










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.64 no.3 Palmira July/Sep. 2015
https://doi.org/10.15446/acag.v64n3.38033
http://dx.doi.org/10.15446/acag.v64n3.38033
Macrofauna edáfica asociada con sistemas agroforestales en la Amazonía Colombiana
Soil macrofauna associated to agroforestal systems in Colombian Amazon
Juan Carlos Suárez Salazar *, Ervin Humprey Duran Bautista y Gelber Rosas Patiño
Universidad de la Amazonía, Facultad de Ingeniería, Programa Ingeniería Agroecológica. Florencia - Caquetá. Colombia. *Autor para correspondencia: juansuarez1@gmail.com
Rec.: 28.02.2014 Acep.: 28.03.2015
Resumen
La densidad, riqueza y diversidad de la macrofauna edáfica son afectadas, entre otros factores, por la configuración de los agroecosistemas y la estacionalidad de la precipitación. Con el fin de evaluar estos efectos en mayo y noviembre de 2011, correspondientes a épocas de máxima y mínima precipitación, respectivamente, se realizaron estudios sobre la composición de la macrofauna edáfica asociada con los arreglos agroforestales ubicados en el Centro de Investigaciones Macagual Cesar Augusto Estrada González, Amazonia, Caquetá, Colombia. El experimento se dispuso en un diseño completo al azar bifactorial con cuatro tratamientos (arreglos agroforestales: AB = abarco - Cariniana pyriformis Miers; CH = caucho Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. ; CP = caucho-parica Schizolobium amazonicum Huber; UV = uvito Genipa Americana L.) y dos épocas (máxima y mínima precipitación), y cuatro repeticiones en parcelas divididas. Para explorar las relaciones entre los órdenes de macrofauna, se realizó un análisis de componentes principales y se evalúo el efecto de los arreglos agroforestales con una prueba de Monte Carlo. Los resultados mostraron que la densidad de la macrofauna fue mayor en el periodo de máxima precipitación (1129 individuos) en comparación con el de mínima (598 individuos). Los arreglos agroforestales influyen sobre la presencia o ausencia de algunos grupos taxonómicos (P < 0.05) como Homoptera (Insecta) y Raphidioptera (Insecta); además los UV y AB pueden favorecer a la macrofauna del estrés por sequía.
Palabras claves: Edafofauna, fauna del suelo, arreglo agroforestal, época, densidad, riqueza, diversidad, Amazonía.
Abstract
The density, richness and diversity of soil macrofauna are influenced by diverse factors such as the configuration of agroecosystems, seasonality, among others. In order to assess this influence, sampling was conducted in May and November 2011, corresponding to the periods of maximum and minimum rainfall respectively on the composition of soil macrofauna associated with agroforestal arragements located in the Research Centre Macagual Cesar Augusto Estrada González property of the University of Amazonia, Colombia. The experiment was conducted under a complete randomized bifactorial design with 4 treatments (agroforestry system) and 2 periods (maximum and minimum precipitation), with 4 replicates in split plots. Principal component analysis was performed to explore the relationships between the orders and the effect of the agroforestal arrangements, which was tested with a test of Monte Carlo. The results showed that the density of the macrofauna was higher in the period of maximum pluvial precipitation compared to the minimum (1129 vs 598 subjects, respectively). The agroforestal arrangements influences the presence or absence of some taxonomic groups of soil macrofauna (P<0.05) as Homoptera and Raphidioptera, besides than UV and AB can favor the macrofauna of the stress due to the drought.
Key words: Soil fauna, agroforestry arrangement, period, density, richness, diversity.
Introducción
La macrofauna en el suelo genera beneficios para la sostenibilidad de los agroecosistemas, entre ellos, incremento en la dinámica de la materia orgánica y cambios de algunas de las propiedades físicas del suelo (Lavelle et al., 2006). La actividad, riqueza y diversidad de estos organismos pueden ser afectadas por variaciones en el tipo de cobertura vegetal, la calidad del material vegetal y las variaciones estacionales (Velásquez, 2004).
Los cambios en la cobertura vegetal en la región amazónica han generado igualmente cambios en la macrofauna del suelo (Barros et al., 2002; Schon et al., 2012). Por otra parte, los arreglos agroforestales protegen la macrofauna del suelo alterada por las variaciones de temperatura y estrés por sequía (Lavelle et al., 2003); por tanto, la optimización del manejo de estos arreglos contribuye a la estabilidad de las poblaciones de macrofauna (Barros et al., 2003) y en consecuencia a la calidad del suelo (Velásquez et al., 2007).
Según Brown et al., 2004) existe mayor riqueza y abundancia de comunidades de invertebrados en suelos con arreglos agroforestales comparado con suelos en pasturas, ésta diferencia se debe al aporte de hojarasca de las especies arbóreas, lo que favorece el microclima y es fuente de energía y nutrientes para la microfauna (Lavelle et al., 2003; Velásquez, 2004; Huerta y Wal, 2012).
En la zona de estudio (departamento del Caquetá, Colombia) se presenta un régimen de lluvias monomodal con un período corto de mínima precipitación entre diciembre y febrero y un período lluvioso en el resto del año, alcanzando sus mayores niveles entre abril y agosto (Olaya et al., 2005). Las épocas de máxima y mínima precipitación afectan la densidad de la macrofauna edáfica, que tiene una alta correlación con la humedad en el suelo y disminuye drásticamente en la época de menor precipitación (Gamboa et al., 2011). Debido a la escasa información sobre los factores que influyen y determinan la distribución de la fauna en el suelo y la hojarasca en los arreglos agroforestales en la Amazonía colombiana y a la necesidad de identificar los factores que influyen en la presencia o ausencia de los grupos tróficos en los arreglos agroforestales, el objetivo de este estudio fue estimar la densidad y riqueza de poblaciones de macrofauna edáfica asociada con arreglos agroforestales existentes en el Centro de Investigaciones Macagual Cesar Augusto Estrada González.
Materiales y métodos
Sitio y áreas de estudio
El Centro de Investigaciones Macagual Cesar Augusto Estrada González está localizado a 1° 37’ N y 75° 36’ O, 300 m.s.n.m., con un clima AF según la clasificación de Köppen, precipitación promedio anual de 3793 mm, brillo solar de 1707 horas/año, temperatura promedio de 25.5 °C y humedad relativa de 84.25%, a 22 km del municipio de Florencia, sur del departamento del Caquetá (Colombia). El Centro tiene una extensión de 380 ha dedicadas a la explotación ganadera con algunos arreglos agroforestales: AB = abarco (Cariniana pyriformis); CH = caucho (Hevea brasiliensis); CP = caucho-parica (Schizolobium amazonicum Huber); UV = uvito (Genipa Americana L.) (Tabla 1), donde se adelantan proyectos de investigación en aspectos relacionados con el manejo eficiente de estos sistemas de producción.
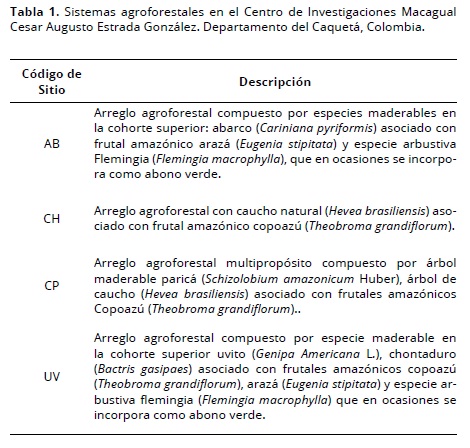
Toma e identificación de la macrofauna
Para la determinación de la macrofauna en el suelo se utilizó la metodología propuesta por Tropical Soil Biology and Fertility (Anderson y Ingram, 1993). En cada arreglo agroforestal, en mayo (época lluviosa) y noviembre (época de mínima precipitación) de 2011 se tomaron al azar cuatro monolitos (bloques de 25 x 25 cm a una profundidad de 30 cm) con el fin de comparar el efecto de la estacionalidad de la precipitación en la composición de la macrofauna. La macrofauna presente en cada monolito se extrajo de forma manual, los invertebrados recolectados fueron conservados en alcohol a 70% y separados de acuerdo con su morfología para su identificación en estereoscopio hasta el nivel de orden con la ayuda de manuales especializados (Triplehorn y Johnson, 2005).
Diseño y análisis de datos
Para los estudios se utilizaron cuatro tratamientos (arreglos agroforestales) que fueron evaluados en épocas de máxima y mínima precipitación, con cuatro repeticiones en un modelo bifactorial con diseño completamente al azar en parcelas divididas. La parcela principal fue el arreglo y la subparcela cada época. El modelo aplicado fue el siguiente:
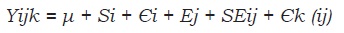
dónde: Yijk = una observación; µ = media; Si = efecto del i-ésimo arreglo; Єi = error debido al arreglo; Ej = efecto de la j-ésima época; SEij = interacción arreglo x época; Єk(ij) = error debido a la época.
La comparación de los arreglos agroforestales se hizo con base en la densidad de individuos por órdenes, riqueza de órdenes (Σpi), equidad o uniformidad mediante el Indice de Pielou (e) (Pielou, 1969), que mide la proporción de la diversidad observada en relación con la máxima diversidad esperada. Su valor varía entre 0 y 1, de forma que 1 corresponde a situaciones donde todas las especies son igualmente abundantes, definido como:
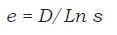
donde, D = Índice de Shannon (Shannon y Weaver, 1949) y S = número de especies o grupos taxonómicos (Magurran, 1988).
Los datos de densidad de edafofauna y riqueza de taxones encontrados en diferentes arreglos agroforestales y épocas de precipitación fueron sometidos a un análisis de varianza y comparación de medias mediante la prueba LSD de Fisher (P < 0.05). Los taxones Blattodea, Dermaptera, Homoptera, Orthoptera y Raphidioptera, que presentaron baja densidad en los arreglos agroforestales en ambas épocas, fueron agrupados como categoría ‘otros’ y tenidos en cuenta para el cálculo de la riqueza taxonómica total (RT). Además, se realizó un Análisis de Componentes Principales (ACP) para determinar la similitud entre arreglos agroforestales por los taxones presentes y explorar las relaciones entre los órdenes. Los datos se transformaron por log10(x +1), para reducir el rango de variación de la densidad, que fue muy alta en insectos sociales en comparación con los demás. Para evaluar el efecto de los arreglos agroforestales se hizo una prueba de Monte Carlo (Lavelle et al.., 2014). El análisis de coordenadas principales se realizó utilizando el paquete R versión 3.1.1 (R Development Core Team, 2014) utilizando la librería Ade4 (Dray y Dufour, 2007; Chessel et al., 2004).
Resultados y discusión
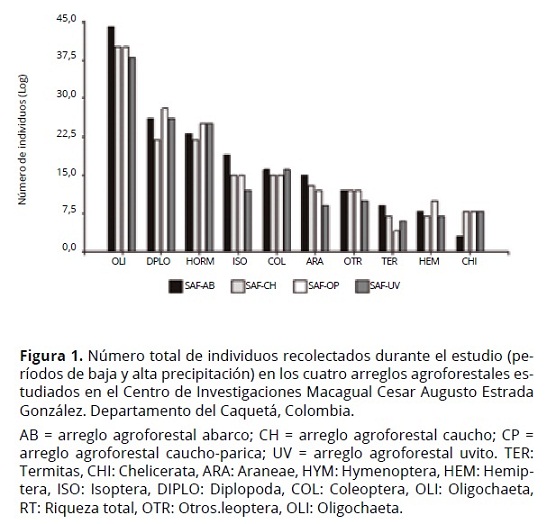
En total, en ambas épocas de evaluación se tomaron 32 muestras y fueron recolectados 1727 individuos. El número de individuos recolectados fue más alto en la época de máxima precipitación (1129 individuos) en comparación con los recolectados en la época de mínima precipitación (598 individuos). Algunos arreglos agroforestales presentaron densidades más altas en ciertos grupos taxonómicos, entre ellos: Oligochaeta (OLI), Diplópodo (DIP) y Isoptera (ISO) en Abarco (AB); Oligochaeta (OLI) y Termitas (TER) en Caucho (CH); Araneae (Arachnida) en Caucho-Parica (CP) y Hormigas (HOR) y Coleoptera (COL) en Uvito (UV) (Figura 1).
Se observó interacción entre la época y el tipo de arreglo agroforestal para la densidad (P < 0.004) y la riqueza (P < 0.0043). En la época de mínima precipitación se presentó un contraste entre la riqueza y la densidad, siendo la primera mayor en los arreglos AB y CP (P < 0.05) (Tabla 2).
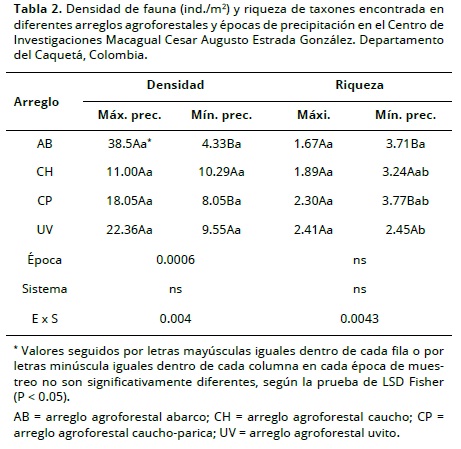
El Índice de Diversidad de Shannon fue más alto en la época de menor precipitación para todos los arreglos agroforestales (P < 0.05). El índice de Equitatividad de Pielou no mostró diferencias (P > 0.05), a pesar de que varió entre los diferentes arreglos y entre época (Tabla 3), lo que posiblemente fue debido a la similitud que presentaron los arreglos en relación con la estructura, una situación que mantiene estable las condiciones climáticas de radiación y temperatura que favorecen la distribución homogénea de las comunidades de fauna.
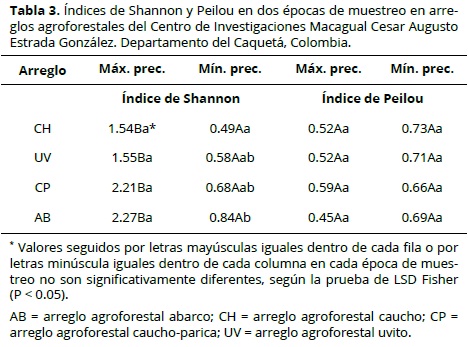
El análisis de los grupos taxonómicos presentes en cada uno de los arreglos agroforestales por cada periodo de precipitación, mostró variaciones en el número de taxones e individuos. Los órdenes con mayor presencia de individuos, independientemente de la época y del arreglo, fueron Oligochaeta y hormigas con 990 y 189 individuos en total, respectivamente.
Grupos taxonómicos como Homoptera en UV y Raphidioptera en CH se encontraron sólo en un arreglo agroforestal, lo cual es una respuesta a la configuración del arreglo relacionada con las condiciones de temperatura y disponibilidad de materia orgánica (Figura 2). En la evaluación presencia/ausencia se encontró que Hemiptera (HEM) fue el único orden que estaba presente de manera exclusiva durante el periodo de máxima precipitación, lo que sugiere la sensibilidad de este grupo ante los cambios de temperatura y humedad. Por su condición de depredadores, algunos grupos taxonómicos, como los Chilopodos, están presentes en esta época debido a la disponibilidad de alimento (Zerbino et al., 2008; Cabrera et al., 2011).
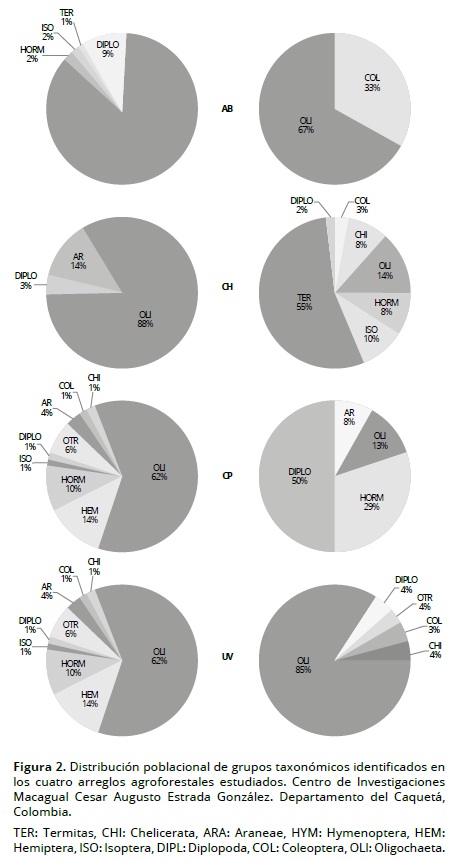
En la Figura 3 se observa el arreglo de los taxones en el plano factorial F1/F2 del ACP de las comunidades de macroinvertebrados. El F1, que explica 26.9% de la varianza, opone sistemas (especialmente CP y UV) con mayor densidad y diversidad de macrofauna, especialmente de Diplopoda, hormigas e invertebrados de la hojarasca (muchos de ellos agrupados en la categoría ‘otros’) a sistemas más pobres (CH). El eje 2 separa el sistema AB de los demás por las densidades mayores de lombrices (OLI), Coleóptera (COL) y Diplopoda, grupos indicadores de medios abiertos. Los Diplopodos (DIPL) son más comunes en sistemas forestales, aunque algunos (Polydesmidae) asociados con cultivos, pueden actuar como plagas. La prueba de Monte Carlo de permutación de las coordenadas de los puntos, indica que la separación entre los sistemas de uso es altamente significativa (P < 0.001) y explica 36.9% de la varianza.

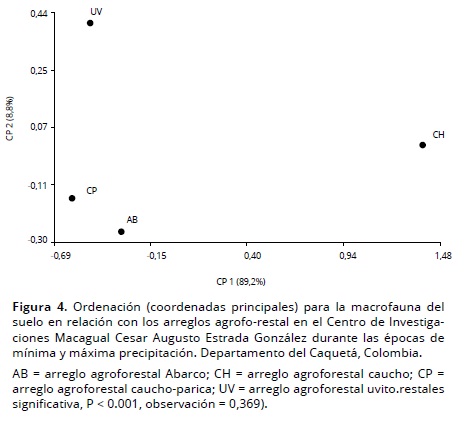
De acuerdo con el análisis de coordenadas principales, los arreglos agroforestales CP y AB comparten taxones, un caso contrario presentan CH y UV (Figura 4). Se encontraron taxones exclusivos para algunos arreglos, por ejemplo: Homoptera y Dermaptera en UV y Raphidioptera en CH. La distribución de los grupos taxonómicos presentes en cada arreglo agroforestal se incluyen en el tabla 4.
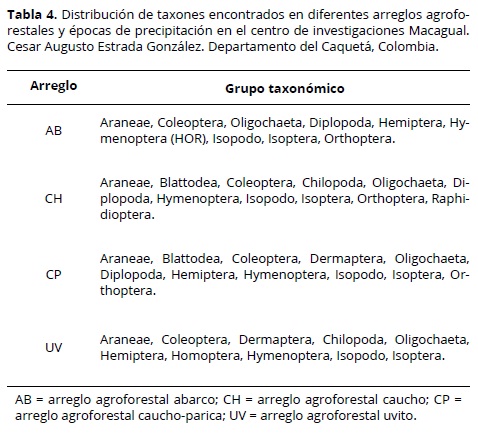
En general, los arreglos agroforestales se diferenciaron de acuerdo con los grupos taxonómicos de macrofauna presente en el suelo, por ejemplo, taxones que desde el punto de vista funcional se agrupan como depredador, detritivoro, herbívoro e ‘ingeniero’ del suelo (Zerbino et al., 2008; Cabrera et al., 2011). Dentro de los taxones dominantes en el grupo CH se encuentra Isóptera (TER), que juega un papel benéfico en la promoción de procesos ecológicos esenciales en los agroecosistemas como son los flujos de carbono y nitrógeno y descomposición de la materia vegetal, que aumentan la fertilidad del suelo (Lavelle et al., 1994). El taxon Diplopoda, al cual pertenecen los que son básicamente organismos detritívoros Zerbino et al., 2008; Cabrera et al., 2011) característicos de los agroecosistemas donde existe un alto aporte de biomasa que favorece el mantenimiento de humedad (Hadjicharalampous et al., 2002), condición propicia para su desarrollo.
Debido a la estructura de arreglos agroforestales, entre ellos CP que contribuye a la regulación de temperatura y del agua, posibilitando la presencia de otros individuos o presas (Bell et al., 2001), existió una mayor presencia del orden Aráneae (Arachnida). Estos resultados evidencian que los arreglos agroforestales AB y CP presentan una menor densidad de macrofauna en la época de mínima precipitación, lo que concuerda con los resultados de Jiménez et al. (2003) quienes concluyen que los valores mensuales de densidad estuvieron estrechamente ligados con la humedad en el suelo, mientras que la presencia de la época seca disminuyó drásticamente la densidad poblacional. Silveira et al. (2013) consideran que la apertura del dosel agroforestal afecta la temperatura y la humedad, así mismo, la densidad y riqueza de algunos taxones como Hymenoptera. Lo anterior se relaciona con la interacción entre las épocas y los arreglos UV y CH obtenida en el presente estudio. Velásquez (2004) y Pauli et al. (2011) encontraron la existencia de una relación negativa entre la densidad y la riqueza de macrofauna en el suelo, probablemente debido a la presencia de un grupo dominante, una situación que se presentó en los grupos AB y CP en la época de máxima precipitación.
El Índice de Shannon mostró que el efecto de la época en la macrofauna del suelo es evidente en todos los arreglos, con valores más altos en la época de máxima precipitación, coincidiendo con lo reportado por Manhães et al. (2013). A pesar de la drástica reducción de este índice en la época de mínima precipitación, el arreglo AB conserva una mayor diversidad, lo que puede estar relacionado con su nivel de complejidad (Barros et al., 2002).
Conclusiones
- Los resultados en este estudio muestran un efecto significativo de la precipitación sobre la diversidad y densidad de los diferentes taxones de la macrofauna edáfica en la Amazonía colombiana, presentando valores más bajos en la época de mínima precipitación.
- Debido a la complejidad estructural del arreglo agroforestal AB (sistemas agroforestal Abarco), este conserva una mayor diversidad, aun durante el periodo de sequía.
- Se presentaron grupos taxonómicos exclusivos en algunos arreglos agroforestales, como Raphidioptera en CH y Homoptera para UV, condición que se relaciona con la configuración agroforestal que favorece la conservación de la humedad por el aporte de biomasa proveniente de las especies que componen el dosel y su disposición espacial, lo que a su vez se relaciona con una mayor densidad de individuos/m2 en los mismos arreglos agroforestales, independientemente de la época de muestreo.
Referencias
Anderson, J. M. e Ingram, J. S. I. 1993. Tropical soil biology and fertility: a handbook of methods. 2nd ed. Wallingford. CABI. p. 240. [ Links ]
Barros, E.; Neves, A.; Blanchart, E.; Fernandes, E.; Wandelli, E. y Lavelle P. 2003. Development of the soil macrofauna community under silvopastoral and agrisilvicultural systems in Amazonia. Pedobiologia 47(3):273 - 280. [ Links ]
Barros, E.; Pashani B.; Constantino R. y Lavelle, P. 2002. Effects of land-use system on the soil macrofauna in western Brazilian Amazonia. Biol. Fertil. Soils 35(5):338 - 347. [ Links ]
Bell, J. R.; Wheater, C. P.; y Cullen, W. R. 2001. The implications of grassland and heathland management for the conservation of spider communities: a review. J. Zool. 255:377 - 387. [ Links ]
Brown, G. G.; Moreno, A. G.; Barois, I.; Fragoso, V.; Rojas, P.; y Hernández, B. 2004. Soil macrofauna in SE Mexican pastures and the effect of conversion from native to introduced pastures. Agric. Ecosys. Environ. 103(2):313 - 327. [ Links ]
Cabrera, G.; Robaina, N. y Ponce de León, L. 2011. Composición funcional de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba. Pastos y Forrajes 34(3):331 - 346. [ Links ]
Chessel, D.; Dufour, A.B., y Thioulouse, J., 2004. The ade4 package-I-One-table methods. R News 4:5 - 10. [ Links ]
Dray, S. y Dufour, A. B., 2007. The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22:1 - 20. [ Links ]
Gamboa, J. A.; Suárez, J. C.; y Chaves, J. A. 2011. Macroinvertebrados edáficos asociados a rastrojos en paisaje de lomerío de la Amazonia colombiana. Valoración de rastrojos para la formación de sistemas silvopastoriles en la Amazonia colombiana. Universidad de la Amazonia p.116 - 128. [ Links ]
Hadjicharalampous, E.; Kalburtji, K. L.; y Mamolos, A. P. 2002. Soil Arthropods (Coleoptera, Isopoda) in organic and conventional agroecosystems. Environ. Manage. 29(5):683 - 690. [ Links ]
Huerta, E. y Wal, V. 2012. Soil macroinvertebrates abundance and diversity in home gardens in Tabasco, Mexico, vary with soil texture, organic matter and vegetation cover. Eur. J. Soil Biol. 50:68 - 75. [ Links ]
Husson, F.; Josse, J.; Le, S.; y Mazet, J. 2012. Facto-MineR: Multivariate exploratory data analysis and data mining with R. R package version 1.18. http://CRAN.R-project.org/package=FactoMineR. [ Links ]
Jiménez, J. J; Moreno, A. G; Decans, T; Lavelle, P; Fisher, M. J. y Thomas, R. J. 2003. Las comunidades de lombrices en las sabanas nativas y en los pastizales introducidos de los Llanos Orientales de Colombia. En: Jiménez, J. J. y R. J. Thomas (eds.). El arado natural: Las comunidades de macroinvertebrados del suelo en las sabanas neotropicales de Colombia. Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia. p. 57 - 75. [ Links ]
Kadamannaya, B. S y Sridhar, K. R. 2009. Leaf litter ingestion and assimilation by two endemic pill millipedes (Arthrosphaera). Biol. Fertil. Soils 45:761 - 768. [ Links ]
Lavelle, P.; Rodríguez, N.; Arguello, O. et al. 2014. Soil ecosystem services and land use in the rapidly changing Orinoco river basin of Colombia. Agriculture, Ecosys. Environ. 185:106 - 117. [ Links ]
Lavelle, P.; Decaëns, T.; Aubert, M. 2006. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 42(1):3 - 15. [ Links ]
Lavelle, P.; Senapati, B.; y Barros, E. 2003. Soil macrofauna. En: Schroth, G. y Sinclair. F. L. (eds.). Trees, crops and soil fertility. CABI Publishing, Wallingford. p. 303 - 324. [ Links ]
Lavelle, P.; Dangerfield, M.; Fragoso, C. et al. 1994. The relationship between soil macrofauna and tropical soil fertility. En: Woomer, P. y Swift, M. (eds.). The Biological management of tropical soil fertility. John Wiley and Sons, Chichester. p. 137 - 170. [ Links ]
Manhães, C. M. C.; Gama-Rodrigues, E. F.; Moço, M. K. S.; y Gama-Rodrigues, A. C. 2013. Meso-and macrofauna in the soil and litter of leguminous trees in a degraded pasture in Brazil. Agroforestry systems, 87(5), 993-1004. [ Links ]
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press, New Jersey. p. 179. [ Links ]
Olaya, E. J.; Gaitan, A. E.; y Millan E. E. 2005. Revisión y ajuste a la propuesta de plan de ordenación y manejo ambiental de las microcuencas La Batea y San Joaquín, municipio de Florencia. Tesis de grado. Facultad de Ingenieria. Programa de Ingenieria Agroecologica. Universidad de la Amazonia. 136 p. [ Links ]
Pauli, N.; Barrios E.; Conacher, A. J. y Oberthür, T. 2011. Soil macrofauna in agricultural landscapes dominated by the Quesungual slash-and-mulch agroforestry system, western Honduras. Appl. Soil Ecol. 47(2)119 - 132. [ Links ]
Pielou, E. C. 1969. An Introduction to mathematical ecology. Wiley-Interscience, Nueva York. [ Links ]
R Development Core Team. 2014. R: A language and environment for statistical computing. R. Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/ [ Links ]
Schon, N. L.; Mackay, A. D. y Minor, M. A. 2012. Vulnerability of soil invertebrate communities to the influences of livestock in three grasslands. Appl. Soil Ecol. 53:98 - 107. [ Links ]
Shannon, C. E.; Weaver, W. 1949. The mathematical theory of communication. Urbaba: University of Illinois Press. p. 78. [ Links ]
Silveira, J. M.; Barlow, J.; Andrade, R. B. et al. 2013. The responses of leaf litter ant communities to wildfires in the Brazilian Amazon: a multi-region assessment. Biodiv. Conserv. 22:513 - 529. [ Links ]
Triplehorn, C. y Johnson, F. 2005. Borror and delong's: introduction to the: study of insects. 7a ed. Ed. Thomson. EE.UU. p. 864. [ Links ]
Velásquez, E. 2004. Bioindicadores de calidad de suelo basado en las poblaciones de macrofauna y su relación con características funcionales del suelo. Ph.D. tesis. Universidad nacional de Colombia sede Palmira. 189 p. [ Links ]
Velásquez, E.; Lavelle, P.; y Andrade, M. 2007. GIQS: a multifunctional indicator of soil quality. Soil Biol. Biochem. 39:3066 - 3080. [ Links ]
Zerbino, S.; Altieri, N.; Morón, A.; y Rodríguez, C. 2008. Evaluación de la macrofauna del suelo en sistemas de producción en siembra directa y con pastoreo. Agrociencia 12:(1)44 - 55. [ Links ]
