










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Agronómica
versão impressa ISSN 0120-2812
Acta Agron. vol.62 no.1 Palmira jan./mar. 2013
Fitomejoramiento
1Ingeniero Agrónomo, estudiante de Maestría en Ciencias Agrarias con énfasis en Fitomejoramiento, Universidad Nacional de Colombia sede Palmira. 2Ingeniero Agrónomo, Msc Bioecnología, Docente Universidad de la Salle. 3Ingeniero Agrónomo, Ph.D. Profesor Asociado, Universidad Nacional de Colombia, sede Palmira.
*Autor para correspondencia: fesilvaag@gmail.com, fsilvaag@unal.edu.co; bersilva@unisalle.edu.co; ‡magarciad@unal.edu.co
El peso de frutos por planta es una característica compleja y controlada por múltiples genes donde el ambiente interfiere en la manifestación fenotípica. La estimación de componentes génicos como los de interacción epistática es de gran importancia en el proceso de mejoramiento genético de una especie. El objetivo de este estudio fue la estimación de parámetros genéticos para el peso del fruto por planta, usando un análisis de medias generacionales (P1, P2, F1, F2, RC1 y RC2), derivados del cruzamiento entre las líneas endocriadas de pimentón serrano y aji cayenne cultivar 958, Capsicum annuum L. El análisis reveló que los componentes genéticos de aditividad, dominancia y las interacciones epistáticas digénicas fueron significantes en la manifestación fenotípica de las variables analizadas, mostrando la necesidad de utilizar un modelo con cuatro parámetros genéticos, donde los efectos aditividad x aditividad [i], revelaron ser significantes para la variable objeto de análisis. Se estimó la presencia de epistasis recesiva o doble recesiva. Finalmente, para aumentar el rendimiento promedio de una población de ají cayene y pimentón, Capsicum annuum L., entendido como el peso de frutos por planta, se puede implementar un método de mejoramiento de selección recurrente, ya que las interacciones epistáticas pueden ser efectivamente explotadas a través de la hibridación entre líneas promisorias.
Palabra clave: Acción génica, aditividad, dominancia, epistasis, heredabilidad.
Yield is a complex trait and is under the control of multiple genes in which the environment interfere on the phenotype. The estimation of genetic components, including non-allelic interactions acquire a great importance on the breeding of a crop. The objective of this study was to estimate the genetic parameters for yield, using generation means analysis (P1, P2, F1, F2, BC1 and BC2) derived from the cross of Pimenton Serrano and Aji Cayenne 958 (inbreed lines of Capsicum annuum). The analysis revealed that additive, dominance and the non-allelic interactions components were significantly for the phenotypic expression, indicating the need for a four-parameters model.Additive by additive [i] effects revealed significance for fruit weight. Recessive or double recessive epistasis, was found. Finally, a breeding program based on recurrent selection to increase the average yield in a pepper (Capsicum annuum) population can be used, due to the epistatic interaction can be effectively exploited through the hybridization among promissory lines.
Key words: Additive, dominance, epistasis, gene action, heritability.
El género Capsicum comprende los ajíes y pimentones, que corresponden a especies picantes y no picantes, respectivamente. Los frutos del ají (Capsicum spp.) están entre los vegetales de mayor consumo en el mundo debido a su pungencia. Esta característica, única del género, es debida a la presencia de compuestos alcaloides del grupo capsaicinoides, presentes en la placenta de los frutos maduros. Varios de los estudios básicos relacionados con el mejoramiento genético del género giran en torno al tipo y magnitud de la variabilidad genética. En algunos casos no se logran modelos válidos para explicar lo observado en las mediciones experimentales, lo que implica la necesidad de ampliar el modelo a uno más complejo que incluya términos de interacción alélica o epistasis (Dhall y Hundal, 2006).
Según Pech et al. (2010) en estudios previos se ha observado la manifestación de vigor híbrido en características de rendimiento. Las cruzas dialélicas son la metodología más utilizada para estimar la herencia de características de rendimiento e identificar los progenitores superiores utilizados en el desarrollo de híbridos y cultivares (Yan y Hunt, 2002, citados por Pech et al, 2010). Según Dhall y Hundal (2006) el rendimiento de Capsicum annuum L. es un carácter complejo y poligénico, en el cual una selección de genotipos superiores basado en componentes de rendimiento, tales como número y peso de frutos por planta, puede ser la más eficiente. Se ha estimado que los efectos aditivos y dominantes en las características del rendimiento son influyentes en la expresión fenotípica; sin embargo si se encuentra que los efectos dominantes son mayores que los aditivos, significa que la dominancia contribuye más en la herencia del número y peso de frutos (Dhall y Hundal, 2006).
En las pruebas realizadas por el Programa de Mejoramiento Genético de Hortalizas de la Universidad Nacional de Colombia sede Palmira se han identificado nichos específicos de consumo de materiales con pungencia, en especial del género Capsicum, entre ellos materiales con características morfológicas de Capsicum annuum L. dulce, pero con contenidos medios y altos de capsaicina, que son muy bien aceptados y despiertan expectativa de nuevos mercados que pueden ser explorados mediante el desarrollo de líneas específicas (García, 2006; Pardey et al., 2006, 2009, 2010).
El conocimiento de la estructura genética y de los mecanismos de herencia de los componentes de rendimiento es esencial para la implementación de un programa de mejoramiento genético encaminado a incrementar los rendimientos por hectárea. Debido a esto, la presente investigación se orientó al análisis de la naturaleza genética y a estimar el tipo de acción génica del peso de frutos por planta en un cruzamiento intra-especifico de la especie Capsicum annuum L., con el fin de desarrollar programas de mejoramiento del rendimiento promedio de la especie.
En el análisis se utilizaron los parentales Pimentón cultivar Serrano y Ají Cayene cultivar 958, con un grado de homocigosis predicho de 96.875% por ser líneas F6 provenientes de procesos de autofecundación consecutivos y con resistencia al virus PepDMV. Ambos parentales presentan características contrastantes en cuanto a peso de frutos por planta. A partir de estas dos líneas se generaron las poblaciones F1, F2 y las generaciones de retrocruza. Las seis generaciones (P1, P2, F1, F2, RC1 y RC2) fueron cultivadas en el segundo semestre de 2010 en las instalaciones del centro experimental de la Universidad Nacional de Colombia sede Palmira, CEUNP, a 3° 24' N y 76° 26' O. Los materiales fueron sembrados bajo un diseño de bloques completos al azar con tres bloques y 18 plantas por bloque de las generaciones homogéneas P1, P2 y F1, 63 plantas por bloque de cada una de las generaciones de retrocruza y 180 plantas por bloque de la generación F2. De cada planta individual se midieron los pesos de fruto (peso de frutos) durante siete pases de cosecha, los cuales fueron totalizados para generar un promedio ponderado.
Para estimar parámetros genéticos en las seis generaciones se hizo un análisis de medias generacionales en la variable medida. Se utilizó el modelo completo de Mather y Jinks (1971) para una media observada en la k-ésima generación, incluyendo todos los efectos génicos. Como los medias de cada generación pueden variar con respecto a la precisión y a los diferentes tamaños de muestra de cada progenie, se realizó una prueba de homogeneidad de varianzas por medio del test de Bartlett al 5% (Ceballos, 1996). Los resultados del test de Bartlett (datos no mostrados) revelaron heterogeneidad en las varianzas de las generaciones no segregantes, razón por la cual se hizo una ponderación del modelo por los recíprocos de los errores estándar al cuadrado o el recíproco de la varianza de cada media (Mather y Jinks, 1971; Kearsey y Pooni, 1998) en una matriz de covarianza donde los elementos de ponderación quedan en la diagonal (Mather y Jinks,1971).
Se evaluó la presencia de las interacciones no alélicas usando la prueba de escalas agrupadas o Joint Scaling Test' (Mather y Jinks, 1971; Ceballos, 1996). Los valores observados de las seis generaciones se utilizaron para estimar los valores de la media parental, m, aditividad, d, dominancia, h, y las interacciones epistáticas digénicas aditivas por aditivas, i, y aditivas por dominantes, j, y/o dominantes por dominantes, l. Estos parámetros genéticos se añadieron uno a uno al modelo, con el fin de encontrar un modelo de cinco, cuatro o tres parámetros con el mejor ajuste. El ajuste del modelo se evaluó mediante el Joint Scaling Test y chi-cuadrado. Adicionalmente, se estimaron los parámetros de varianza ambiental, E, dominante, H, aditiva, D y número de genes, n, a través del método de Medias/Varianzas Generacionales propuesto por Mather y Jinks (1971), de la forma siguiente:
E = ¼ (s2P1 + s2P2 +2 s2F1)
D = 4s2F2 -2(s2RC1 + s2RC2)
H = 4(s2RC1 + s2RC2 - s2F2 – E)
n = ((P1 - P2)2(1 + 0.5k2))/8 s2g
De acuerdo con los datos que se observan en el Cuadro 1, el parental 958, P1, fue el de menor promedio (33.7 g por fruto/planta), siendo el de menor expresión fenotípica. Los valores medios de la generación F1 no revelan efectos de sobredominancia. Los valores medios de la generación de retrocruza RC1, fueron similares al parental inferior (Cayene cultivar 958), mientras que la RC2 fue similar al parental superior.
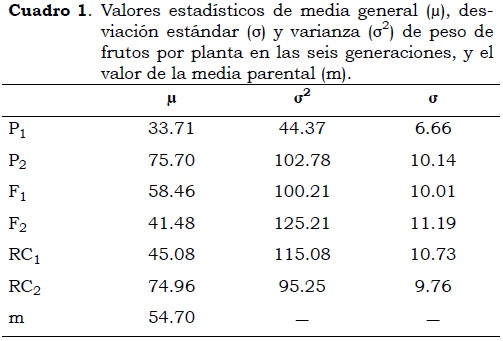
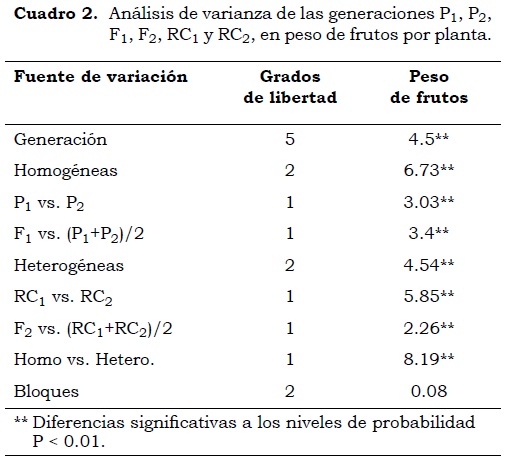
El análisis de varianza mostró diferencias altamente significativas (P < 0.01) en las generaciones (Cuadro 2). Esto indica que se acepta la hipótesis alterna, donde existe variabilidad genética en al menos una de las seis generaciones, creando un efecto diferente de las demás sobre la variable dependiente. De acuerdo con los contrastes ortogonales, se encontraron diferencias altamente significativas (P < 0.01) entre los parentales (P1 vs. P2), ello revela que estos fueron contrastantes para la variable de peso de frutos por planta. La comparación entre generaciones homogéneas y heterogéneas da un estimativo sobre el efecto genético en los materiales a usar y permite confirmar la segregación que existe en las generaciones F2, RC1 y RC2. El hallazgo de diferencias altamente significativas (P < 0.01) indica que las generaciones heterogéneas (F2, RC1 y RC2) presentan individuos genéticamente similares a los parentales y a la generación F1.
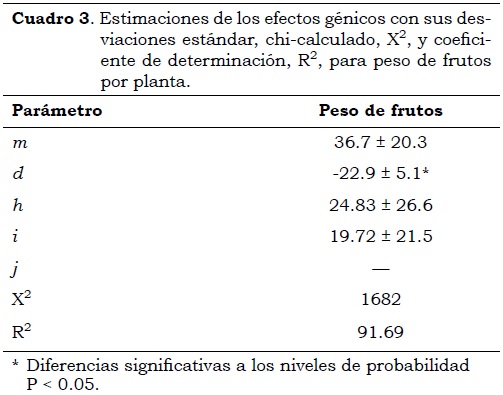
Cuando se estimaron los efectos génicos y el modelo fue desarrollado agregando secuencialmente parámetros, el mejor ajuste de los datos se obtuvo con un análisis de medias generacionales considerando un modelo con cuatro parámetros genéticos, siendo, por consiguiente, suficiente el uso de las seis generaciones para la estimación de los parámetros genéticos en peso de frutos por planta. El modelo de aditividad-dominancia no fue suficiente para explicar la variabilidad de los datos, donde los efectos de interacción epistáticos de aditividad por aditividad [i] mostraron tener influencia en la expresión fenotípica (Cuadro 3). Se debe anotar que los efectos aditivos mostraron diferencias significativas negativas (P < 0.05). El signo negativo en el efecto de aditividad no reviste importancia, ya que ello se debe a que el parental Cayene cultivar 958 elegido como el P1, es el que tiene menor peso de frutos promedio por planta, por lo cual puede ser identificado para esta variable como el parental que muestra la menor expresión fenotípica (Cukadar-Olmedo y Miller, 1997).
Los estimativos de los componentes de varianza aditivos, dominantes y ambientales, heredabilidades en sentido estricto y amplio y heterosis para las variables aparecen en el Cuadro 4. Se observa que la varianza dominante fue nula, causando un efecto en el valor de las heredabilidades. Este valor nulo en la varianza es debido posiblemente a que los valores de todos y cada uno de los efectos dominantes individuales fueron igualmente nulos, lo cual indicaría que para cada alelo hay sólo acción génica aditiva y ausencia de dominancia (Ceballos, 1996). Otra explicación es que los valores de todos los efectos dominantes individuales son distintos a cero pero, al tener signos opuestos, el resultado final es el de una sumatoria igual a cero (Ceballos, 1996). La heredabilidad en sentido estricto menor que la heredabilidad en sentido amplio, indica la presencia de efectos de dominancia que influyen en la manifestación de la característica. El estimativo del grado medio de dominancia (GMD) encontrado en este experimento no es evidencia fehaciente de ausencia de dominancia, ya que es posible obtener este resultado cuando los efectos individuales de dominancia se cancelan entre sí (Ceballos, 1996). La generación F1 se encuentra sólo 7% por encima del promedio parental, lo que muestra la gran importancia de los efectos aditivos en la expresión fenotípica.

En el peso de frutos por planta los efectos aditivos revelaron mayor significancia en la herencia de la característica, permitiendo emplear un proceso de mejoramiento genético en el cual se dé prioridad a la selección de plantas con mayores pesos de fruto. El signo en los efectos dominantes está en función del valor medio de la generación F1, en relación con la media parental, e indica cuál de los parentales está contribuyendo a los efectos dominantes. (Cukadar-Olmedo y Miller, 1997). El valor positivo de los efectos dominantes, los cuales muestran el comportamiento de los genes en combinaciones heterocigóticas, indica que el parental 958 tipo Cayene es el que más contribuye en la expresión.
Los efectos aditivos han sido los más representativos con efectos epistáticos bajos y no significativos en el desarrollo final de las diferentes generaciones. Sin embargo, el hecho de que los efectos epistáticos contribuyen a la expresión del peso de los frutos, sugiere que la herencia de esta característica es compleja y puede deberse a caracteres poligénicos. La presencia de efectos epistáticos de aditividad-aditividad [i] en el peso de frutos por planta, indica que esta característica puede ser fijada en una población a través de selección y/o producción de líneas endocriadas (Azizi et al., 2006). En esta variable se presenta un tipo de interacción de epistasis recesiva o doble recesiva, ya que según Mather y Jinks (1971) cuando los efectos [h], [i] y [l] tienen los mismos signos, existe este tipo de interacción epistatica. Sin embargo, no es posible distinguir entre estas alternativas ya que se requiere del conocimiento de las magnitudes relativas y los signos individuales de los efectos [d] de los genes que interactúan. Los signos asociados con los efectos epistáticos aditividad-aditividad [i] y aditividad-dominancia [j] indican la dirección en la cual los efectos génicos influencian la media poblacional (Azizi et al., 2006), además de mostrar un indicativo sobre la dispersión o asociación de genes en los parentales(Mather y Jinks, 1971). En la variable peso de frutos por planta, el signo positivo de los efectos digénicos aditivos-aditivos, indican que no existen niveles de dispersión en los genes de los parentales (Mather y Jinks, 1971).
Para aumentar el peso promedio del fruto por planta en una población de ají Cayene y Pimentón, Capsicum annuum L., se puede proceder con la generación de líneas endocriadas a partir de un procedimiento como el de descendencia de semilla única, ya que las otras interacciones epistáticas digénicas pueden ser efectivamente explotadas a través de la hibridación entre líneas promisorias tratando de acumular genes favorables. Los bajos valores en la heredabilidad en sentido estricto para la característica, sugiere que la herencia es compleja y no tiene relación con el nivel de significancia de los efectos aditivos, además de mostrar la presencia de genes con acciones no aditivas. Aunque los estimativos de la varianza dominante sean nulos, se ve claramente el efecto de dominancia influenciando la característica, ya que los valores de heredabilidad en sentido estricto fueron inferiores a los de la heredabilidad en sentido amplio. El grado de dominancia que una línea ejerce sobre otra en la expresión del carácter en estudio indica que existe una ausencia de dominancia, donde ambos alelos contribuyen en la expresión del carácter. No obstante, este estimativo no es lo suficientemente cierto, ya que este grado de dominancia depende de los efectos individuales de aditividad y dominancia y por tanto, en casos donde este estimativo sea igual a cero (0) no es evidencia fehaciente de ausencia de dominancia, ya que es posible obtener este resultado cuando los efectos aditivos individuales se cancelan entre sí (Ceballos, 1996).
Azizi, F.; Rezai, A.; y Saeidi, G. 2006. Generation mean analysis to estimate genetic parameters for different traits in two crosses of corn inbred lines at three planting densities. J. Agric. Sci. Techn. 8(2):153 - 169. [ Links ]
Ceballos, H. 1996. Manual de Genética Cuantitativa y Mejoramiento Genético. Universidad Nacional de Colombia. Palmira. 337 p. [ Links ]
Cordell, H. 2002. Epistasis: What it means, What it doesn't mean, and Statistical methods to detect it in Humans.. Human Mol. Gen. 11:2463 - 2468. [ Links ]
Cukadar-Olmedo, B. y Miller, J. 1997. Inheritance of the stay green trait in sunflower. Crop Sci. 37:150 - 153. [ Links ]
Eta-Ndu, J. T. y Openshaw, S. J., 1999. Epistasis for grain yield in Two F2 population of maize. Crop Sci. 39:346 - 352. [ Links ]
Fehr, W. R. 1991. Principles of cultivar development. Theory and technique. s.l.:s.n. [ Links ]
Gamble, E. 1962c. Gene effects in corn (Zea mays L.). III. Relative stability of the gene effects in different environments. Canadian J. Plant Sci. 42:626 - 634. [ Links ]
Garcés, A., 2007. Inheritance of Capsaicin and dihidrocapsaicin, determined by HPLC-ESI/MS, in an intraespecific cross of Capsicum annuum L.. En: Estudio de los componentes del caracter picante en pimiento (Capsicum spp.). Zaragosa. Centro de Investigación y Tecnología Agroalimentria. p. 49 - 68. [ Links ]
García, M. A., 2006. Estudio de la diversidad genética de las introducciones de Capsicum spp. del Banco de Germoplasma de la Universidad Nacional de Colombia sede Palmira, Palmira, Universidad Nacional de Colombia sede Palmira. [ Links ]
Kearsey, M. y Pooni, H. 1998. The genetical analysis of quantitative traits. Cheltenham: Stanley Thornes Publ. Ltd. [ Links ].
Lynch, M. y Walsh, B., 1998. Genetics and analysis of quantitative traits. Estados Unidos: Sinauer Associates, inc. [ Links ].
Mather, K. y Jinks, L. 1971. Biometrical Genetics. s.l.:s.n. [ Links ]
Pardey, C., García, M.; y Vallejo, F. 2006. Caracterización morfológica de cien introducciones del Banco de Germoplasma de la Universidad Nacional de Colombia sede Palmira. Acta Agronómica 55(3). [ Links ]
Pardey, C., García, M.; y Vallejo, F. 2009. Evaluación agronómica de Capsicum del Banco de Germoplasma de la Universidad Nacional de Colombia sede Palmira. Acta Agronómica 58(1). [ Links ]
Pardey, C., Posso, A.; y García, M. 2010. Evaluación de accesiones de Capsicum spp por su reacción al virus del mosaico defromante del pimentón (PepDMV). Acta Agronómica 59(1). [ Links ]
Vega, C. O. 1987. Introducción a la teoria de genética cuantitativa, con especial referencia al mejoramiento de plantas. Caracas, Venezuela. Ediciones de la Biblioteca de la Universidad Central de Venezuela. [ Links ]
