










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Agronómica
versão impressa ISSN 0120-2812
Acta Agron. vol.65 no.3 Palmira jul./set. 2016
https://doi.org/10.15446/acag.v65n3.49555
http://dx.doi.org/10.15446/acag.v65n3.49555
Efecto del nitrógeno en los parámetros fotosintéticos y de producción del cultivo de la gerbera (Gerbera jamesonii H. Bolus ex Hook. f.)
Effect of nitrogen on photosynthetic parameters and yield of gerbera crop (Gerbera jamesonii Bolus ex Hook. f.)
Maykoll Corrales–González1*, Fermín Rada1 y Ramón Jaimez2
1Universidad de Los Andes. Instituto de Ciencias Ecológicas y Ambientales. Mérida, Venezuela. 2Universidad de Los Andes. Facultad de Ciencias Forestales y Ambientales. Laboratorio de Ecofisiología de cultivos. Mérida, Venezuela. *Autor para correspondencia: maycogon@alumni.uv.es
Rec.:08.03.215 Acep.:17.07.2015
Resumen
Actualmente, existe un interés por comprender los aspectos fisiológicos involucrados en la obtención de altas producciones en diferentes cultivos y su relación con la disponibilidad de nutrientes en el sustrato. Este conocimiento ayudará en las recomendaciones de dosis acertadas de nutrientes, evitando problemas asociados a desequilibrios tanto en el suelo como en el medio ambiente. Este trabajo evaluó algunas respuestas fisiológicas de Gerbera jamesonii frente al efecto de cuatro tratamientos de fertilización nitrogenada (75, 56, 94 y 112.5 mgN planta-1 día-1). Al mismo tiempo se buscó relacionar estas posibles respuestas con características de crecimiento. Las plantas se distribuyeron en un diseño completamente al azar con 3 repeticiones, a las cuales se les midieron las variables de intercambio de gases (asimilación de CO2, transpiración, conductancia estomática), potenciales hídricos foliares y rendimiento cuántico del fotosistema II. Estas variables se relacionaron con el crecimiento vegetativo (área foliar total) y las variables de calidad y producción de flores (longitud y diámetro de tallos florales, diámetro del capítulo floral y el número de flores mensuales). Los resultados muestran que el aumento en la dosis de N a nivel del sustrato trajo como consecuencia un aumento en la asimilación de CO2 (de 9.2 a 12.1 µmol m-2 s-1). Sin embargo, es posible reducir la dosis de fertilización de N en un 25% (56 mgN planta-1 día-1) sin afectar la producción mensual de flores ni la calidad de estas (no hubo diferencias significativas entre los tratamientos (P>0.05)).
Palabras clave: Asimilación de CO2, fluorescencia de clorofila a, intercambio de gases, producción de flores.
Abstract
There is a current interest in understanding the physiological aspects involved in gaining high crop yields, and their relationship with biomass production and substrate nutrient availability. This knowledge will allow the recommendation of accurate nutrient doses, avoiding problems associated to imbalances in both the soil and the environment. This study evaluated some physiological responses of Gerbera jamesonii (cultivar Cirilo®) under the effect of four nitrogen fertilization treatments (75, 56, 94 and 112.5 mgN plant-1 day-1). We also sought to relate these possible responses with growth characteristics. Plants were arranged in a completely randomized design with 3 replications to which gas exchange (CO2 assimilation, transpiration, and stomatal conductance), leaf water potential and quantum yield of photosystem II were measured. These variables were related to vegetative growth (total leaf area) and quality and flower production variables (floral stem length and diameter, flower head diameter and number of monthly flowers). The results show that increasing substrate N doses resulted in an increase in CO2 assimilation for this cultivar (from 9.2 to 12.1 mmol m-2s-1). However, it is possible to reduce N fertilization by 25% (56 mg N plant–1 day–1) without affecting monthly flower production and quality variables (no significant differences were found between treatments (P>0.05)).
Keywords: Chlorophyll a fluorescence, CO2 assimilation, flower production, gas exchange.
Introducción
El nitrógeno es el principal limitante de la productividad en las plantas, por ello en la producción agrícola, según el estado nutricional del suelo, se utilizan fertilizantes nitrogenados inorgánicos para compensar algunas deficiencias. Sin embargo, la aplicación de fertilizantes nitrogenados es en general ineficiente debido a que sólo una tercera parte del fertilizante aplicado es absorbido de forma eficiente por el cultivo (Shrawat et al., 2008). Esta eficiencia es expresada como: unidad de biomasa generada por unidad de nutriente aplicado. En el caso del nitrógeno, la eficiencia fisiológica va a depender de las características de la especie y de la disponibilidad de dicho nutriente (Shrawat et al., 2008). Dado que la maquinaria fotosintética requiere más de la mitad del nitrógeno foliar, la fotosíntesis será afectada por la disponibilidad de nitrógeno, independientemente de que la variación de nitrógeno foliar sea causada por la disponibilidad de este nutriente en el suelo, por la edad de la hoja o por incrementos en la radiación.
Los procesos primarios que determinan el crecimiento vegetal son los que involucran el intercambio de gases entre las hojas y el aire circundante, fotosíntesis, respiración y transpiración (Taiz y Zeiger, 2006). Estos procesos también influyen en la distribución de biomasa entre órganos y en el rendimiento del fruto (Peil y Galvez, 2012). El intercambio de gases se lleva a cabo a través de los estomas y responde a diversos factores ambientales, entre los que destacan la luz, concentración de CO2 y disponibilidad de nitrógeno (Schulze et al., 2005). Los asimilados, producidos por la fotosíntesis en los órganos fuente pueden ser almacenados o transportados, vía floema, a los diferentes órganos sumideros. Se considera que el suministro de nitrógeno a la planta traería consigo un aumento de su tasa fotosintética, incrementando el crecimiento de los cultivos dado que toda la materia seca producida por la planta depende totalmente de este proceso (Peil y Galvez, 2012).
Por ejemplo, en trigo (Triticum aestivum L.), girasol (Helianthus annuus L.) y maíz (Zea mays L.) se demostró que al aumentar el suministro de N se aumentó la asimilación de CO2 (Lindquist, 2001; Cechin y de Fátima, 2004). Aunque hasta ahora no se utilice la tasa fotosintética como indicador directo para incrementar el rendimiento de un cultivo dentro de un programa de mejoramiento genético, algunos cultivares de trigo seleccionados para alto rendimiento de grano presentan tasas altas de fotosíntesis como característica asociada. Lo mismo ocurre con genotipos de soya (Glycine max L.), frijol (Pisum sativum L.) y rosas, donde los mayores rendimientos mostraron también altas tasas de asimilación de CO2 (Gutiérrez et al., 2005; Plaut et al., 2006).
Específicamente en Gerbera jamesonii, este patrón descrito anteriormente: aumento de la producción del cultivo debido a un aumento en la fertilización nitrogenada parece no estar muy claro, ya que se ha encontrado que aunque se aumente la cantidad de N en un 30 o 40% esto no repercute en un aumento en la producción mensual de flores, sin importar la variedad o cultivar de G. jamesonii estudiado (Ortega et al. 2006; Şirin 2011). Zheng et al. (2005) concluyen que es posible reducir la fertilización nitrogenada hasta en un 50% en el cultivo de las gerberas, sin afectar el desarrollo de la planta ni el rendimiento floral de estas.
Actualmente existe un interés por comprender los aspectos fisiológicos involucrados en la obtención de altas producciones en diferentes cultivos y su relación con la disponibilidad de nutrientes en el sustrato. Este conocimiento ayudará en las recomendaciones de cantidades acertadas de nutrientes, evitando problemas asociados a desequilibrios tanto en el suelo como en el medio ambiente. Dado que existe poca información fisiológica que permita explicar las respuestas de G. jamesonii a cambios en el suministro de nitrógeno, lo cual es importante para comprender su relación con la producción de flores. El objetivo de este trabajo fue evaluar el efecto de diferentes tratamientos de fertilización nitrogenada, sobre las tasa de intercambios de gases y actividad fotoquímica, y su relación con la calidad y productividad del cultivo de G. jamesonii.
Materiales y métodos
El estudio se llevó a cabo en un invernadero ubicado en la Estación Experimental Santa Rosa, Instituto de Investigaciones Agropecuarias (IIAP), Universidad de Los Andes (ULA), Mérida, Venezuela, localizado geográficamente a 8°35'30'' N y 71°08'30'' O, a una elevación de 1920 msnm. El invernadero es tipo túnel, con un área de 162 m2 y altura máxima de cumbrera de 6 m. El microclima dentro del invernadero fue monitoreado constantemente cada 30 minutos con sensores de temperatura del aire y densidad de flujo fotónico fotosintético conectados a una microestación (Marca HOBO, Onset Inc., USA). Se registró una temperatura máxima promedio de 24.5°C a las 12:00 h, y una mínima de 12.5°C a las 6:00 h. Las máximas radiaciones promedio se registraron entre las 11:00 y las 13:00 h estando siempre alrededor de los 500 µmol m-2 s-1.
El experimento se hizo en plantas de 3 años de edad del cultivar Cirilo® (Florist) de la especie G. jamesonii, sembradas en bolsas de 30 cm de alto y en un sustrato compuesto en partes iguales por arena y compost de caballerías. Se colocaron 10 plantas m-2 en un diseño completamente al azar.
Tratamientos. Se evaluaron 4 tratamientos de fertilización nitrogenada (Tabla 1), teniendo como control (T0) la cantidad de nitrógeno usado para el cultivo de gerberas en la estación experimental IIAP – ULA (75 mgN planta-1 día-1; Jaimez et al., 2013), un tratamiento con una concentración 25% por debajo del control (T1), otro 25% por encima (T2) y uno último 50% por encima del control (T3). Las dosis se aplicaron 3 veces por semana, de igual modo cada tratamiento recibió la misma formulación de los macro y micro nutrientes: K2O= 48%; Ca= 0.05%; Mg=0.10%; S= 0.05%; Fe= 0.10%; Zn= 0.05%; Cu= 0.05%; B=0.02%; Mo= 0.0005%; Mn= 0.05% (obtenidos de 1.12 g día-1 del producto comercial soluble Grow More®) y la misma cantidad de agua (325 ml día-1, mediante sistema de riego por goteo).
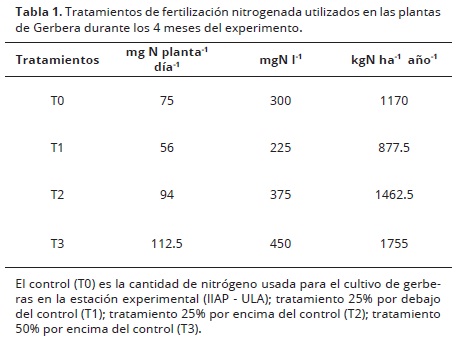
Para tener certeza de que el estado hídrico o lumínico no estuviera determinando diferencias entre los tratamientos, mensualmente se midió el potencial hídrico foliar (Ψ) y la fluorescencia de clorofila a. Las mediciones del Ψ se llevaron a cabo a las 11:00 h simultáneamente con las mediciones de intercambio de gases, en una hoja de tres individuos distintos por tratamiento, utilizando una cámara de presión de Schölander (MS modelo 600, Cornwallis, Oregon, USA). La eficiencia fotosintética (Fv/Fm) fue medida con un fluorímetro portátil modelo PAM 2100 (Heinz Wals GmbH, Alemania), en una hoja por planta, de tres individuos diferentes, para cada uno de los tratamientos. Dichas mediciones se hicieron en hojas previamente cubiertas por dos horas, con el fin de garantizar que todos los centros de reacción del fotosistema II estuvieran abiertos.
Medición de intercambio de gases. Para las mediciones correspondientes al intercambio de gases (asimilación de CO2 (A), transpiración (E) y conductancia estomática (Gs)) se utilizó un analizador infrarrojo de gases portátil (ADC LC–4). Las mediciones se hicieron en 3 individuos, en hojas completamente expandidas (una hoja por cada planta). Estas variables se midieron mensualmente, a partir de los 15 días después del inicio de la primera fertilización, a la hora de máxima radiación (entre las 10:00 y las 12:00 h).
Determinación del área foliar total. El crecimiento vegetativo se determinó midiendo el área foliar total (AFT), la cual se obtuvo de la sumatoria del área foliar (AF) de cada planta de cada tratamiento. Dicha AF tuvo que ser estimada a partir de un ajuste matemático lineal. Esta estimación nos permite obtener las AFT sin eliminar hojas de las plantas (Gutierrez y Lavin, 2000), ya que para una óptima producción de flores las plantas de gerbera deben mantener entre 16 y 18 hojas, con lo cual se reducen tanto problemas fitosanitarios, como deformaciones en los pedúnculos e inflorescencias (Jaimez et al., 2013). Fue necesario usar un método no destructivo, realizando una relación entre los valores del AF (la cual fue medida con un medidor de área foliar LI–COR 3100) y el largo (L) y ancho máximo (Am) de 39 hojas de distintos tamaños. El resultado de ajustar un modelo de regresión lineal múltiple para describir la relación entre AF y dos variables independientes (L y Am), se muestra en la ecuación 1.
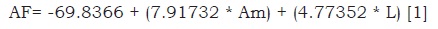
Donde: AF: es el área foliar, Am: es el ancho máximo de la hoja y L: es el Largo de la hoja; R2= 0.85, (P<0.001)
Calidad y producción de flores. El efecto de la dosis de N en la calidad de la flor se determinó midiendo la longitud del tallo floral, el diámetro del tallo floral y el diámetro del capítulo floral. Para la medida del diámetro del capítulo se tomó la distancia entre los extremos de las lígulas; la medida del diámetro del tallo florar se tomó en su parte media, y la longitud de este se registró midiendo desde la base hasta la unión con el receptáculo (Ortega et al., 2006). Para la evaluación de la producción de flores se contaron el número promedio de flores mensuales por planta. Las flores listas para cosechar eran cortadas cuando la inflorescencia presentaba dos o tres filas de flores masculinas abiertas.
Análisis estadístico. Los datos obtenidos de las mediciones anteriores (estado hídrico, fotoquímica, intercambio de gases, área foliar total y calidad y producción de flores) presentaron una distribución no normal, por lo cual se usó estadística no paramétrica. Se realizó una prueba de Kruskal–Wallis para hacer comparaciones entre los distintos tratamientos para cada una de las variables medidas. También se hicieron Análisis de correlación de Spearman y análisis de regresiones para medir la relación y la asociación entre variables. Para estos análisis se utilizó el programa Statistix 8® con un nivel de significancia del 95% (P<0.05).
Resultados y Discusión
Los Ψ estuvieron alrededor de –0.5 MPa durante los 4 meses del experimento lo que demuestra que las plantas estuvieron bajo condiciones hídricas favorables durante todo el período de estudio. No hubo diferencias significativas entre los tratamientos (P>0.05).
Por otra parte, todos los tratamientos mostraron una alta eficiencia fotosintética (Fv/Fm) ya que en promedio los valores siempre estuvieron por encima o igual a 0.8. Típicamente el índice Fv/Fm se halla alrededor de 0.8 en plantas no estresadas, y para las gerberas se han reportado valores por encima de 0.81 (Jaimez et al., 2013). Cualquier proceso estresante como: exceso de luz, altas o bajas temperaturas, déficit o problemas ligados al metabolismo del de nitrógeno, se pueden reflejar en una disminución en este índice. No se encontraron diferencias significativas para esta variable entre los tratamientos (P>0.05), lo que permite afirmar que en ningún tratamiento las plantas presentaron daño en el aparato fotosintético.
Intercambio de gases. La tabla 2, resume los valores promedios mensuales de las mediciones de intercambio de gases. La tasa de E mostró un incremento significativo en el T3 con respecto al tratamiento control, encontrándose diferencias significativas entre los tratamientos (P<0.05). Sin embargo, la eficiencia en el uso del agua (A/E) no es afectada debido a que la asimilación de CO2 también aumenta. Aunque la Gs no mostró ninguna diferencia estadísticamente significativa entre los tratamientos (P>0.05), los valores más altos para esta variable fueron encontrados en el T3 (tabla 2).
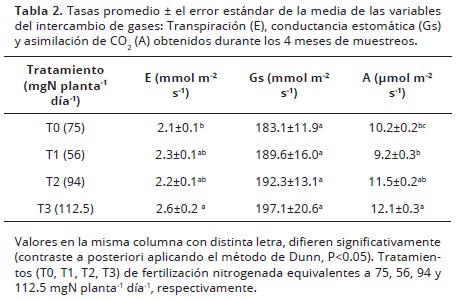
La tasa de A, mostró diferencias significativas entre los tratamientos (P<0.001; Tabla 2), el análisis de correlación de Spearman, mostró que hubo una tendencia hacia el aumento de A con el aumento de la dosis de N aplicada (r2= 0.86). Aumentar el suministro de nitrógeno implicó una mayor tasa de A (R2= 0.70), esta misma tendencia ha sido hallada en muchos cultivos Lindquist (2001), inclusive en plantaciones de girasol (Cechin y de Fátima, 2004). Los resultados de esta investigación muestran que una concentración de N por encima de los 112 mgN planta-1 día-1 resulta en mayores tasas de A. Estos resultados indican que una mayor dosis de N contribuye a mantener mayores tasas de asimilación de CO2 y a su vez determinan una mayor asignación de asimilados a reproducción en plantas que se encuentran en sus últimas etapas de producción (3 años).
Las tasas de A reportadas para G. jamesonii, son mayores que las encontradas en este trabajo, lo cual podría ser atribuido a las diferencias genotípicas existentes entre los distintos cultivares de esta especie o a prácticas agronómicas distintas (i.e. la cantidad de N suministrado al cultivo ó cantidad de radiación en el invernadero). Autores como Paradiso et al. (2003), y Zheng et al. (2005), reportan tasas que oscilan entre 12 – 15 µmol m-2 s-1 en condiciones de invernadero, a radiaciones alrededor de los 500 µmol m-2 s-1, pero en otros cultivares (Brittani®, Golden Serena® y Shogun®). Por otro lado, Jaimez et al. (2013), encontraron para el cultivar Cirilo® de un año de edad en el mismo invernadero, tasas de A promedio de 10 µmol m-2 s-1 a una dosis de 75 mgN planta-1 día-1, valor similar al encontrado en este trabajo usando similar dosis de N, lo que pareciera indicar que la tasa de asimilación no cambió con la edad de las plantas.
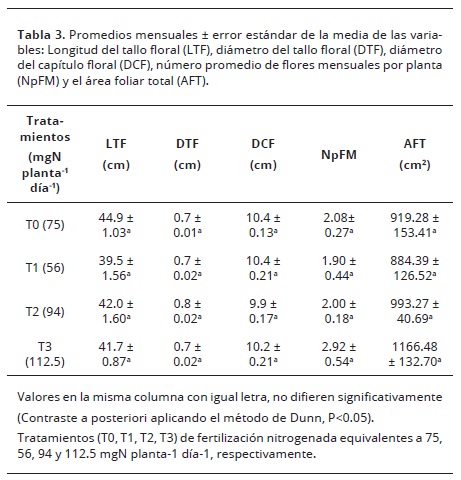
Área foliar, calidad y producción de flores. El suministro de nitrógeno en el cultivo de las gerberas no afectó el AFT (P>0.05) (Tabla 3), lo que coincidiría con los resultados hallados por Zheng et al. (2005). Sin embargo, otros autores han encontrado que incrementando el suministro de N aumenta el AFT. El análisis de correlación de Spearman en los resultados del presente estudio, demuestra que existe un incremento en el AFT a medida que aumenta la dosis de N (r2= 0.95).
Lawlor (2002), explica que el suministro de N es crucial para el crecimiento de la hoja debido a la función de las proteínas en el crecimiento de las paredes celulares y el citoesqueleto y por lo tanto en la expansión de las células. Además, concluye que la producción en los cultivos es totalmente dependiente de la provisión de N en cantidades adecuadas en los tiempos correctos del desarrollo de este, y que la consecuencia de los eventos metabólicos basados en proteínas, es que la energía de la luz es utilizada en la reducción de CO2 y NO3 y en la síntesis de asimilados que se utilizaran para el crecimiento vegetativo y reproductivo con un consecuente aumento en la producción. Afirmación que se ve reflejada en la alta relación encontrada en este trabajo entre A, el N suministrado y AFT (R2= 0.84), donde altos valores del área foliar total dependerían del aporte de N, lo que produciría un incremento del área para la asimilación de CO2 por planta con lo que se esperaría un aumento en la producción de biomasa (vegetativa o reproductiva) por planta, Aguilar et al. (2005) encontraron que un mayor crecimiento del dosel vegetal proporciona una mayor intercepción de luz, lo cual incrementa la fotosíntesis y producción de biomasa como resultado de un mayor aprovechamiento de los recursos hídricos y nutrimentales. Ahora bien, con base a esto y a nuestros resultados podemos sugerir que con una mayor área foliar por planta, la producción superior de biomasa en plantas del T3 fue posible, debido a una mayor tasa de fijación de CO2 por unidad de área foliar (Righetti et al., 2007).
La Tabla 3, expone los valores promedio de los indicadores de calidad y producción de las flores para todos los meses de muestreo (exceptuando el primer mes, ya que se tomó como mes de adaptación de las plantas a los tratamientos de fertilización nitrogenada).
El promedio en el número de flores por mes (NpFM), el cual se toma como estimador del rendimiento del cultivo alcanzó su valor más alto (2.9 flores mes-1) en el T3, lo que estaría correlacionado con los valores más altos de A (r2= 0.73). También se obtuvo una alta relación entre la interacción de las variables A, AFT y NpFM en un modelo de regresión lineal múltiple (R2= 0.84). Tanto investigaciones de campo como de laboratorio muestran que el crecimiento de las plantas es muy sensible a la fertilización nitrogenada. La deficiencia de N reduce la producción de hojas y el área foliar total (Toth et al., 2002), lo que resulta en una reducción de la superficie para la intercepción de luz utilizada en la fotosíntesis y una reducción en la asignación de asimilados a órganos sumideros como las flores (Peil y Galvez, 2012). Esto podría indicar que la tendencia de aumento en NpFM en el tratamiento T3 fue debido a menores limitaciones de recursos, y que una gran parte de los fotoasimilados se destinó a esta estructura reproductiva (inflorescencia). El suministro de N no afectó la longitud del tallo floral, el diámetro del tallo floral ni el diámetro del capítulo floral (P>0.05).
Autores como Jaimez et al. (2013), para este mismo cultivar y suministrando 75 mgN planta-1 día-1, reportaron una producción mensual de 2 flores por planta. Autores como Ortega et al. (2006), y Şirin (2011), concluyeron que las variables de calidad de las flores en gerberas no se incrementan cuando se aumenta la dosis de N suministrada. Sin embargo, el hecho de que en este trabajo, bajo cualquiera de las dosis estudiadas, las flores tengan una longitud del tallo floral igual o superior a 40 cm y diámetro del capítulo floral mayor de 8 cm, indicaría que las inflorescencias están clasificadas en el mercado como de calidad 1 o de tipo exportación. Esto permitiría recomendar nuestro tratamiento con menor cantidad de N (T1= 56 mgN planta-1 día-1) ya que con este se mantiene la producción mensual de flores y no se ve afectada su calidad.
Lograr una agricultura sustentable plantea un dilema complejo en donde la meta de alcanzar altos beneficios económicos de los cultivos contrasta con la necesidad de reducir el deterioro ambiental generado en el proceso productivo. Sin embargo, en lo que respecta a la utilización de fertilizantes, un manejo racional y acertado de los nutrientes agregados permite lograr óptimos niveles de productividad y al mismo tiempo se minimiza el impacto sobre el ambiente.
Conclusiones
- Bajo las condiciones climáticas de este estudio, los resultados de este experimento muestran que un aumento del suministro de nitrógeno al cultivar Cirilo® de G. jamesonii determina una mayor tasa de asimilación de CO2 y en consecuencia podría repercutir en un aumento de la biomasa de la planta.
- Este estudio sugiere la viabilidad en términos de una reducción en un 25% de la dosis de nitrógeno en la solución fertilizante utilizada en los Andes venezolanos, para el cultivar Cirilo® de G. jamesonii, sin afectar el desarrollo de la planta y manteniendo los mismos estándares de producción y calidad del cultivo.
Agradecimientos
Este trabajo fue apoyado técnicamente por el Instituto de Ciencias Ambientales y Ecológicas (ICAE) y el Laboratorio de Ecofisiología de cultivos del Instituto de Investigaciones Agropecuarias de la Universidad de los Andes (IIAP–ULA), Mérida, Venezuela.
Referencias
Aguilar. L. Escalante. J. Fucikovsky. L. Tijerina. L. Mark. E. (2005). Área foliar, tasa de asimilación neta, rendimiento y densidad de población en girasol. Terra Latinoamericana, 23(3), 303–310. [ Links ]
Cechin. I. de Fátima. T. (2004). Effect of nitrogen supply on growth and photosynthesis of sunflower plants grown in the greenhouse. Plant Sci, 166(5), 1379–1385. doi:10.1016/j.plantsci.2004.01.020. [ Links ]
Gutiérrez. A. Lavin. A. (2000). Mediciones lineales en la hoja para la estimación no destructiva del área foliar en vides Cv. Chardonnay. Agricultura Técnica, 60(1), 69–73. [ Links ]
Gutiérrez. M. Reynolds. M. Escalante. J. Larqué. A. (2005). Algunas consideraciones en la relación entre fotosíntesis y el rendimiento de grano en trigo. Ciencia ergo sum, 12(02), 149 – 154. [ Links ]
Jaimez. R. Araque. O. Espinoza. W. Azocar. C. (2013). Dinámica de producción de flores de cultivares de gerbera (Gerbera jamessoni H. Bolus): relación con tasas fotosintéticas. Rev Fac Agron Univ Zul, 30(4), 178–191. [ Links ]
Lawlor. D. (2002). Carbon and nitrogen assimilation in relation to yield: mechanisms are the key to understanding production systems. J Exp Bot, 53(370), 773–787. doi: 10.1093/jexbot/53.370.773. [ Links ]
Lindquist. J. (2001). Light–saturated CO2 assimilation rates of corn and velvetleaf in response to leaf nitrogen and development stage. Weed Sci, 49(6), 706–710. doi: 0043– [ Links ]1745(2001)049[0706:LSCARO]2.0.CO;2.
Ortega. L. Miranda. D. Sandoval. M. (2006). Densidad de huevos y ninfas de mosca blanca Trialeurodes vaporarium (west.) en Gerbera jamesonii H. Bolus, con diferentes regímenes de fertilización nitrogenada. Agrociencias, 40(3), 363–371. [ Links ]
Paradiso. R. De Pascale. S. Aprea. F. Barbieri. G. (2003). Effect of electical conductivity levels of nutrients solutions on growth, gas exchange of two gerbera cultivars in soilless system. Acta Hort (ISHS), 609, 165–171. http://www.actahort.org/books/609/609_22.htm. 09.06.2014. [ Links ]
Peil. R. Galvez. J. (2012). Reparto de materia seca como factor determinante de la producción de las hortalizas de fruto cultivadas en invernadero. R. bras. Agrociência, 11(1), 5–11. [ Links ]
Plaut. Z. Dayan. E. Grava. A. Presnov. E. Matan. E. Liu. F. (2006). Long Term Production of Rose Flowers in the Greenhouse under different Cooling Methods: II. Responses of the Flower Bud. Eur J Hortic Sci, 71(1), 7–14. [ Links ]
Righetti. T. Vasconcelos. C. Sandrock. D. Ortega. S. Moreno. Y. Meza. F. (2007). Assessments of CO2 Assimilation on a Per–leaf–area Basis are related to Total Leaf Area. J Am Soc Hortic Sci, 132(2), 230–238. [ Links ]
Schulze. E. Beck. E. Müller. K. (2005). Plant ecology. Springer – Verlag, Berlin, Alemania. 702 p. [ Links ]
Shrawat. A. Carroll. R. DePauw. M. Taylor. G. Good. A. (2008). Genetic engineering of improved nitrogen use efficiency in rice by the tissue–specific expression of alanine aminotransferase. Plant biotech j, 6(7), 722–732. doi: 10.1111/j.1467–7652.2008.00351.x. [ Links ]
Şirin. U. (2011). Effects of different nutrient solution formulations on yield and cut flower quality of gerbera (Gerbera jamesonii) grown in soilless culture system. Afr J Agric Res, 6(21), 4910–4919. [ Links ]
Taiz. L. Zeiger. E. (2006). Plant Physiology. 4th ed. Sinauer Associates, Inc. Publishers. Sunderland, Massachussets. USA. 764 p. [ Links ]
Toth. V. Meszkaros. I. Veres. S. Nagy. J. (2002). Effects of the available nitrogen on the photosynthetic activity and xanthophyll cycle pool of maize in field. J Plant Physiol, 159(6), 627–634. doi: 10.1078/01761610222260879. [ Links ]
Zheng. Y. Graham. T. Richard. S. Dixon. M. (2005). Can low nutrient strategies be used for pot gerbera production in closed–loop sub– irrigation? Acta Hort, 691(1), 365–372. [ Links ]

