










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Agronómica
versão impressa ISSN 0120-2812
Acta Agron. vol.62 no.4 Palmira oct./dez. 2013
Protección de cultivos
1Investigador, estudiante de Maestría en Ciencias Agropecuarias, Universidad Nacional de Colombia sede Palmira. 2Profesora asociada, Universidad Nacional de Colombia, sede Palmira. Facultad de Ciencias Agropecuarias, 3Profesor Titular, Universidad Nacional de Colombia, sede Palmira. Facultad de Ciencias Agropecuarias. Autora para correspondencia: ygarciav@unal.edu.co.
En la Universidad Nacional de Colombia sede Palmira, en condiciones de campo y casa de malla, se evaluaron por preferencia para oviposición y número de ninfas de mosca blanca (Bemisia tabaci (Gennadius)) varias accesiones silvestres y poblaciones segregantes de tomate resistentes al pasador del fruto, Neoleucinodes elegantalis (Guenée). Se utilizó un diseño de bloques completos al azar con cuatro repeticiones, donde se evaluaron como variables el número de huevos y de ninfas del insecto por hoja. En cada observación se incluyeron tres hojas del tercio superior, medio e inferior del tallo de la planta. Las poblaciones de mosca blanca fueron muy bajas en los dos primeros ciclos de cultivo (en campo) debido, probablemente, a las altas precipitaciones. Estas condiciones no permitieron una selección de materiales resistentes o susceptibles a este insecto. En el tercer ciclo de cultivo, bajo condiciones de casa de malla, no se encontraron diferencias significativas entre los retrocruzamientos respecto al testigo susceptible para la variable preferencia por oviposición. El genotipo 4212P4 presentó el menor número de ninfas, mostrando que tiene posiblemente un mecanismo que le confiere menor atracción para B. tabaci. Se encontró que los genotipos menos preferidos para oviposición y con menor número de ninfas fueron PI 134417, PI 134418 y PI 126449 (Solanum habrochaites var. Glabratum S. Knapp y D. M Spooneer).
Palabras clave: Bemisia tabaci, mosca blanca, resistencia, Solanum.
Wild accessions and segregating populations of tomato resistant to Neoleucinodes elegantalis (Guenee) were evaluated in three cycles of cultivation to oviposition preference and number of nymphs of Bemisia tabaci (Gennadius), under field conditions and greenhouse. The experiments were carried out with a randomized complete block design with four replicates. The variables evaluated were: eggs number per leaf and number of nymphs per leaf. Each observation was composed by the mean obtained for three leaves let sof the upper, middle and lower stem. White fly populations were very low in the first two growing seasons due to high rain fall comparable. These conditions did not allow performing a susceptible or resistant material selection to the insect. In the third cultivation cycle under greenhouse conditions were no any significant differences between the back cross with respect to the control variable subject to the oviposition preference. The genotype 4212P4 performed the lowest number of nymphs showing that this genotype possibly has a mechanism that would give less attraction for B. tabaci. Three crop cycles were evaluated under field conditions and greenhouse, all of three evaluations, the genotypes less preferred for oviposition and number of nymphs were: PI 134417, PI 134418 and PI 126449 (Solanum habrochaites var. Glabratum S. Knapp y D. M Spooneer).
Key words: Bemisia tabaci, resistance, Solanum, whitefly.
Bemisia tabaci (Gennadius) fue descrita hace más de 100 años y desde entonces se ha convertido en una de las plagas más importantes en la agricultura del trópico y del subtrópico. Esta especie se adapta fácilmente a las plantas hospederas y a nuevas regiones geográficas y sus biotipos se han identificado en diferentes áreas del mundo, lo cual sugiere que se trata de una especie compleja que cuando es sometida a estrés puede evolucionar. Estos biotipos pueden presentar diferencias en la eficiencia y transmisión de virus, en las tasas desarrollo, en endosimbiontes y en el aprovechamiento y daños fisiológicos causados al hospedero (Oliveira et al., 2001).
En Colombia se confirmó la presencia del biotipo B de B. tabaci atacando 23 especies cultivadas y 11 especies de malezas, mostrando una mayor agresividad que el biotipo A (Quintero et al., 1998). Tanto los adultos como los estados inmaduros de esta plaga ocasionan daños directos a los cultivos al succionar la savia de las plantas, produciendo, además, amarillamiento, encrespamiento y defoliación acompañados de secreción de sustancias azucaradas que recubren las hojas y sirven de sustrato para el crecimiento de hongos de micelio negro (fumagina) pertenecientes a varios géneros, entre ellos Cladosporium y Capnodium, los cuales cubren la parte superior de la hoja e interfieren los procesos de fotosíntesis (Morales et al., 2006).
B. tabaci es la principal plaga que afecta el cultivo de tomate en el valle geográfico del río Cauca, Colombia. Los daños que ocasiona se han incrementado a través del tiempo, pasando a ser considerada una plaga primaria contra la cual se hace uso excesivo de insecticidas (Quirós, 1993); no obstante, muestra resistencia a varios productos usados para su control (Cardona et al., 2001), hasta el punto de que en el futuro los insecticidas de síntesis no serán efectivos para ello (Araya et al., 2005).
La presencia de algunas especies silvestres del género Solanum con resistencia a diferentes insectos plaga (Vallejo, 1999), entre ellas Solanum habrochaites (S. Knapp y D. M Spooneer) que comprende las formas botánicas tipicum y glabratum, caracterizadas por plantas robustas, de gran tamaño, vellosidades muy densas, un fuerte aroma característico, alta densidad de tricomas y resistencia a 14 especies diferentes de insectos plaga, pueden ser utilizadas para el manejo de plagas por introgresión genética de las especies silvestres al tomate cultivado (Simmons y Gurr, 2005), ya que son una fuente natural potencial de genes de resistencia contra la mosca blanca (Sánchez et al., 2006).
En la Universidad Nacional de Colombia sede Palmira, el Programa de Mejoramiento Genético, Agronomía y Producción de Semillas de Hortalizas trabaja actualmente en el desarrollo de una variedad resistente al pasador del fruto (Neoleucinodes elegantalis (Guenée)) a través de introgresión genética a partir de 12 accesiones de las especies silvestres S. habrochaites y S. peruvianum (L.) Mill, y una accesión de tomate S. lycopersicum (L.) que son fuentes de resistencia a diferentes plagas limitantes del tomate, entre ellas B. tabaci (Vallejo, 1999).
En los trabajos realizados por Restrepo (2007) se identificaron tres accesiones silvestres de la variedad glabratum de S. habrochaites (PI 134417, PI134418 y PI 126449) con alta resistencia a N. elegantalis. Estas accesiones se emplearon para obtener poblaciones F1, F2, F3, RC1 y RC2 a partir de cruzamientos interespecíficos con tomate cultivado S. lycopersicum cv. Maravilla (♀). Los trabajos han continuado hasta RC3 con las tres accesiones silvestres y hasta RC4 para la accesión PI134418, utilizando los retrocruzamientos uno (RC1) y dos (RC2), obtenidos en la etapa anterior. Además, se efectuaron cruzamientos en cadena entre plantas resistentes con el fin de incrementar la frecuencia de alelos de resistencia. En los RC3, RC4 y cruzamientos en cadena se logró seleccionar plantas individuales con resistencia al pasador del fruto, que permitirán seguir avanzando hasta lograr cultivares con resistencia, buena producción y calidad de frutos para mercado en fresco (Pérez, 2010).
El Programa de Mejoramiento Genético de la Universidad en Palmira busca producir una variedad de tomate con características ideales agronómicas y con resistencia a diferentes plagas del cultivo, por ello se evaluó en estos materiales resistentes a N. elegantalis la posible resistencia a B. tabaci; por tanto este trabajo tuvo como objetivo evaluar la resistencia a B. tabaci en poblaciones segregantes producidas a partir de cruzamientos entre tomate cultivado S. lycopersicum cv. Maravilla y tres accesiones de la variedad glabratum de S. habrochaites resistentes al pasador del fruto, N. elegantalis.
El estudio se realizó en el campo del Centro Experimental de la Universidad Nacional de Colombia sede Palmira (CEUNP), ubicado en el municipio de Candelaria, Valle del Cauca, corregimiento El Carmelo a 3° 24' norte y 76° 26' oeste, a 980 m.s.n.m., 24 °C de temperatura promedio, 69% HR y 1009 mm de precipitación promedio anual. El estudio en casa de malla tuvo lugar en el corregimiento El Bolo-Italia del municipio de Palmira.
Los genotipos de tomate utilizados fueron: PI 134417, PI 134418, PI 126449 (S. habrochaites var. Glabratum), Unapal-Maravilla (S. lycopersicum), 3143P4 (RC3PI 134417 x Unapal Maravilla), 3247P4 (RC3PI 134418 x Unapal Maravilla), 36110P3 (RC3PI 126449 x Unapal Maravilla) y 4212P4 (RC4PI 126449 x Unapal Maravilla). En campo se realizaron dos ensayos o ciclos de cultivo, correspondientes a ciclos de cultivo, en un diseño experimental de bloques completos al azar con cuatro repeticiones y ocho tratamientos. Cada parcela experimental estaba conformada por siete plantas de cada material, de las cuales cinco centrales se consideraron como parcela útil. En los extremos y en los surcos intercalados de cada parcela se sembraron plantas de tomate cv. Maravilla (testigo susceptible) para garantizar la presencia de mosca blanca en el cultivo. El manejo de plagas y enfermedades del cultivo fue convencional siguiendo sus protocolos de producción, pero sin aplicación de insecticidas desde el trasplante.
Un tercer ensayo se realizó en condiciones de casa de malla, con un diseño experimental de bloques completos al azar y cuatro repeticiones, con siete tratamientos compuestos por las RC3 con las tres accesiones silvestres como padre y S. lycopersicum cv. Maravilla como madre, RC4 con la accesión PI 134418 (♂), Unapal-Maravilla y dos acciones silvestres (PI 134418 PI 126449). Cada parcela estaba conformada por cinco plantas sembradas en recipientes plásticos con capacidad para 12 lt. En los bordes de este ensayo se sembraron plantas de tomate cv. Maravilla preinfestadas con ninfas de cuatro instar de B. tabaci. El manejo de las plagas fue similar al utilizado en los ensayos de campo.
En el primer ciclo de cultivo, la medición de los estados inmaduros de mosca blanca se hizo mediante muestreos semanales en dos plantas de cada genotipo, a partir de la primera semana de trasplante hasta finalizar el ciclo. En el segundo, se hicieron muestreos cada dos semanas en tres plantas de cada genotipo. En casa de malla, cada semana a partir de la segunda semana después de la infestación, se evaluaron cinco plantas de cada genotipo. Para el conteo de individuos durante los tres ciclos, de cada planta se tomó una hoja de los tercios superior, medio e inferior del tallo y en laboratorio con el uso de un estereoscopio se hizo el conteo respectivo de huevos y ninfas.
Se realizó un análisis de varianza mediante el procedimiento GLM de SAS, con el fin de determinar diferencias significativas entre los materiales para el conjunto de variables evaluadas. Cuando la prueba de 'F' fue significativa se hizo la comparación de medias a través de la prueba de Tukey. Todos los datos fueron transformados por Ön + 0.5. Además, se hicieron correlaciones simples entre las diferentes variables utilizadas y las variables de clima en la zona.
En campo. En ambos ciclos de cultivo se encontró baja población de mosca blanca, siendo mayor el número de ninfas que de huevos, especialmente en los tercios medio e inferior de la planta. Los genotipos más preferidos para oviposición fueron Unapal-Maravilla (M) y 36110P3 (36) con promedios de 0.051 y 0.041 huevos/hoja, respectivamente. Para el promedio de ninfas se encontró que el genotipo 3247P4 (32) fue el más afectado por B. tabaci con un promedio de 0.27 ninfas/hoja. Las accesiones silvestres PI 134417, PI 134418 y PI 126449 (P1, P2 y P6) presentaron el menor número de huevos y de ninfas (Figura 1). En este estudio las poblaciones fueron muy bajas, si se comparan con los resultados hallados por Morales y Carmeli (2001) quienes en cultivares de tomate encontraron 12.967 y 6.125 huevos y ninfas/m2, respectivamente.
En la Figura 2 se revela el comportamiento de las poblaciones del insecto de acuerdo con los cambios en precipitación, temperatura y humedad relativa. Para el día 35 después de siembra (DDS) se observó el pico máximo de la población de ninfas con un promedio de 0.39 y una precipitación promedio de 2.13 mm/día; el máximo número de huevos se halló el día 49 DDS con una precipitación promedio de 0.15 mm/día.
Es necesario resaltar que durante el periodo de evaluación las precipitaciones promedio/día fueron altas, lo que afectó la colonización del insecto en los genotipos evaluados; no obstante las correlaciones lineales entre la presencia de huevos y ninfas con las variables de clima no fueron significativas (Cuadro 1) ya que la precipitación no permitió variaciones significativas en humedad relativa (82%) y temperatura (23 °C). Por otro lado, se observó una correlación negativa entre las variables biológicas y la precipitación, lo que tiene efecto sobre la fluctuación de las poblaciones del insecto, al disminuir la presencia de huevos y ninfas.
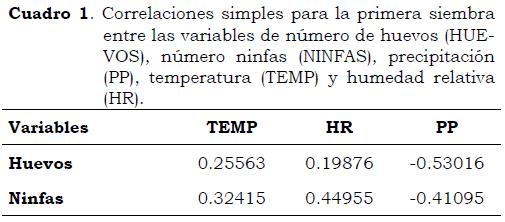
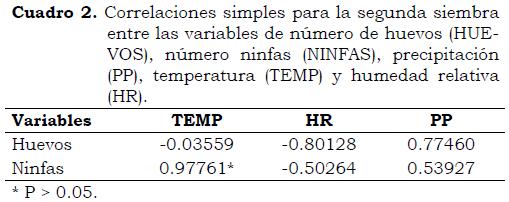
En el segundo ciclo, la incidencia de estados inmaduros de B. tabaci mostró que el genotipo 3247P4 presentó las mayores poblaciones tanto de huevos como de ninfas, mientras que los genotipos PI 134417, PI 134418 y PI 126449 tuvieron las poblaciones más bajas (Figura 3). En la Figura 4 se observa la fluctuación de las poblaciones de estados inmaduros de B. tabaci en este ciclo de cultivo en relación con la precipitación promedio por día. Las poblaciones máximas de huevos y ninfas 30 DDS fueron 0.028 y 0.187, respectivamente, cuando la precipitación promedio fue de 6.8 mm. Entre 45 y 60 DDS el número de huevos y ninfas disminuyó hasta 0.0034 para los primeros y 0.028 para los segundos, con precipitaciones diarias de 6.8 y 7.8 mm/día, respectivamente, y temperatura máxima de 24 °C y mínima de 22 C°. Al igual que en el ciclo anterior de cultivo, las poblaciones del insecto fueron afectadas por las condiciones de clima. En el Cuadro 2 se incluyen las correlaciones simples entre estas variables y muestran que el promedio de ninfas se correlacionó significativamente con la temperatura (R = 0.97), lo que confirma que las altas temperaturas favorecen el desarrollo de las ninfas, como lo demostraron Byrne y Bellows (1991)y Ruiz y Aquino (1999); por el contrario, la alta precipitación es un condicionante de las poblaciones de mosca blanca, ya que cuando la lluvia acumulada durante dos días es > 30 mm la población se reduce drásticamente, lo que muestra que la lluvia es un control natural para esta plaga (Ruiz y Medina 2001). Además de los factores de clima, la mortalidad de huevos y ninfas por parasitoides, fallas en el cuarto instar ninfal, depredación, enfermedades fungosas en las ninfas y mortalidad desconocida, pueden regular las poblaciones de B. tabaci (Byrne y Bellows, 1991).
En casa de malla. Debido a que las condiciones de clima no permitieron hacer una selección inicial de materiales de tomate por resistencia o susceptibles a este insecto, se decidió realizar un tercer ensayo en condiciones de casa de malla. No se encontraron diferencias en ovoposición entre los genotipos 3143P4, 3247P4, 36110P3, 4212P4 vs. el testigo susceptible Unapal-Maravilla (S. lycopersicum), lo que significa que B. tabaci oviposita en forma general en estos genotipos y demuestra que estas plantas no poseen barreras que impidan el ataque del insecto. Las accesiones silvestres PI 134418 y PI 126449 fueron significativamente diferentes con respecto a estos genotipos, presentando promedios de 0.228 y 0.09 huevos/hoja respectivamente (Figura 5). Los genotipos PI 134418 y PI126449 mostraron el menor número de ninfas/hoja (0.049) (P < 0.05) con una tendencia igual para ovoposición, seguido del genotipo 4212P4 con un promedio de 6.59 ninfas/hoja; mientras que Unapal-Maravilla presentó la mayor infestación con un promedio de 12.4 ninfas/hoja (Figura 6). En esta Figura se observa que el menor número de ninfas se presentó en el retrocruzamiento 4212P4, lo que sugiere la existencia de una posible resistencia en este genotipo. Costa et al. (1991) señalan que el número de huevos colocados no es indicativo de la tasa de sobrevivencia. Por otra parte, se han encontrado híbridos entre S. lycopersicum y especies silvestres de S. habrochaites, S. pennellii y S. cheesmanie que producen niveles de resistencia a ácaros e insectos (Simmons y Gurr, 2005). La resistencia de S. habrochaites se debe a su contenido de metabolitos secundarios como zingibereno, 2-cetona, y 2 undecanona y 2 tridecanona segregados por los tricomas glandulares presentes en el follaje (Carter et al., 1989; Eigenbrode et al., 1994; Dimock y Kennedy, 1983; Chatzivasileiadis y Sabelis, 1996) cuyos genes pueden ser recombinados a partir de híbridos interespecíficos entre S. habrochaites y tomate cultivado. Estos efectos de resistencia están controlados por un locus mayor presente en un alelo de S. habrochaites que condiciona el alto contenido de estos metabolitos tóxicos (Freitas et al., 2002). Lo anterior se ha demostrado para el contenido de zingibereno, el cual se mantiene a lo largo de dos retrocruzamientos obtenidos del cruzamiento entre S. lycopersicum y S. habrochaites var. Glabratum, lo que evidencia que es posible obtener plantas resistentes a Tetranychus evansi Baker y Pritchard, seleccionando indirectamente por su alto contenido de zigibereno en las hojas (Gonçcalves et al., 2006).
En los tres ciclos de cultivo en campo y casa de malla y en todas las evaluaciones se observó que los genotipos menos preferidos para la oviposición y número de ninfas fueron PI 134417, PI 134418 y PI 126449 (ver Figuras 1, 3, 5 y 6), resultados que coinciden con los de Fancelli et al. (2003) quienes encontraron que los genotipos LA716, PI134418, PI134417, LA1739, LA1740 y PI127826 fueron menos preferidos para la oviposición debido a la secreción pegajosa producida por los tricomas glandulares que cubren las hojas y tallos de estos genotipos. Estos tricomas son característicos de especies silvestres del género Solanum spp. y son un factor importante para mejoramiento por resistencia a insectos y ácaros (Toscano et al., 2001).
- En ambos ciclos de cultivo en campo, las condiciones de clima entre junio y setiembre de 2010 no permitieron seleccionar genotipos susceptibles o resistentes a B. tabaci.
- En condiciones de casa de malla, los genotipos menos preferidos, según las variables ovoposición y número de ninfas, fueron PI 134417, PI 134418 y PI 126449. Para la variable número de huevos no se encontraron diferencias significativas entre los genotipos derivados de los retrocruzamientos, no obstante el retrocruzamiento 4212P4 fue en el que menor número de ninfas se encontraron.
Araya, L.; Carazo, E.; y Cartin, V. 2005. Diagnóstico del uso de insecticidas utilizados contra Bemisia tabaci (Gennadius) en tomate y chile en Costa Rica. Manejo Integrado Plagas y Agroecol. 75:68 - 76. [ Links ]
Byrne, D. N. y Bellows, Jr. T. S. 1991. Whitefly biology. Annu. Rev. Entomol. 36:431 - 457. [ Links ]
Cardona, C.; Rendón, F.; García, J.; López-Ávila, A.; Bueno, J. M.; y Ramírez, J. 2001. Resistencia a insecticidas en Bemisia tabaci y Trialeurodes vaporariorum (Homóptera: Aleyrodidae) en Colombia y Ecuador. Rev. Col. Entom. 27:33 - 38. [ Links ]
Carter, C.; Sacalis, J.; y Gianfagna, T. 1989. Zingiberene and resistance to Colorado potato beetle in Lycopersicon hirsuturn f. hirsuturn. J. Agric. Food Chem. 37:206 - 210. [ Links ]
Chatzivasileiadis, E.; y Sabelis. M.1996. Toxicity of methyl ketones from tomato trichomes to Tetranychus urticae Koch. Exp. Appl. Acarol. 21(6 - 7):473 - 484. [ Links ]
Costa, H. S.; Brown, J. K.; y Byrne, D. 1991. Life histry traits of the whitefly L. Bemisia tabaci, (Homoptera: Aleyrodidae) on six virus-infected or healthy plant species. Environ. Entomol. 20: 1102 - 1107. [ Links ]
Dimock, M. B. y Kennedy, G. G. 1983. The role of glandular trichomes in the resistance of Lycopersicon hirsutum f.glabratum to Heliothis zea. Entomol. Exp. Appl. 33:263 - 68. [ Links ]
Eigenbrode S.; Trumble, J.; Millar, J.; y White, K. 1994. Topical toxicity of tomato sesquiterpenes to the beet armyworm and the role of these compounds in resistance derived from an accession of Lycopersicon hirsutum f. typicum. J. Agric. Food Chem. 42:807 - 810. [ Links ]
Fancelli, M.; Vendramim J. D.; Lourenção, A.; y Dias, C. 2003. Atratividade e preferência para oviposição de Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) Biótipo B em genótipos de tomateiro. Neotrop. Entomol. 32(2):319 - 328. [ Links ]
Freitas, J.; Maluf, W.; Cardoso, M.; Gomes L.; y Bearzotti, E. 2002. Inheritance of foliar zingiberene contents and their relationship to trichome densities and whitefly resistance in tomatoes. Euphytica 127:275 - 287. [ Links ]
Gonçalves, L.; Maluf, W.; Cardoso, M.; De Resende, J.; De Castro, E.; Santos, N. Rodrigues, I.; y Faria, M. 2006. Relação entre zingibereno, tricomas foliares e repelencia de tomateiros a Tetranychusevansi. Pesq. agropec. Bras. 41(2):267 - 273. [ Links ]
Morales, F.; Cardona, C.; Bueno J.; y Rodríguez, I. 2006. Manejo de integrado de enfermedades de plantas causadas por virus transmitidos por moscas blancas. Centro Internacional de Agricultura Tropical (CIAT), DFID, INISAV, Tropical White Fly IPM Project [ Links ]
Morales, F. y Cermeli M. 2001.Evaluación de la preferencia de la mosca blanca Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) en cinco cultivos agrícolas. Entomotropica Tropical White Fly IPM Project 16(2):73 - 78. [ Links ]
Oliveira, M. R.; Henneberryb, T. J.; y Andersonc, P. 2001.History, current status, and collaborative research projects for Bemisia tabaci. Crop Protection 20:709 - 723. [ Links ]
Pérez, M. 2010. Mejoramiento genético en Solanum lycopersicum para la resistencia al pasador del fruto Neoleucinodes elegantalis Guenée (Lepidoptera: Crambidae). Tesis de maestría. Universidad Nacional de Colombia sede Palmira. Facultad de Ciencias Agropecuarias. [ Links ]
Quintero, C.; Cardona, C.; Ramirez, D.; y Jimenez, N. 1998. Primer registro del biotipo B de Bemisia tabaci (Homoptera: Aleyrodidae) en Colombia. Rev. Col. Entom. 24(1 - 2):23 - 28. [ Links ]
Quiros, C. A. 1993. Adopción y evaluación de la tecnología de semilleros en tomate para el manejo de la mosca blanca, Bemisia tabaci (Gannadius), con participación de los agricultores, en Grecia y Valverde Vega, Alajuela, Costa Rica. Tesis Doctoral. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Programa de Enseñanza Área Posgrado. [ Links ]
Restrepo, E. 2007. Estudios básicos para iniciar la producción de cultivares de tomate Solanum lycopersicum L. con resistencia al pasador del fruto Neoleucinodes elegantalis (Guenée). Tesis Doctoral. Universidad Nacional de Colombia sede Palmira. Facultad de Ciencias Agropecuarias. [ Links ]
Ruiz, J. V. y Aquino, T. B.1999. Manejo de Bemisia tabaci mediante barreras vivas y Paecilomyces en Oaxaca, México. Revista Manejo Integrado de Plagas 52:80 - 88. [ Links ]
Ruiz, J. y Medina, J. Z. 2001. Avances en el manejo integrado de Bemisia tabaci en tomate y chile en Oaxaca, México. Manejo Integrado de Plagas 59:34 - 40. [ Links ]
Simmons, A. y Gurr, G. 2005. Trichomes of Lycopersicon species and their hybrids: effects on pests and natural enemies. Agric. Forest Entom. 7(4):265 - 276. [ Links ]
Snyder, J.; Thackerr. Y.; y Zhang X. 2005.Genetic transfer of a twospotted spider mite (Acari: Tetranychidae) repellent in tomato hybrids. J. Economic Entomol. 98(5):1710 - 1716. [ Links ]
Sánchez, P.; Oyama, J.; Núñez, K.; Fornoni, J.; Hernández, S.; J. Márquez y Garzón, J. 2006. Sources of resistance to whitefly (Bemisia spp.) in wild populations of Solanum lycopersicum var. cerasiforme (Dunal) spooner G. J. Anderson R. K. Jansen in Northwestern Mexico. Genetic Res. Crop Evol. 53:711 - 719. [ Links ]
Toscano, L.; Boiça, J, A.; Maia, J.; y Almeida, J. 2001. Tipos de tricomas em genótipos de Lycopersicon. Hortic. Bras. 19(3):204 - 206. [ Links ]
Vallejo, F. A. 1999. Mejoramiento genético y producción de tomate en Colombia. Palmira: Universidad Nacional de Colombia sede Palmira. p 5,17-22, 45. [ Links ]
