










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.63 no.2 Palmira Apr./June 2014
https://doi.org/10.15446/acag.v63n2.30210
http://dx.doi.org/10.15446/acag.v63n2.30210
Protección de cultivos
Evaluación de la resistencia a Prodiplosis longifila Gagné (Diptera: Cecidomyiidae) en genotipos de tomate cultivados y silvestres
Evaluation of resistance to Prodiplosis longifila Gagné (Diptera: Cecidomyiidae) in cultivated and wild tomato genotypes
Yuri Mercedes Mena Pérez *, Nora Cristina Mesa Cobo, Edgar Iván Estrada Salazar, y Yeimy García Valencia
Universidad Nacional de Colombia Sede Palmira, Facultad de Ciencias Agropecuarias. Autora para correspondencia: ymenap@unal.edu.co.
Rec.: 13.06.2012 Acep.: 13.02.2014
Resumen
Se evaluó la resistencia a Prodiplosis longifila Gagné en tres accesiones silvestres de Solanum habrochaites var. glabratum (PI-134417, PI-134418 y PI-126449) y los materiales avanzados en mejoramiento genético para resistencia al pasador del fruto Neoleucinodes elegantalis: tres genotipos de retrocruzamiento 3: RC3(3143P4, 3247P4, 36110P3) y un genotipo de retrocruzamiento 4: RC4(4212P4), como testigo susceptible se incluyó la variedad Solanum lycopersicum cv. Unapal-Maravilla. Las evaluaciones se realizaron en condiciones de campo y de casa de malla, utilizando un diseño de bloques completos al azar con cuatro repeticiones. Se evaluó el número de brotes sanos/planta, número de brotes con daño por P. longifila, número de brotes con presencia de larvas vivas y muertas del insecto, y número de larvas vivas o muertas por brote. Durante el experimento en campo las poblaciones de P. longifila fueron 0.2 larvas vivas/brote y 4.17% de daño, lo cual no permitió el establecimiento de diferencias entre los genotipos estudiados. En casa de malla, las poblaciones del insecto fueron elevadas, sin embargo las accesiones silvestres no mostraron daño ni fueron preferidas para la oviposición, presentando algún grado de resistencia. Unapal-Maravilla y los genotipos correspondientes a la RC3, fueron susceptibles al insecto. El genotipo 4212P4 mostró el menor daño de las poblaciones segregantes con solo 5.36% de brotes dañados, siendo considerado como de menor preferencia por el insecto plaga. Los genotipos provenientes de retrocruzamientos evaluados en estos ensayos fueron previamente seleccionados por resistencia a N. elegantalis, y en este procedimiento es posible que hayan podido descartarse genotipos con resistencia a P. longifila.
Palabras clave: Prodiplosis longifila, prodiplosis, resistencia a plagas, Solanum lycopersicum, Solanum habrochaites.
Abstract
Resistance was evaluated Prodiplosis longifila Gagné in three wild accessions Solanum habrochaites var. glabratum (PI134417, PI134418 and PI 126449) and advanced materials in breeding for resistance to fruit borer Neoleucinodes elegantalis: three genotypes of three backcross RC3 (3143P4, 3247P4, 36110P3) and four backcross genotype RC4 (4212P4) as susceptible check variety S. lycopersicum cv. Unapal-Maravilla. Evaluations were performed under field conditions and screen house, using a completely randomized design with four replications. Number of healthy/plant, number of shoots sprout damage was assessed with P. longifila, number of shoots with the presence of live and dead and number of living and dead larvae larvae per shoot. During the experiment field populations of P. longifila were 0.2 live larvae/shoot and 4.17 % damage, which prevented the establishment of differences between the genotypes studied. In screen house, insect populations were high, however wild accessions showed no damage, nor were preferred for oviposition, showing some degree of resistance. Unapal-Maravilla and genotypes corresponding to the RC3, were susceptible to insect. 4212P4 genotype showed the least damage of segregating populations with only 5.36% of shoots damaged, regarded as less preferred by the insect pest. Genotypes from backcrosses evaluated in these trials had previous selection toward resistance N. elegantalis, and in this process they may have been discarded genotypes with resistance to P. longifila.
Key words: Prodiplosis longifila, prodiplosis, resistance, Solanum lycopersicum, Solanum habrochaites.
Introducción
Prodiplosis longifila (Gagné), comúnmente denominado prodiplosis, negrita o caracha, se encuentra entre los principales problemas fitosanitarios limitantes para el cultivo del tomate (Solanum lycopersicum L.). Esta plaga es una mosca neotropical, pequeña y polífaga, considerada de importancia económica por los graves daños que causa, ya que afecta severamente los brotes tiernos, las inflorescencias y los frutos pequeños, deformándolos y limitando la producción de tomate. Su presencia ha sido confirmada en Colombia, Ecuador y Perú, países donde el daño es producido por las larvas en cogollos y tejidos tiernos (Valarezo et al., 2003). Delgado (1998) documentó la presencia del díptero P. longifila en las diferentes zonas tomateras del Valle del Cauca, siendo actualmente una de las plagas más limitantes desde el punto de vista económico para dicho cultivo, donde genera pérdidas hasta de 60% de la producción (Valarezo et al., 2003).
La complejidad de prodiplosis ha obligado a la mayoría de los productores de tomate al uso indiscriminado de insecticidas químicos que promueven un alto grado de contaminación en el ambiente y en los frutos, lo que conlleva deterioro del equilibrio biológico, desarrollo de resistencia, pérdida de rentabilidad y por último, abandono del cultivo. Además, los hábitos del insecto permiten su presencia a través del desarrollo fenológico del cultivo (Delgado, 1998). Lo anterior limita las opciones de manejo y obliga a la búsqueda de resistencia varietal como la mejor alternativa para la protección del cultivo.
Vallejo (1999) considera que es difícil encontrar resistencia a insectos plagas en las formas cultivadas de S. lycopersicum; no obstante en especies silvestres como S. habrochaites var. glabratum, se ha identificado resistencia a la mayoría de los artrópodos plagas en tomate (Parra et al., 1993; Nippes, 2005) y en particular al cogollero Tuta absoluta (Meyrick) por la posible asociación de la resistencia con la presencia de metabolitos secundarios presentes en la planta entera, inclusive en los frutos inmaduros.
En el Programa de Investigación de Mejoramiento Genético, Agronomía y Producción de Semillas de Hortalizas, de la Universidad Nacional de Colombia sede Palmira, Restrepo (2007) encontró alta resistencia al pasador del fruto (Neoleuciones elegantalis (Guenée) (Lepidoptera: Crambidae)) en 12 accesiones de la especie silvestre S. habrochaites. El método de retrocruzamiento permitió producir poblaciones recurrentes, a partir de cruzamientos entre tomate cultivado y las accesiones silvestres resistentes al pasador. Pérez (2010) efectuó cruzamientos en cadena entre plantas resistentes, con el fin de incrementar la frecuencia de alelos de resistencia. En las generaciones RC3, RC4 y cruzamientos en cadena lograron seleccionar plantas individuales con resistencia al pasador del fruto.
Frente a la susceptibilidad del cultivo de tomate y la necesidad de proponer soluciones a los severos daños ocasionados por P. longifila, la identificación de fuentes de resistencia a este insecto constituye un avance para la obtención futura de cultivares resistentes, lo que permitirá a los agricultores acceder a un control limpio, ecológico y duradero del insecto plaga. Con este propósito en este trabajo se evaluó la resistencia a P. longifila en accesiones silvestres de S. habrochaites y en poblaciones recurrentes derivadas de cruzamientos interespecificos, seleccionadas previamente por su resistencia al pasador del fruto N. elegantalis.
Materiales y métodos
Evaluación de germoplasma en campo.
El trabajo se realizó en el Centro Experimental de la Universidad Nacional de Colombia sede Palmira (3° 21'N y 76° 18'O, 1024 m.s.n.m., 23 °C y 1000 mm). Se evaluaron ocho genotipos, entre ellos: tres accesiones de la especie silvestre S. habrochaites var. glabratum; tres genotipos de retrocruzamiento 3 (RC3) producidos a partir de cruzamientos entre tomate cultivado S. lycopersicum cv. Maravilla (♀) y las accesiones silvestres PI 134417, PI 134418 de S. habrochaites var glabratum (♂) y 1624 de S. habrochaites var. Typicum (♂); un genotipo de retrocruzamiento 4 (RC4) con la accesión PI 134418, resistente al pasador del fruto N. elegantalis; y una variedad de tomate S. lycopersicum cv. Unapal-Maravilla, la cual se usó como testigo susceptible a prodiplosis (Cuadro 1). La semilla utilizada para las diferentes evaluaciones fue suministrada por el Programa de Investigación de Mejoramiento Genético, Agronomía y Producción de Semillas de Hortaliza de la Universidad Nacional de Colombia sede Palmira, la cual fue obtenida en el Proyecto: Estudios Básicos para Iniciar la Producción de Cultivares de Tomate S. lycopersicum con resistencia al pasador del fruto N. elegantalis.

Se utilizó un diseño experimental de bloques completos al azar de la forma: Yij = µ + τi + βj + Eij con ocho tratamientos y cuatro repeticiones; cada parcela experimental estaba conformada por cinco plantas. Para eliminar el efecto de bordes, en el perímetro de los bloques se establecieron plantas de tomate Unapal-Maravilla (susceptible a prodiplosis) y para garantizar una fuente adecuada de insectos en el campo se sembraron surcos intercalados entre las parcelas de este mismo cultivar susceptible.
Para evaluar la susceptibilidad de los genotipos al daño por prodiplosis, a partir de la primera semana después del trasplante en campo, se evaluaron: el número de brotes sanos/planta, el número de brotes con daño por P. longifila, el número de brotes con presencia de larvas vivas y muertas, el número de larvas vivas o muertas por brote (Foto 1). Estas evaluaciones se hicieron durante dos ciclos del cultivo (junio - septiembre y octubre - diciembre de 2010). En ambos ciclos se realizó un manejo convencional del cultivo que incluyó fertilización y control de enfermedades y arvenses (malezas).

Evaluación del germoplasma en casa de malla.
El trabajo se realizó en el vivero Hawai en el corregimiento El Bolo, municipio de Palmira, Valle del Cauca. Se evaluaron siete de los ocho genotipos previamente estudiados en la etapa de campo (Cuadro 1). El genotipo correspondiente a la accesión silvestre PI134417 no pudo ser evaluado por su escasa germinación. Las plantas fueron sembradas en recipientes plásticos, con un volumen de 12 lt, con un diseño similar al utilizado en la etapa de campo.
Las liberaciones de pupas de prodiplosis comenzaron cinco días después del trasplante. Estas fueron recolectadas en campos infestados y mantenidas en condiciones de laboratorio hasta su liberación uniforme en la casa de malla. Las liberaciones se realizaron hasta 23 días después del trasplante de las plántulas, alcanzando más de mil pupas con una relación machos:hembras de 1:1 durante este periodo.
La evaluación de la susceptibilidad de los genotipos de tomate al daño por prodiplosis se hizo semanalmente en todas las plantas, e incluyó los mismos parámetros medidos para el caso del ensayo en campo. Estas evaluaciones se realizaron solo durante la etapa vegetativa y de floración del cultivo.
Análisis de datos.
Los resultados obtenidos, tanto en campo como en casa de malla, fueron sometidos a análisis de varianza (Anova); cuando la prueba de 'F' mostró diferencias significativas entre tratamientos se hizo una prueba de comparación de medias (Duncan), utilizando el paquete Statistical Analysis System SAS versión 9.0. Para disminuir la falta de continuidad de la información, los datos fueron transformados usando arcoseno √variable de respuesta + 0,1. Se calcularon las correlaciones simples entre las diferentes variables utilizadas y las variables climáticas de la zona. La información de clima fue suministrada por la estación meteorológica del Centro de Investigación de la Caña de Azúcar de Colombia (Cenicaña), localizada en el corregimiento de San Antonio de los Caballeros (Florida, Valle del Cauca).
Resultados y discusión
En campo
En general, las poblaciones en campo de P. longifila en ambos ciclos del cultivo fueron muy bajas y sólo algunas variables mostraron diferencias significativas entre los genotipos evaluados. El cultivar Unapal-Maravilla presentó, en promedio, 6.44% de brotes con daños en el primer ciclo de cultivo y 4.34% en el segundo. La población recurrente RC3 36110P3 mostró porcentajes iguales de brotes dañados que el cultivar Unapal-Maravilla y los demás genotipos evaluados. En el segundo ciclo de cultivo, la población RC4 4212P4 presentó un comportamiento similar a los demás genotipos evaluados en relación con las variables brotes con larvas vivas (%) y larvas vivas por brote. La ausencia de larvas vivas y de daños en brotes, indica que las tres introducciones silvestres evaluadas no fueron preferidas para ovipositar por parte de los adultos de prodiplosis, por tanto, éstas se pueden considerar no preferidas por el insecto para ovipositar o alimentarse (Cuadro 2). Vallejo et al. (2008) encontraron que en las introducciones PI- 134417, PI–134418 y PI-126449 de la var. glabratum los frutos no fueron afectados por el insecto; igualmente observaron que algunos frutos de estas introducciones silvestres tenían posturas de pasador (N. elegantalis), pero sin presencia de daños.
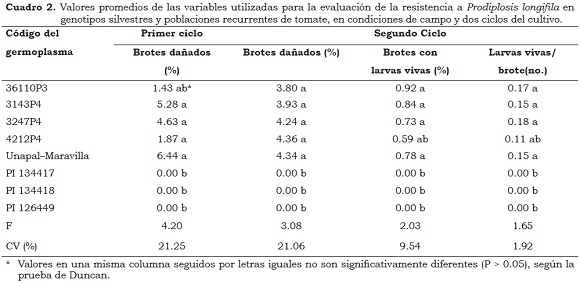
En el primer ciclo del cultivo no se hallaron diferencias (P < 0.05) entre los genotipos evaluados para las variables brotes con presencia de larvas vivas (%) y número promedio de larvas vivas por brote. Por el contrario, en el segundo ciclo se presentaron diferencias altamente significativas (P < 0.001) entre los ocho genotipos evaluados para el conjunto de variables evaluadas para resistencia a prodiplosis; así el genotipo 4212P4 mostró 4.36% de brotes con daño, 0.59% de brotes con larvas vivas y 0.11 larvas vivas/brote (Cuadro 2).
Las accesiones de S. habrochaites var. glabratum (PI-134417, PI–134418 y PI-126449) no presentaron daño por P. longifila en comparación con las poblaciones recurrentes, correspondientes a los retrocruzamientos 3 y 4 y al testigo susceptible cv. Unapal-Maravilla, en ambos ciclos del cultivo.
Los estudios sobre los mecanismos de resistencia del cultivo de tomate a P. longifila son escasos, sin embargo, hay algunas investigaciones realizadas por Kennedy (2003) sobre el mecanismo de resistencia a otras plagas en este cultivo, lo que está condicionado con factores asociados a los tricomas glandulares que abundan en el follaje y que contienen la toxina 2-tridecanona. La accesión PI-134417 de tomate silvestres L. hirsutum cv. glabratum fue seleccionada previamente por su alto nivel de resistencia a Manduca sexta L. y Heliothiszea (Boddie), además de ser resistente al escarabajo de la patata (Leptinotar sadecemlineata Say). Restrepo (2007) cita que las tres accesiones evaluadas de la variedad glabratum de S. habrochaites (PI-134417, PI–134418 y PI-126449) presentaron alta resistencia al pasador del fruto de tomate (N. elegantalis) ya que los frutos no revelaron orificios de salida de larvas ni presencia de éstas dentro del fruto. Anteriormente Salinas et al. (1993) habían reportado una introducción de S. habrochaites var. glabratum que no presentaba frutos afectados por pasador y que clasificaron como altamente resistente al ataque del insecto.
Al relacionar la infestación -daño y larvas vivas/brote- con la precipitación, se encontró que los mayores porcentajes de infestación se presentaron en la época seca, con temperatura promedio de 24 °C entre 49 y 77 días después del trasplante de las plántulas, no obstante, los valores no sobrepasaron el umbral de acción (10% y 20% de brotes infestados con larvas vivas) propuesto por Valarezo et al. (2003). Esto muestra que el nivel de población pudo haber afectado una mejor expresión de la resistencia para la selección de plantas en los diferentes germoplasmas. Dahms (1992) menciona como uno de los requisitos para realizar la evaluación de la resistencia a insectos, el uso de niveles de infestación uniformes y controlados, ya que las infestaciones bajas no son confiables y pueden presentar 'escapes'.
Durante el primer ciclo del cultivo (junio - septiembre de 2010) el porcentaje de infestación fue muy bajo y alcanzó un valor máximo de 0.75%, lo cual muestra la influencia de la época de lluvias. Por otra parte, el promedio de larvas vivas/brote y el porcentaje de daño fueron nulos durante la época de lluvias, alcanzando valores de cero para ambas variables. El mayor valor lo hubo en la época seca entre los 56 y 77 días después de la siembra, presentando un promedio de 0.36 larvas/brote y un porcentaje de brotes con daño de hasta 5.64% (Figura 1).

La temperatura y la humedad relativa promedios en el primer ciclo del cultivo fueron, respectivamente, 22.68 ± 0.43 °C y 81.80 ± 2.98%. Estas variables tienen una influencia importante tanto en el cultivo como en el insecto, aunque la temperatura promedio se encontraba entre los rangos óptimos para el insecto, la humedad relativa alcanzó en el extremo del rango máximo (Díaz, 2009 ).
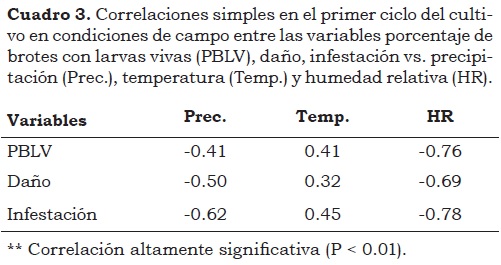
Las correlaciones lineales entre porcentaje de larvas vivas por brote (PLVB) vs. precipitación (PRECI) y humedad relativa (HR) fueron de -0.45 y -0.76, respectivamente, por tanto, al incrementar estas variables climáticas, las larvas disminuyen y consecuentemente el daño ocasionado a la planta es menor. Es posible que estos factores (precipitación y humedad relativa) sean los responsables del descenso de las poblaciones de P. longifila durante el primer ciclo de evaluación. Valarezo et al. (2003) observaron que los días con baja luminosidad y alta humedad relativa, así como los días calurosos con temperaturas medias alta, favorecen el desarrollo de P. longifila. Es importante tener en cuenta que en 2010-11, cuando se desarrolló la presente investigación, ocurrió el fenómeno de La Niña (Cuadro 3), caracterizado por una alta precipitación.
Para el segundo ciclo del cultivo (octubre - diciembre de 2010) los resultados fueron similares a los del primer ciclo. El porcentaje de infestación por el insecto fue muy bajo durante todo el periodo con variación entre 0.03% y 1.2 %. Los promedios más altos de larvas vivas/brote (0.2) y porcentajes de daño (4.17%) se presentaron en la época seca. Los promedios y porcentajes disminuyeron durante la época lluviosa después de 35 días de trasplante en campo (Figura 2). En este segundo ciclo, los promedios de temperatua y humedad relativa fueron de 22.79 ± 0.83 °C y 79.93 ± 3.58%, respectivamente. En este caso, aunque la temperatura se encontraba en el rango óptimo, la humedad relativa se hallaba en el límite del rango máximo (Díaz, 2009).
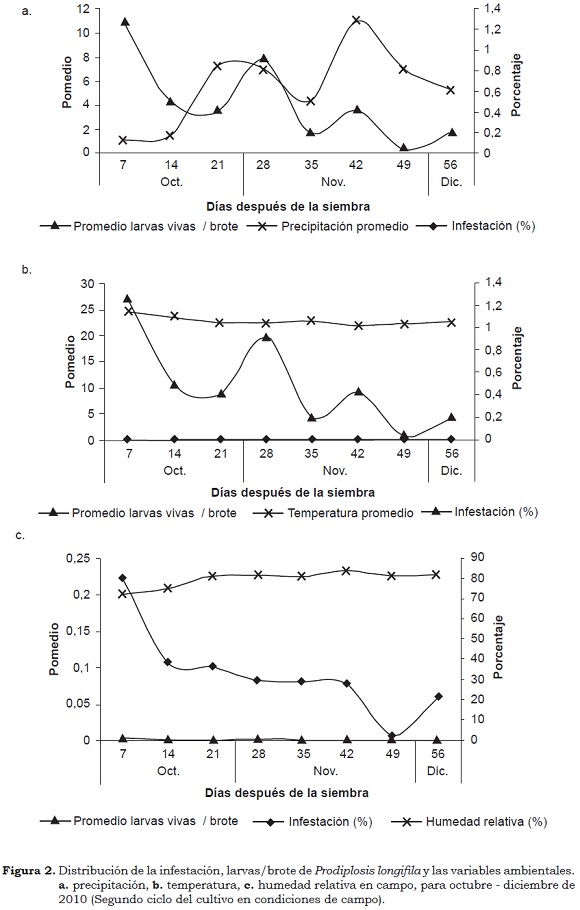
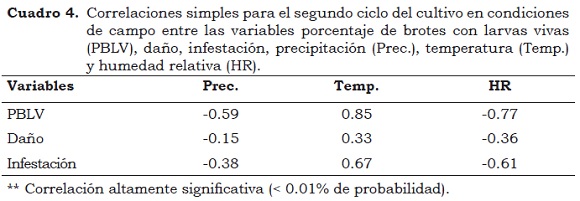
Al igual que en el primer ciclo del cultivo, el porcentaje de larvas vivas/brote (PLVB) presentó una asociación negativa con la precipitación (Prec.) y la humedad relativa (HR), lo que sugiere que a mayor precipitación y humedad relativa se presenta un menor porcentaje de larvas vivas/brote, entre el primero y el tercero instar larval, lo que se refleja en una baja infestación y en menor daño (Cuadro 4). Valarezo et al. (2003) consideran que la época de mayor ataque de la plaga varía con el clima en cada zona de cultivo; los meses secos son los más críticos y en los lluviosos la plaga disminuye drásticamente su ataque. Haddad y Pozo (1994) consideran que la alta temperatura y la humedad relativa favorecen la formación de una nueva generación del insecto cada 7 - 10 días y que en condiciones opuestas, el estado de pupa se prolonga.
En casa de malla
En estas condiciones, se presentaron diferencias (P < 0.05) entre los genotipos evaluados para el porcentaje de brotes/planta dañados. Los genotipos silvestres de S. habrochaites var. glabratum (PI-134417, PI–134418 y PI-126449) no mostraron daño por prodiplosis; mientras que el testigo Unapal-Maravilla y el genotipo 3143P4 presentaron los mayores porcentajes de daño, siendo de 8.26% y 7.74%, respectivamente. El genotipo 4212P4 mostró el menor daño en brotes (5.36%) en comparación con los materiales segregantes, aunque este valor es muy próximo al del testigo susceptible (Cuadro 5). Por otra parte, las accesiones silvestres no revelaron daños en brotes por larvas vivas, mientras que el cultivar Unapal- Maravilla y el genotipo3143P4 mostraron los mayores porcentajes de infestación -2.07% y 2.03%, respectivamente. Los demás genotipos (3247P4, 36110P3, 4212P4) mostraron valores estadísticamente iguales. En cuanto a las larvas vivas/brote, las accesiones silvestres no fueron atractivas ni preferidas para la ovoposición por lo que no presentaron larvas vivas en brotes.
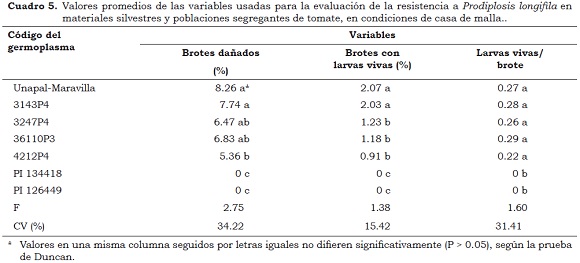
Las plantas de las accesiones silvestres PI-134417, PI–134418 y PI-126449 de la variedad glabratum de S. habrochaites no presentaron brotes afectados por prodiplosis ni fueron preferidas para ovoposición, lo que muestra algún grado de resistencia; por el contrario, el cultivar Unapal Maravilla de S. lycopersicum y los materiales 3143P4, 3247P4, 36110P3 tuvieron un comportamiento similar al testigo susceptible y presentaron daños entre 5.36% y 8.26% de brotes afectados, por lo que fueron clasificados como susceptibles al insecto. El genotipo 4212P4 tuvo menos daño y puede ser clasificado como moderadamente resistente o con algún grado de resistencia a prodiplosis.
El germoplasma 4212P4 conservó un comportamiento similar (P > 0.05) a los demás genotipos recurrentes y al testigo comercial en relación con el número de larvas vivas/ brote y consecuentemente con un menor daño. Se considera que prodiplosis presentó igual preferencia para ovipositar en todos los genotipos y menor preferencia para alimentarse en este genotipo, lo cual indica que el genotipo 4212P4 puede ser seleccionado por la presencia de algún grado de resistencia, que debe ser evaluado con mayor intensidad para determinar la posible expresión de antibiosis como mecanismo de resistencia, considerando los efectos particulares en la biología del insecto.
Conclusiones
- Las accesiones de tomate silvestres PI- 134417, PI–134418 y PI-126449 de la variedad glabratum de S. habrochaites no presentaron brotes afectados por prodiplosis, por lo que se consideran con algún nivel de resistencia.
- El cultivar Unapal-Maravilla de S. lycopersicum y los genotipos correspondientes a la RC3 (3143P4, 3247P4 y 36110P3) fueron susceptibles o muy susceptibles a P. longifila; mientras que el genotipo 4212P4 fue el menos preferido por este insecto.
- El genotipo de tomate correspondiente a la RC4 (4212P4) fue clasificado como el menos preferido por P. longifila y presentó el menor daño.
- Las expresiones de resistencia observadas en estudio deben ser confirmadas en otros estudios, ya que las bajas poblaciones del insecto plaga no permitieron una mejor y más segura comparación entre los cultivares.
Referencias
Delgado, A. 1998. Biología y evaluación de métodos de manejo de Prodiplosis longifila Gagné (Diptera:Cecidomyiidae) en un cultivo de tomate Lycopersicon esculentum del Valle del Cauca. Tesis pregrado. Universidad Nacional de Colombia, Palmira. [ Links ]
Dahms, R. G. 1972. Techniques in the evaluation and development of host plant resistance. J. Environ. Quality, 1: 254-259. [ Links ]
Díaz, F. 2009. Manejo Integrado de Prodiplosis longifila en Perú. Disponible en: http://ffernandodiazs.galeon.com/index.html. [11-03-2012] [ Links ]
Haddad, R. y Pozo, M. 1994. Morfologia y comportamiento de Prodiplosis sp. en papa, Solanum tuberosum. 36 Convencion Nacional de Entomologia: Resúmenes y Programas. Tarapoto, Perú. p. 17. [ Links ]
Kennedy, G. 2003. Tomato, pests, parasitoids, and predators: Tritrophic Interactions Involving the Genus Lycopersicon. Annu. Rev. Entomol. 48:51 - 72. [ Links ]
Nippes, G. 2005. Seleção para alto teor de acilaçúcares em genótipos de tomateiro e sua relação com a resistência ao ácaro vermelho (Tetranychus evansi) e à traça (Tuta absoluta). Universidade Federal de Larvas. Minas Gerais-Brasil. Trabajo de grado (Doctorado). Disponible en: http://www.prpg.ufla.br/genetica/Disserta%E7%F5es%20e%20Teses/Tese%20-%20Guilherme%20Victor%20Nippes%20Pereira.pdf. [10-09-2009] [ Links ].
Parra, A.; Barona, G.; y Vallejo, F. A. 1993. Evaluación de especies silvestres de Lycopersicon sp. como fuente de resistencia al insecto plaga Scrobi palpula absoluta (Meyrick) y su intento de transferencia a la especie cultivada Lycopersicon esculentum Mill. Tesis pregrado. Universidad Nacional de Colombia, Palmira. [ Links ]
Pérez, M. 2010. Mejoramiento genético en Solanum lycopersicum para la resistencia al pasador del fruto Neoleucinodes elegantalis Guenée (Lepidoptera: Crambidae). Tesis Magíster. Universidad Nacional de Colombia, Palmira. [ Links ]
Restrepo, E. 2007. Estudios básicos para iniciar la producción de cultivares de tomate Solanum lycopersicum L. con resistencia al pasador del fruto Neoleucinodes elegantalis (Guenée). Tesis Doctor. Universidad Nacional de Colombia, Palmira. [ Links ]
Salinas, A. H.; Estrada, E. I.; y Vallejo,F. A. 1993. Evaluación de la resistencia al pasador del fruto del tomate Neoleucinodes elegantalis (Guenée) en materiales de Lycopersicum hirsutum Hum y Bonpl y Lycopersicum pimpinellifolium (Just) Mill y su transferencia a materiales cultivados de tomate L. esculentum Mill. Tesis Maestría. Universidad Nacional de Colombia., Palmira. 106 p. [ Links ]
Thomazini, A.; Vendramim, J.; Brunherotto, R.; y Lopes, M. 2001. Efeito de Genótipos de tomateiro sobre a biología e oviposição de Tuta absoluta (Meyrick) (Lep.:gelechiidae). Neotrop. Entomol. 30(2). [ Links ]
SAS Institute Inc. 2009. Statistical Analysis System user's guide. Version 9.0. Cary, Statistical Analysis System Institute. [ Links ]
Thomazini, A.; Vendramim, J.; Brunherotto, R.; Lopes, M. 2001. Efeito de Genótipos de tomateiro sobre a Biología e Oviposição de Tuta absoluta (Meyrick) (Lep.:gelechiidae). Neotropical Entomology. vol.30 no.2 Londrina June 2001. Print version ISSN 1519-566X. Disponible en: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1519-566X2001000200012. [10-09-2009] [ Links ]
Valarezo, O.; Cañarte, E.; Navarrete, B.; y Arias, M. 2003. Prodiplosis longifilia (Diptera:Cecidomyiidae) principal plaga del tomate en el Ecuador. Diagnóstico, Bioecología y Manejo. Iniap, Promsa y Cedege. Manual no. 51. [ Links ]
Vallejo, A.; Restrepo, E.; y Lobo, M. 2008. Resistencia al perforador del fruto del tomate derivada de especies silvestres de Solanum spp. Rev. Fac. Nal. Agr. Medellín 61(1):4316 - 4324. [ Links ]
