediante el cultivo de microorganismos puede ser un factor limitante para la posible recuperación y conteo de aquellos que no son cultivables, los cuáles podrían estar cumpliendo una función determinada dentro del ciclo de N en el suelo, aunque no sean detectados.
Los suelos de páramo presentaron menor densidad de PRO comparados con los suelos usados para cultivo de papa y ganadería (Cuadro 1) debido, posiblemente, al aumento de nutrientes fácilmente disponibles en estos últimos a causa de la aplicación de fertilizantes de síntesis química (N, P, K), como también a las labores mecánicas que se realizan en estos suelos que aumentan los procesos de mineralización de los mismos y, por tanto, la disposición de nutrientes para el crecimiento de microorganismos. No obstante, los suelos en ganadería presentan menor densidad de microorganismos con respecto al cultivo de papa, ya que estos hacen parte de un sistema de rotación en el cual se cultiva papa por dos años y posteriormente son utilizados a ganadería (Ramírez, 2011).
La densidad de PRO no varió (P > 0.05) por efecto de la altitud, pero sí por efecto de la época (P = 0.0003), siendo mayor la abundancia en época seca (Cuadro 1). Lo anterior posiblemente se debe a que en la época de lluvias el agua afecta la temperatura del suelo, la humedad y el pH, factores que influyen en la actividad microbiana (Roldán et al., 2009).
La densidad de AMO varió por efecto del uso del suelo (P = 0.001) con una menor densidad en el ecosistema páramo (Cuadro 1). Calvo-Vélez et al. (2008) señalan que en suelos tropicales la tasa de amonificación es afectada por el pH del suelo; así, un pH entre 5.5 y 6.0 es ideal para una actividad máxima de los microorganismos, pero en condiciones de mayor acidez disminuye. En el presente estudio no se encontraron diferencias en pH debidas al uso de los suelos, por lo que en este factor no sería una condición limitante para la amonificación en el ecosistema páramo. Uno de los factores que puede condicionar la baja densidad de AMO en páramo es la menor cantidad de sustrato proveniente de PRO.
Las diferencias en las densidades de AMO en las altitudes del ensayo no fueron significativas (P > 0.05). Por otro lado, en la época seca se encontró una mayor densidad que en época de lluvias. Este comportamiento de los microorganismos AMO y PRO es concordante, ya que en suelos donde se presentaron menores PRO también hubo menores AMO, lo cual es coherente ya que los productos de los PRO son potenciales sustratos de los AMO. No obstante, estos últimos pueden ser tanto aerobios como anaerobios y en algunos casos son capaces de formar estructuras reproductivas de resistencia viables para periodos adversos, ya sea por falta de humedad, carencia de nutrientes o anegamiento (Ponzuelo, 1991).
Las concentraciones de BOA fueron diferentes (P < 0.0001) con respecto al uso del suelo, siendo los suelos en ganadería los que presentaron las mayores densidades (Cuadro 1), pero no ocurrió lo mismo con el cultivo de papa (P = 0.526); al igual que con los grupos anteriores, la densidad de BOA fue menor en el ecosistema páramo. Flórez-Zapata (2010) encontraron 1.5x102 propágulos/g de BOA en un suelo cultivado con papa, y Philippot y Germon (2005) consideran que densidades entre 105 y 107 UFC bacteriales/g son comunes en suelos de uso agrícola.
Las densidades de BOA en suelos con ganadería pueden estar relacionadas con el aumento de residuos orgánicos por estiércol de ganado y orina de los animales, los cuales son un sustrato ureico promotor de crecimiento de microorganismos (Orozco-Patiño, 1999). El aumento de microorganismos en suelos con cultivo con respecto al páramo puede estar relacionado con el uso de fertilizantes, lo cual aumenta la disponibilidad de N, que puede ser empleado como fuente de energía por microorganismos quimiolitotrofos (BOA y BON) (Roldán et al., 2009). Además, la biomasa total de microorganismos involucrados en el ciclo de N incrementa con la intensidad del uso del suelo en pasturas (Berner et al., 2011). No obstante, una cantidad de NH4+ mayor que 800 mg/kg inhibe el desarrollo de la microflora nitrificante autótrofa (Ponzuelo, 1991), lo que hace posible utilizar el grupo funcional (BOA) como indicador de los cambios por uso del suelo (Roldán et al., 2009).
Los BOA presentaron un aumento en la densidad poblacional a medida que disminuyó la altitud, esto puede estar relacionado con el aumento de temperatura que se presenta a medida que la altura disminuye, aproximadamente 0.57 °C por cada 100 m (Guevara- Díaz, 2003).
Los BON presentaron una baja densidad poblacional en el ecosistema páramo (Cuadro 1), lo cual puede estar relacionado con la baja cantidad de sustrato proveniente de la oxidación de NH4+ en este ecosistema llevada por BOA.
Ponzuelo (1991) considera que el NO2- es una forma fácilmente inmovilizable debido a su capacidad de reaccionar con la materia orgánica del suelo (MOS), en especial con compuestos fenólicos. Estas reacciones ocurren en todo tipo de suelos y son favorecidas por la acidez y los altos contenidos de MOS, como los encontrados en suelos de páramo cuyo rango varió entre 9.05 y 18.81% (datos no presentados).
Los BON no mostraron diferencias entre las altitudes de las fincas en el estudio ni por efecto de la época, aunque se observó mayor densidad en la época seca (Cuadro 1). Los DEN revelaron menor densidad en páramo y mayor en ganadería (P = 0.006).
La denitrificación es un proceso heterotrófico que ocurre en condiciones de anoxia o hipoxia (Francis et al., 2007) y está relacionado con ambientes de humedad y densidades aparentes altas, como es el caso del uso de suelo para ganadería, generado por procesos de compactación en los primeros 20 cm, que puede favorecer altas densidades de DEN (Ramírez, 2011). Esto también conlleva una disminución de la biomasa microbiana del suelo (Li et al., 2004) y un aumento de la actividad de microorganismos DEN, que representa cerca de 5% de la biomasa microbiana del suelo (Hai, 2009), es decir, que dichos microorganismos se encuentran en altas densidades en los suelos comparado con AMO, BOA y BON. Flórez-Zapata (2010) considera que los DEN responden a las aplicaciones de fertilizantes minerales, debido a que obtienen energía a partir de estos compuestos. En el presente estudio los DEN fueron afectados por la época del año (P = 0.001), siendo más alta su concentración en época de lluvias (Cuadro 1). No obstante, en páramo y ganadería de la finca El Edén se observó mayor actividad en la época seca, lo que fue debido, posiblemente, a la escasa variación de la humedad en el suelo entre épocas (datos no publicados).
En el análisis multivariado se observaron diferencias en concentración de microorganismos debidas al uso del suelo (Figura 1), en donde el suelo de páramo presenta un comportamiento diferente a los suelos en cultivo de papa y ganadería; estos últimos se correlacionan (0.8 y 0.73, respectivamente) al eje 1, mientras que en el páramo la correlación al eje 1 fue negativa (-0.48). La tendencia similar entre los valores de sistema ganadería/cultivo de papa puede ser debida a la rotación entre ellos lo cual puede ocasionar fragmentación de complejos humus-alófanas que favorecen la descomposición de la MOS y el incremento de la densidad de microorganismos asociados a estos procesos (Orozco- Patiño, 1999).
Los AMO presentaron la menor densidad en suelos con uso de ganadería, esto puede deberse a que el estiércol de ganado contiene compuestos como metilamina que inhibe el funcionamiento de los AMO (Orozco-Patiño, 1999). Así mismo ocurre en cultivo debido a las aplicaciones de urea durante la fertilización, ya que como es de esperar, la aplicación de fertilizantes químicos incrementa la disponibilidad de nutrientes (Roldán et al., 2009). Por otra parte, se observó una menor densidad de microorganismos en el ecosistema páramo (Cuadro 2 y Figura 1) lo cual puede estar relacionado con la estabilidad de la MOS, por el predominio de sustancias húmicas y presencia de alófana que no permiten la descomposición de esta materia debido a su estructura estable.
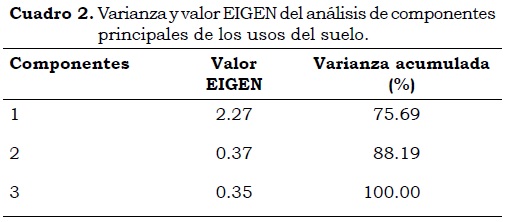
Se observó que los PRO fueron más abundantes en el páramo, lo que es importante debido a que las proteínas, la principal fuente de nitrógeno en los ecosistemas naturales, son necesarias para otros procesos en los que están involucrados los demás grupos funcionales (Li et al., 2004).
En el ACP realizado con las variables: PRO, AMO, BON, BOA y DEN (Cuadro 3; Figura 2 y Figura 3) se tomaron los tres primeros componentes, debido a que acumulan y explican el 83.02% de la varianza de los datos. En la Figura 2 se observa que las variables más correlacionadas en el eje 1 fueron PRO, AMO y BON, con valores de 0.51; 0.74 y 0.76, respectivamente, mientras que en el eje 2 están relacionados DEN y BOA con 0.86 y 0.37 respectivamente. El CP1 está relacionado con procesos de mineralización, donde los PRO fragmentan la materia orgánica en aminoácidos libres, los cuales posteriormente por la actividad de los AMO son transformados a amonio que sirve como sustrato para la nitrificación (Ponzuelo, 1991), proceso que ocurre por la intervención de grupos distintos de microorganismos, bacterias oxidantes de amonio (BOA) y bacterias oxidantes de nitrito (BON) (Francis et al., 2007).
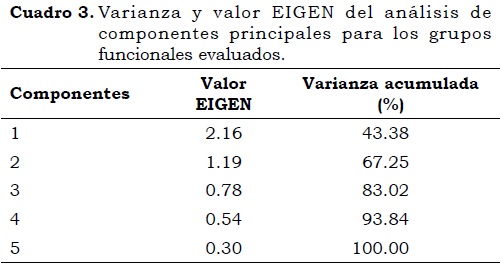
Los procesos de denitrificación, donde participan los DEN reduciendo el NO3- a NO2-, N2O y NO (CP2) se encuentran relacionados con los BOA (Figura 2), esto puede deberse a que en condiciones de intensificación agrícola, incrementos en el pH, temperatura y porosidad del suelo facilitan la conversión de amonio a amoniaco (NH3+) y éste se volatilice en forma de nitrógeno lo que limita la fuente de sustrato para la oxidación de amonio (Flórez-Zapata, 2010). Es posible que exista relación entre BOA y DEN, como lo proponen Francis et al. (2007) y Le Roux et al. (2008) quienes consideran que el proceso de oxidación de amonio puede ocurrir en condiciones anaeróbicas (Anammox), sin embargo, aún no es claro el papel de este tipo de bacterias en el ciclo edáfico del nitrógeno.
En la Figura 3 se observa que los PRO están más correlacionados con el eje 3, comparado con los DEN, por lo que aparentemente presentan un comportamiento independiente. Con lo anterior es posible relacionar cada uno de los componentes con un proceso en el ciclo del N, donde el eje 1 (CP1) representa procesos de amonificación y oxidación de N, mientras que el CP2 representa procesos de pérdida de N de los ecosistemas por denitrificación o Anammox, y el CP3 indica procesos relacionados con la fragmentación de proteínas. En general, las poblaciones de microorganismos PRO muestran abundancias superiores respecto a los microorganismos nitrificantes y denitrificantes, y esto se debe a que en los suelos hay una alta disponibilidad de materia orgánica que favorece el desarrollo de estos microorganismos (Flórez-Zapata, 2010).
- Se encontraron diferencias entre las densidades de poblaciones de las comunidades microbianas en relación con el uso del suelo y las prácticas agrícolas. La menor densidad de grupos funcionales se observó en ecosistema de páramo y la mayor en sistemas con cultivo de papa y ganadería, lo cual puede ser debido al efecto de fertilizaciones y al manejo mecánico del suelo.
- Los PRO fueron el grupo funcional de mayor abundancia en los ecosistemas evaluados, lo que indica que en ecosistemas poco intervenidos dichos microorganismos median el proceso que proporciona energía para los demás microorganismos en el ciclo del N.
- En general, la densidad poblacional microbiana no se afectó significativamente por la altitud, no obstante se presentaron pequeñas diferencias sin una tendencia definida.
- Los grupos funcionales PRO, AMO, BOA, BON presentaron mayor densidad en la época seca, lo que se relaciona con la humedad en el suelo, ya que estos microorganismos son aerobios y su abundancia es afectada en condiciones de hipoxia en el suelo. Lo contrario ocurrió con los DEN, que son microorganismos anaerobios.
- Los resultados mostraron mayor densidad de DEN en ecosistema de páramo, comparado con los demás grupos funcionales (a excepción de PRO), lo que sugiere que en este ecosistema existe un balance negativo en el ciclo del N, donde las pérdidas por denitrificación pueden ser mayores que las ganancias por mineralización.
Al Centro Colombiano de Genómica y Bioinformática de Ambientes Extremos – GeBiX. A Colciencias y a la Universidad Nacional de Colombia, quienes financiaron y suministraron los recursos necesarios para la realización de esta investigación. A Luis Miguel álvarez y a la comunidad de la vereda El Bosque, por su valiosa atención y colaboración para la realización de esta investigación.
Acosta-Martínez, V.; Acosta-Mercado, D.; Sotomayor -Ramírez, D.; y Cruz-Rodríguez, L. 2008. Microbial communities and enzymatic activities under different management in semiarid soils. Appl. Soil Ecol. 38:249 - 260. [ Links ]
Balch, W. E.; Schoberth, S.; Tanner, R. S.; y Wolfe, R. S. 1977. Acetobacterium, a new genus of hydrogen- oxidizing, carbon dioxide-reducing, anaerobic bacteria. Int. J. Syst. Bacteriol. 27(4):355 - 361. [ Links ]
Berner, D.; Marhan, S.; Keil, D.; Poll, C.; Schützenmeister, A.; Piepho, H.-P.; y Kandeler, E. 2011. Land-use intensity modifies spatial distribution and function of soil microorganisms in grasslands. Pedobiologia 54(5-6):341 - 351. [ Links ]
Calvo-Vélez, P.; Meneses, L. R.; y Zúñiga-Dávila, D. 2008. Estudio de las poblaciones microbianas de la rizósfera del cultivo de papa (Solanum tubersosum) en zonas altoandinas. Ecol. Apl. 7(1 - 2):141 - 148. [ Links ]
Cochran, W. G. 1950. Estimation of bacterial densities by means of the Most Probable Number. Biometrics 6(2):105 - 116. [ Links ]
Flórez-Zapata, N. 2010. Determinación de parámetros biológicos y físico-químicos asociados al metabolismo edáfico del nitrógeno en cultivos de Solanum phureja en el departamento de Cundinamarca. Tesis de Maestría. Facultad de Ciencias. Universidad Nacional de Colombia. Bogotá. 105 p. [ Links ]
Flórez-Zapata, N.; y Uribe-Vélez, D. 2011. Biological and physicochemical parameters related to the nitrogen cycle in the rhizospheric soil of native potato (Solanum phureja) crops of Colombia. Appl. Environ. Soil Sci. 2011. Article ID 847940, 10 p. [ Links ]
Francis, C. A.; Beman, J. M.; y Kuypers, M. M. 2007. New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation. Intern. Soc. Micr. Ecol. 1:19 - 27. [ Links ]
Guevara-Díaz, J. M. 2003. Métodos de estimación y ajuste de datos climáticos. Consejo de Desarrollo Científico y Humanístico. Universidad Central de Venezuela. Caracas. 133 p. [ Links ]
Hai, B. 2009. Impact of environmental factors on key functional groups involved in nitrogen cycling in agricultural ecosystems. Doktorgrades der Naturwissenschaften. Fakultät für Biologie. Ludwig- Maximilians-Universität München. 114 p. [ Links ]
Han, X.; Wang, R.; Guo, W.; Pang, X.; Zhou, J.; Wang, Q.; Zhan, J.; y Dai, J. 2011. Soil microbial community response to land use and various soil elements in a city landscape of north China. Afr. J. Biotechn. 10(73):16554 - 16565. [ Links ]
Hofstede, R.; Segarra, P.; y Mena-Vásconez, P. 2003. Los páramos del mundo: Proyecto Atlas Mundial de los Páramos. Global Peatland Initiative/NCIUCN/ EcoCiencia. Quito. 299 p. [ Links ]
IGAC (Instituto Geográfico Agustín Codazzi). 2006. Métodos analíticos de laboratorio de suelos. IGAC. Sexta ed. 648 p. [ Links ]
Kaschuk, G.; Alberton, O.; y Hungria, M. 2011. Quantifying effects of different agricultural land uses on soil microbial biomass and activity in Brazilian biomes: inferences to improve soil quality. Plant Soil 338:467 - 481. [ Links ]
Kirk, J. L.; Beaudette, L. A.; Hart, M.; Moutoglis, P.; Klironomos, J. N.; Lee, H.; y Trevors, J. T. 2004. Methods of studying soil microbial diversity. J. Microbiol. Methods 58:169 - 188. [ Links ]
Kruskal, W. H. y Wallis, W. A. 1952. Use of ranks in one criterion variance analysis. J. Amer. Stat. Association 47(260):583 - 621. [ Links ]
Le Roux, X.; Poly, F.; Currey, P.; Commeaux, C.; Hai, B.; Nicol, G. W.; Prosser, J. I.; Schloter, M.; Attard, E.; y Klumpp, K. 2008. Effects of aboveground grazing on coupling among nitrifier activity, abundance and community structure. Intern. Soc. Micr. Ecol. 2(2):221 - 232. [ Links ]
Li, Q.; Allen, H. L.; y Wollum II, A. G. 2004. Microbial biomass and bacterial functional diversity in forest soils: effects of organic matter removal, compaction, and vegetation control. Soil Biol. Biochem. 36(4):571 - 579. [ Links ]
Loomis, R. S. y Connor, D. J. 2002. Ecología de Cultivos. Mundi-Prensa Libros. Madrid. 591 p. [ Links ]
Nannipieri, P.; Ascher, J.; Ceccherini, M. T.; Landi, L.; Pietramellara, G.; y Renella, G. 2003. Microbial diversity and soil functions. Eur. J. Soil Sci. 54:655 - 670. [ Links ]
Orozco-Patiño, F. H. 1999. La biología del nitrógeno: Conceptos básicos sobre sus transformaciones biológicas. Universidad Nacional de Colombia. Sede Medellín. Facultad de Ciencias. Editorial Gráficas Montoya, Medellín. 231 p. [ Links ]
Philippot, L. y Germon, J. C. 2005. Contribution of bacteria to initial input and cycling of nitrogen in soils. En: Buscot, F. y Varma, A. (eds.). Soil Biology, Volume 3, Microorganisms in Soils: Roles in Genesis and Functions. © Springer-Verlag Berlin Heidelberg. p. 159 - 176. [ Links ]
PNN (Parques Nacionales Naturales de Colombia). 2011. Disponible en: http://www.parquesnacionales.gov.co [Fecha revisión: Julio de 2011] [ Links ]
Pochon, J. 1954. Manuel technique d'analyse microbiologique du sol. Masson et Cie Editeurs. Paris. 124 p. [ Links ]
Ponzuelo, J. 1991. Estudio de grupos funcionales de microorganismos edáficos en la rizosfera de Alnus glutinosa (L.) Gaetern. Tesis Doctorado en Ciencias Biológicas. Universidad Complutense. Madrid. 270 p. [ Links ]
Ramírez, M. 2011. Los páramos en la historia. Rev. Páramos 1:42 - 57. [ Links ]
Roldán, F.; Varela, A.; García, E.; Sierra, L.; Vallejo, V.; Berdugo, M.; Aguilera, M.; Santos, D.; Cubillos, A.; Gómez, M.; Gómez-Sarmiento, M.; Latorre, N.; y Vela, A. 2009. Evaluación del efecto de diferentes usos del suelo sobre grupos funcionales microbianos edáficos en la ecorregión cafetera colombiana. En: Rodríguez, J. M.; Camargo, J. C.; Niño, J.; Pineda, A. M.; Arias, L. M.; Echeverry, M. A.; y Miranda, C. L. (eds.). Valoración de la biodiversidad en la ecorregión del Eje Cafetero. Centro de Investigaciones y Estudios en Biodiversidad y Recursos Genéticos. Ciebreg. Pereira, Colombia. p. 209-221. [ Links ]
SAS Institute Inc. 2009. SAS/STAT® 9.2 User's Guide: Mixed Modeling (Book Excerpt). Cary, NC (USA). [ Links ]
Stata Corp, Stata Statistical Software. 2005. Release 9, StataCorp LP, College Station, Tex, USA. [ Links ]
