










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Agronómica
versão impressa ISSN 0120-2812
Acta Agron. vol.63 no.2 Palmira abr./jun. 2014
https://doi.org/10.15446/acag.v63n2.38454
http://dx.doi.org/10.15446/acag.v63n2.38454
Agronomía
Respuesta fotosintética de Coffea arabica L. a diferentes niveles de luz y disponibilidad hídrica
Photosynthetic response of Coffea arabica L. to different water and light availabilities
Yesid Alejandro Mariño
Escola Superior de Agricultura Luiz de Queiroz - Universidade de São Paulo, Brazil Autor para correspondencia: yesidmarino@usp.br
Rec.:14.06.2013 Acep.: 02.04.2014
Resumen
El sombrío permite una mejor aclimatación del café (Coffea arabica L.) en zonas donde el invierno se caracteriza por bajas temperaturas nocturnas seguido de días soleados y el verano, por altas temperaturas y lluvias ocasionales. En este estudio, realizado en Viçosa-MG, Brasil, se analizaron los efectos de dos condiciones de luminosidad (pleno sol y 15% de la radiación solar) y disponibilidades de agua en el suelo sobre las relaciones hídricas y el desempeño fotosintético en C. arabica en dos épocas contrastantes. Independientemente de la época de evaluación, las plantas cultivadas en condiciones de baja disponibilidad hídrica presentaron reducciones en la fotosíntesis (A), conductancia estomática (gs) y en la razón Ci/Ca en los dos tratamientos lumínicos. Estos cambios fueron acompañados con la reducción en la conductancia hidráulica y la transpiración. No se presentaron diferencias significativas en la producción cuántica efectiva, en el coeficiente de extinción no-fotoquímica y en la productividad cuántica basal de los procesos no fotoquímicos. Sobre la disponibilidad hídrica adecuada, las plantas cultivadas al sol presentaron reducciones en A, gs y Ci/Ca en la época de invierno. Adicionalmente, se encontraron limitaciones fotoquímicas con la reducción en la razón de las fluorescencias variable e inicial. Los resultados indican que cualquier recomendación de uso de sombrío como práctica de manejo debe ser tomada con precaución.
Palabras clave: Café, estrés hídrico, fotosíntesis, sombra.
Abstract
Shading would allow a better acclimation of coffee plants in response to winter conditions where the nights are relatively cool followed by sunny, warm days and summer conditions with high temperature and rain, in southeastern Brazil. This study aimed to explore the effects of contrasting light conditions combined with two levels of available soil water on water relations and photosynthesis in Coffea arabica L. plants under winter and summer conditions. In both seasons, decreases in photosynthetic rates (A), stomatal conductance (gs) and Ci/Ca ratio were found in droughted plants grown in both the shade and full sun. These changes were accompanied by decreases in transpiration rate and hydraulic conductance. There were no differences in the quantum efficiency of electron transport by photosystem II (PSII), nonphotochemical quenching and non-regulated non-photochemical energy loss in PSII. Compared to shade grown plants, sun-grown plants showed lower A, gs and Ci/Ca ratio under winter conditions. The decrease in the variable-to-initial chlorophyll fluorescence ratio suggests that photochemical events played a role in limitating photosynthesis. The results suggest that any recommended as a cultural management practice should be considered with reserve.
Key words: Coffee, photosynthesis, shading, water stress.
Introducción
El café (Coffea arabica L), es un cultivo nativo de regiones tropicales de África que evolucionó del sotobosque, razón por la cual las primeras plantaciones se establecieron bajo sombra, con el fin de simular su hábitat natural (DaMatta y Rodríguez, 2007). No obstante, las plantas mejoradas de café presentan buena productividad en condiciones a pleno sol, lo que generó a partir de la década de 1950 la eliminación del sombrío en varias regiones en el mundo, incluido Brasil (Da- Matta, 2004a; DaMatta y Rodriguez, 2007). Actualmente, se observa un interés en la reimplantación del sombrío en los cafetales, especialmente en áreas con grandes oscilaciones de temperatura y déficit hídrico, con el fin de utilizar menos insumos y obtener una producción más sostenible (DaMatta y Rena, 2002).
En países productores de café, el déficit hídrico es considerado como la mayor limitación para la producción y está frecuentemente asociado con altas temperaturas (DaMatta, 2004b). El efecto combinado de ambas condiciones estresantes pueden llevar a reducciones en la tasa fotosintética y daños foto-oxidativos en los cafetales (Pinheiro et al., 2004). En ese contexto, el sombrío podría minimizar los impactos negativos del déficit hídrico sobre la fotosíntesis debido a que la menor incidencia de radiación solar causaría un menor déficit de presión de vapor entre la hoja y la atmósfera (DaMatta, 2004a).
La luz y el agua son dos factores que varían e interaccionan, particularmente en ambientes secos, por tanto las respuestas de las plantas a la luz van a estar muy afectadas por la disponibilidad hídrica (Valladares y Pearcy 1997). Cavatte et al. (2012) evaluaron una serie de características morfológicas y fisiológicas en plantas de café y observaron que los efectos de la interacción entre luz y agua fueron independientes. Sobre este efecto no existen estudios comparativos en la zona da Mata Mineira, Brasil, entre sequía y disponibilidad de luz sobre la fisiología del cafeto en condiciones contrastantes, es decir, en el invierno, caracterizado por bajas temperaturas (especialmente en la noche) y días soleados y en el verano, caracterizado por altas temperaturas y lluvias. En el presente estudio, en dos épocas contrastantes del año, se evaluaron el desempeño fotosintético y las relaciones hídricas en plantas de cafeto cultivadas en dos ambientes lumínicos y dos niveles de disponibilidad hídrica.
Materiales y métodos
Material vegetal y condiciones de crecimiento
El estudio se desarrolló en Viçosa (20deg; 45' S, 42deg; 54' O) a 650 m.s.n.m. en el estado de Minas Gerais, Brasil. Se utilizaron plantas de C. arabica cv. Catuaí Vermelho IAC 44 propagadas por semillas y cultivadas en macetas de polipropileno con capacidad de 8 lt. En total fueron cultivadas 48, la mitad de ellas a pleno sol y la otra mitad en ambiente sombreado, aproximadamente 15% de la radiación solar. Después de 6 meses, se tomaron 12 plantas de cada tratamiento lumínico y fueron sometidas a sendos tratamientos hídricos: la mitad de las plantas continuaron con irrigación permanente, recibiendo el 100% del agua evapotranspirada (Capacidad de Campo, CC) y, la otra mitad fue irrigada con cantidades decrecientes de agua transpirada por las plantas a CC (cálculo estimado por gravimetría, con balanza de precisión 0,1 g) hasta alcanzar 35% de humedad disponible en el suelo (plantas con déficit hídrico, DH), durante 76 dias (Dias et al., 2007). El agua disponible fue calculada de los valores de humedad volumétrica en la curva de retención de humedad en el suelo desde CC (-0.010 Mpa) hasta el punto de marchitez permanente (-1.5 MPa). Por tanto, se distribuyeron en cuatro tratamientos (seis plantas por tratamiento) en esquema factorial 2 x 2 (dos tratamientos lumínicos x dos tratamientos hídricos) durante 76 días en cada época, las cuales fueron cosechadas para análisis en época de invierno (junio). Las 24 plantas restantes fueron cosechadas en época de verano (enero). La temperatura al sol fue registrada con un sensor (LI-1400-102, Li-Cor Bioscience Inc., Lincoln, EE.UU.) conectado a un sistema de toma de datos (LI-1400, LI-Cor, Lincoln, EUA), cada minuto y guardando el valor promedio cada 5 min.
Parámetros fotosintéticos
La tasa de fotosíntesis (A), la conductividad estomática (gs) y la razón entre las concentraciones externa e interna de CO2 (Ci/Ca) fueron registrados entre las 09:00 a.m y las 11:00 a.m sobre radiación fotosintéticamente activa (RFA) de 1000 µmol m2/s y una presión parcial de CO2 de 40 Pa con un analizador de gases infrarrojo LI-6400 (LI-COR Biosciences Inc., Lincoln, EE.UU.). Los parámetros de fluorescencia de clorofila a se obtuvieron de las mismas hojas utilizadas para las mediciones de fotosíntesis, usando un fluorímetro portátil modulado (MINI-PAN Walz, Effeltrich, Alemania). La eficiencia cuántica potencial del FSII se calculó después de 30 min de adaptación en la oscuridad, mediante la razón Fv/Fm = (Fm - F0)/Fm, donde F0 es la fluorescencia mínima, excitada por luz roja modulada de baja intensidad (0.03 µmol m2/s) y Fm la fluorescencia máxima obtenida por la aplicación de un pulso de 0.8 s de luz actínica saturante (3000 µmol m2/s). Las hojas fueron aclimatadas a la luz actínica (1000 µmol m2/s) durante 40 s, con el fin de lograr la fluorescencia basal (Fs) y a continuación se aplicó otro pulso saturante para obtener la fluorescencia máxima adaptada a la luz (Fm'). El valor de la fluorescencia mínima adaptada a la luz (Fo') fue calculado según Oxborough y Baker (1997). Con estos parámetros se calcularon la producción quántica efectiva del FSII (ΦFSII), el coeficiente de extinción no-fotoquímica de Stern- Volmer (ΦNPQ) y la productividad cuántica basal de los procesos no fotoquímicos (ΦNO) (Demmig-Adams et al., 1996).
Relaciones hídricas.
El potencial hídrico fue determinado en hojas individuales en las primeras horas del día y al medio día, utilizando una bomba de presión tipo Scholander. La transpiración (E) se determinó por gravimetría según la metodología de Nagakura et al. (2004). La conductividad hidráulica (KL) fue estimada a partir de la transpiración de las plantas entre las primeras horas del día y el medio día y la diferencia del potencial hídrico (ΔΨw) [variación del potencial hídrico entre ambas mediciones (Ψam - Ψmd) observado en ese intervalo según la metodología de Tausend et al. (2000).
Los datos fueron analizados usando un Anova con 2 tratamientos hídricos x 2 tratamientos lumínicos x 2 épocas, siguiendo un diseño completamente aleatorizado y la prueba de 'F' para comparación de medias (P < 0.05), utilizando el programa Sisvar 5.1 Build 7.2.
Resultados y discusión
En general, los valores de la fotosíntesis cambiaron en relación con los tratamientos lumínicos e hídricos en ambas épocas del año. En el invierno, se observó una reducción en los valores de A sobre DH, en mayor extensión en las plantas al sol (75%) que en las plantas a la sombra (63%), comparadas con las plantas irrigadas (Cuadro 1), las que fueron acompañadas de reducciones en gs y en la razón Ci/Ca, lo cual indica que las limitaciones difusivas, en comparación con las limitaciones bioquímicas, en mayor medida afectan la fotosíntesis (Chaves y Oliveira 2004; Oliveira 2009).
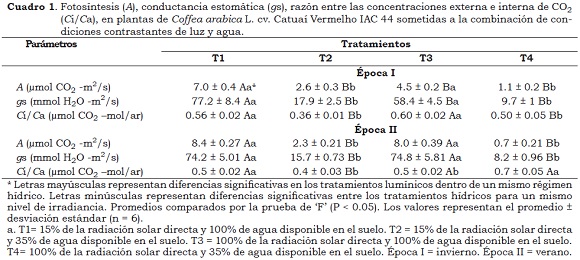
En condiciones de 100% de agua disponible, la reducción de 35% en el valor de A en las plantas cultivadas a exposición de sol, comparadas con las plantas a la sombra, fueron acompañadas por la disminución en el valor gs , sin cambios en la razón Ci/Ca, lo que podría indicar que la fotosíntesis presenta limitaciones no estomáticas. Así mismo, las reducciones en los valores en las razones Fv/Fm (6.4%) y, especialmente en la razón Fv/F0 (24%) en las primeras horas (Figura 1), sugieren la presencia de daños foto-inhibitorios en los centros de reacción del FSII, que afectaron el desempeño fotosintético. Según Maxwell y Johnson (2000) la razón Fv/F0 es un indicador sensible para estimar la foto-inhibición, por tanto, las reducciones en esta relación indican que se presenta una foto-inhibición crónica, es decir, limitaciones fotoquímicas de la fotosíntesis. Resultados similares fueron obtenidos por Silva et al. (2004), Chaves et al. (2008) y Pompelli et al. (2010), quienes anteriormente habían demostrado bajas tasas de fotosíntesis en plantas de cafeto sobre disponibilidad hídrica adecuada en plantas cultivadas al sol, en condiciones de invierno en el sudeste brasilero. DaMatta et al. (1997b) sugieren que las bajas temperaturas nocturnas reducen la conductancia estomática, incluso cuando la temperatura en el día es adecuada para el intercambio gaseoso en las plantas de cafeto.
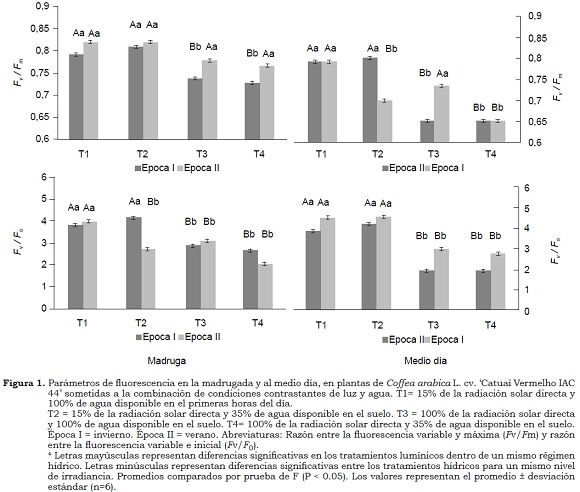
En contraste, en la época de verano a CC se observó un incremento en A (77%) y gs (37%) en las plantas al sol, en relación con la época de invierno (Cuadro 2). Da Matta (2004) sugiere que en la época de verano, el aumento de la humedad relativa a través del día estimula la abertura estomática, cuando la disponibilidad hídrica del suelo no es limitante. En esta misma época con disponibilidad hídrica adecuada no se presentaron diferencias significativas en A y gs en los diferentes tratamientos lumínicos. Adicionalmente, no se encontraron valores < 0.75 en la relación Fv/Fm, a pesar de que las potenciales reducciones en esta relación (plantas/DH en ambos tratamientos lumínicos en el medio día) indican que se presentó foto-inhibición dinámica, asociada con daños fotoquímicos que en relación a foto-protección al aparato fotosintético (Demmig-Adams y Adams, 1992).
Como se sabe, las plantas de cafeto se caracterizan por presentar bajos valores en A, inclusive en óptimas condiciones de cultivo. Posiblemente las altas resistencias estomáticas y mesofílicas en la difusión del C02, desde las cavidades subestomáticas hasta los sitios de carboxilación, explicarían esta respuesta (Silva et al., 2004). Los trabajos de DaMatta (2004a) indican la ocurrencia de altos valores en las tasas de fotosíntesis en un tratamiento lumínico determinado, como resultado de los mayores valores en la conductancia estomática. Los resultados en el presente estudio contrastan con los obtenidos por Cavatte et al. (2012) quienes en época de verano encontraron mayores valores de fotosíntesis en las plantas cultivadas al sol, en comparación con plantas en sombra.
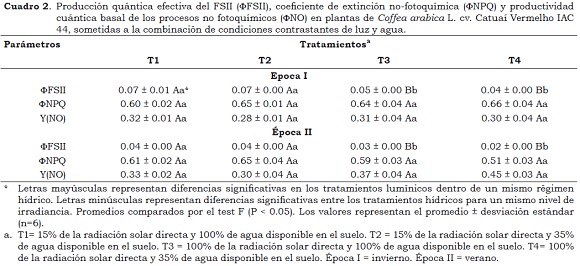
De otro lado, los parámetros fotosintéticos ΦFSII, ΦNPQ y ΦNO, independientemente de los ambientes lumínicos, no respondieron a los tratamientos hídricos en ambas épocas evaluadas (Cuadro 2), lo que confirma estudios previos de DaMatta et al. (1997a) y Praxedes et al. (2006) en los cuales el déficit hídrico no afecta las reacciones fotoquímicas en plantas de cafeto.
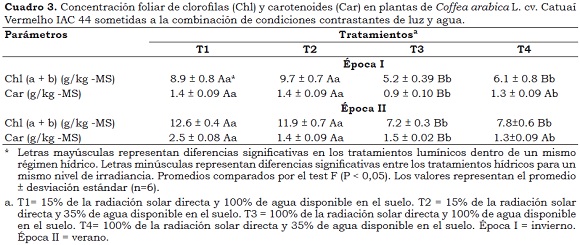
En condiciones de disponibilidad hídrica adecuada, las plantas a la sombra, en comparación con las plantas al sol presentaron un aumento en ΦFSII de 17% y 25% en épocas de invierno y verano, respectivamente. Estos resultados, junto con los aumentos en los valores de la relación Fv/Fm y las concentraciones de los valores de clorofilas y carotenoides (Cuadro 3), son estrategias de las plantas para aumentar su capacidad de absorción a la luz y evitar procesos de foto-inhibición. Para ΦNPQ no se presentaron diferencias (P > 0.05) entre los ambientes lumínicos, no obstante se esperaría una diferencia entre estos valores, debido a la mayor necesidad de disipación de energía en forma de calor para las plantas al sol (Demmig-Adams et al., 1996).
Los parámetros agro-meteorológicos para la colecta en la época de invierno registraron temperaturas máxima de 27.8 deg;C y mínima de 10.3 deg;C; para la época de verano se registraron 30.4 deg;C y 19.8 deg;C, respectivamente. En ambas épocas, las plantas fueron sometidas a deshidratación por 76 días cuando el potencial hídrico en la madrugada (Ψam) alcanzó un valor de -2.0 MPa, valor que representa déficit hídrico moderado (Praxedes et al., 2006). No se encontraron diferencias (P > 0.05) en el Ψmd en las plantas a CC, en ambos ambientes lumínicos; sin embargo en las plantas sobre DH, el Ψmd fue negativo en las plantas a la sombra (Cuadro 4).

En la época de invierno, la reducción en los valores de E sobre DH en ambos ambientes lumínicos, junto con las reducciones en la conductancia estomática, sugieren que C. arabica presenta un eficiente control estomático en la tasa de transpiración en condiciones de déficit hídrico (DaMatta, 2004b). De la misma manera, las reducciones en KL, independiente del ambiente lumínico, se correlacionan con las reducciones en E, debido a que éste es limitado por KL (Tausend et al., 2000). Resultados similares encontraron Dias et al. (2007) y Cavatte et al. (2012), lo que significa conservación de la humedad en el suelo durante un periodo mayor debido a que las plantas absorben el agua con menor intensidad por la menor tensión de la transpiración, que estaría limitada por el cierre estomático.
En contraste, en la época de verano, en las plantas a sol se observó un incremento en E (60%) y KL (61%), en relación con la época de invierno, debido a que las plantas que se desarrollaron en temperaturas más elevadas exigieron una mayor demanda hídrica para su crecimiento y refrigeración. En la época de verano se observaron reducciones en los valores E y KL en condiciones hídricas adecuadas en plantas a la sombra comparadas con las plantas al sol, sin cambios en gs. El sombreamiento puede ser una alternativa eficaz para proteger las plantas contra daños foto-oxidativos, al reducir los extremos de la temperatura y conservar la humedad relativa del aire (DaMatta, 2004a; Beer et al., 1998). En esta condición, el gradiente absoluto de presión de vapor entre los espacios internos de la hoja y el aire disminuye y la transpiración depende más de la resistencia del aire que de la resistencia estomática. Por tanto, aunque las plantas a la sombra presenten una abertura estomática similar para la entrada de CO2 que las plantas al sol, esto no es acompañado por mayor pérdida de agua, en virtud de la contribución de la resistencia del aire para superar la resistencia estomática, optimizando la utilización de agua (DaMatta, 2004a)
Conclusiones
- En la época de invierno, las plantas de cafeto cultivadas a pleno sol, independiente de la disponibilidad hídrica, presentan reducciones en la tasa de fotosíntesis asociada con limitaciones difusivas, debido a bajas tasas de conductancias estomáticas y a limitaciones fotoquímicas, debido a foto-inhibición crónica.
- El sombreamiento permitió una mejor aclimatación de las plantas contra los efectos de las bajas temperaturas nocturnas, reduciendo los efectos negativos del exceso de energía de los días soleados. No obstante, el sombrío no mitigó los impactos de la sequía en el cultivo de café. En la época de verano, los incrementos en la tasa de fotosíntesis en las plantas al sol se deben al aumento de la abertura estomática cuando se presentan buenas condiciones hídricas. En la sombra las reducciones de conductividad hidráulica y transpiración, indican una optimización del uso del agua. De cualquier manera, la recomendación del uso de sombrío como práctica cultural debe ser tomada con precaución.
Agradecimientos
Este trabajo fue financiado por el CNPq, Conselho Nacional de Desenvolvimento Científico e Tecnológico - National Council of Technological and Scientific Development y la Universidad Federal de Viçosa, Brasil.
Referencias
Beer, J.; Muschler, R.; Kass, D.; y Somarriba, E. 1998. Shade management in coffee and cacao plantation. Agrof. Sys. 38:139 - 164. [ Links ]
Cavatte, P. C.; Oliveira, A.; Morais, L. E.; Martins, S. C. V.; Sanglard, L. M., y DaMatta, F. M. 2012. Could shading reduce the negative impacts of drought on coffee? A morphophysiological analysis. Physiol. Plant. 144:111 -122. [ Links ]
Chaves, M. M. y Oliveira, M. M. 2004. Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. J. Exp. Bot. 55:2365 - 2384. [ Links ]
Chaves, A. R. M.; Ten-Caten, A.; Pinheiro, H. A.; Ribeiro, A.; y DaMatta, F.M. 2008. Seasonal changes in photoprotective mechanisms of leaves from shaded and unshaded field-grown coffee (Coffea arabica L.) trees. Trees 22:351 - 361. [ Links ]
Dias, P. C.; Araújo, W. L.; Moraes, G. A. B. K.; Barros, R. S.; y DaMatta, F. M. 2007. Morphological and physiological responses of two coffee progenies to soil water availability. J. Plant Physiol. 164:1639 - 1647. [ Links ]
DaMatta, F. M.; Maestri, M.; y Barros, R. S. 1997a. Photosynthetic performance of two coffee species under drought. Photosynthetica 34:257 - 264. [ Links ]
DaMatta, F. M.; Maestri, M.; Mosquim, P. R.; y Barros, R. S. 1997b. Photosynthesis in coffee (Coffea arabica and C. canephora) as affected by winter and summer conditions. Plant Sci. 128:43 - 50. [ Links ]
DaMatta, F. M. y Rena, A. B. 2002. Ecofisiologia de cafezais sombreados e a pleno sol. En: Zambolim L (ed.). O Estado da arte de tecnologias na produção de café. Editora UFV. Viçosa, Brasil. p. 93 - 135. [ Links ]
DaMatta, F. M. 2004a. Ecophysiological constraints on the production of shaded and unshaded coffee: a review. Field Crop Res. 86:99 - 114. [ Links ]
DaMatta, F. M. 2004b. Exploring drought tolerance in coffee: a physiological approach with some insights for plant breeding. Br. J. Plant Physiol. 16:1 - 6. [ Links ]
DaMatta, F. M. y Rodríguez, N. 2007. Producción sostenible de cafetales en sistemas agroforestales del neótropico. Una visión agronómica y ecofisiológica. Rev. Agron. Col. 25:113 - 123. [ Links ]
Demmig-Adams, B. y Adams, W. W. 1992. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43:599 - 626. [ Links ]
Demmig-Adams, B.; Adams, W. W.; Baker, D. H.; Logan, B. A.; Bowling, D. R.; y Verhoeven, A. S. 1996. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 98:253 - 264. [ Links ]
Maxwell. K. y Johnson, G. N. 2000. Chlorophyll fluorescence a practical guide. J. Exp. Bot. 345:659 - 668. [ Links ]
Nagakura, J.; Shigenaga H.; Akama, A.; y Takahashi, M. 2004. Growth and transpiration of Japanese cedar (Cryptomeria japonica) and Hinoki cypress (Chamaecyparis obtuse) seedlings in response to soil water content. Tree Physiol. 24:1203 - 1208. [ Links ]
Oliveira, A. G. 2009. Aclimatação da maquinaria fotossintética do cafeeiro cultivado em diferentes níveis de luz e de disponibilidade hídrica. Tesis de maestria. Universidade Federal de Viçosa, Viçosa, Brasil. 40 p. [ Links ]
Oxborough, K. y Baker, N. R. 1997. Resolving chlorophyll a fluorescence images of photosynthetic efficiency into photochemical and non-photochemical components calculation of qP and Fv'/ Fm' without measuring F0'. Photosyn. Res. 54:135 - 142. [ Links ]
Pompelli, M.; Martins, S. C.; Antunes, W.; Chaves, A. R.; y DaMatta, F. M. 2010. Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J. Plant Physiol. 167:1052 - 1060. [ Links ]
Pinheiro, H. A.; DaMatta, F. M.; Chaves, A. R. M.; Fontes, E. P. B.; y Loureiro, M. E. 2004. Drought tolerance in relation to protection against oxidative stress in clones of Coffea canephora subjected to long-term drought. Plant Sci. 167:1307 - 1314. [ Links ]
Praxedes, S.C.; DaMatta, F. M.; Loureiro, M. E.; Ferrão, M. A. G.; y Cordeiro, A. T. 2006. Effects of longterm soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ Exp. Bot. 56:263 - 273. [ Links ]
Silva, E. A., DaMatta, F. M.; Ducatti, C.; Regazzi, A. J. y Barros. R. S. 2004. Seasonal changes in vegetative growth and photosynthesis in arabica coffee trees. Field Crop Res. 89:349 - 357. [ Links ]
Tausend, P. C.; Meinzer, F. C.; y Goldstein, G. 2000. Control of transpiration in three coffee cultivars: the role of hydraulic and crown architecture. Trees 14:181 - 190. [ Links ]
Valladares, F. y Pearcy, R. W. 1997. Interactions between water stress, sun-shade acclimation, heat tolerance and photoinhibition in the sclerophyll Heteromeles arbutifolia. Plant Cell Environ. 20:25 - 36. [ Links ]
