











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La macrofauna del suelo representa una parte importante de la biodiversidad de los ecosistemas y agroecosistemas e influye en la naturaleza y el funcionamiento de los suelos (Lavelle, et al., 2006). Estos macroinvertebrados desempeñan un papel importante modificando la fertilidad mediante el aumento de la macroporosidad, la humificación y la mineralización de la materia orgánica; procesos que ayudan a mejorar la estructura del suelo, control de la erosión, intercambio gaseoso, infiltración, retención de agua, propiedades hidráulicas, propiedades químicas y nutricionales del suelo (Lavelle et al., 2006; Brussaard, Ruiter & Brown, 2007). Por tanto, la pérdida de diversidad de macrofauna edáfica como consecuencia de la deforestación y la conversión de los sistemas naturales para la agricultura intensiva puede afectar los procesos ecológicos y la sostenibilidad del agroecosistema a largo tiempo (Wall et al., 2012).
En la Amazonia colombiana la ganadería es la actividad predominante en términos de superficie de la tierra y la principal motivación de la deforestación, para el periodo 2012-14 se encontró una pérdida de 358,000 ha de bosque, equivalente a 179,000 ha por año, lo que es debido a la intervención antrópica de esa área de bosques (Murcia, Gualdrón & Londoño, 2016). Frente a este panorama se han venido promoviendo los arreglos agroforestales como sistemas productivos sostenibles por su cobertura vegetal continua, diversificación de la producción y ciclaje de nutrientes (Monroe, Gama- Rodrigues, Gama-Rodrigues & Marques, 2016). No obstante, existe una alta diversidad de arreglos en el marco genérico de los cultivos agroforestales. Por lo anterior, en este estudio se evaluó el efecto de diferentes arreglos agroforestales en la Amazonia colombiana sobre la densidad y la diversidad de la comunidad de macroinvertebrados utilizando como indicadores los efectos en las propiedades del suelo.
Materiales y métodos
Área de estudio
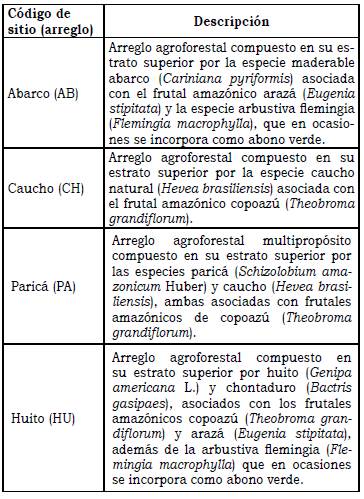
El estudio fue desarrollado en el Centro de Investigaciones Macagual Cesar Augusto Estrada González, situado en el occidente de la Amazonia colombiana (1°37’N y 75°36W) en una región húmeda con una precipitación promedio anual de 3793 mm, un régimen de lluvias monomodal con máxima precipitación entre abril y septiembre), brillo solar de 1707 horas por año, temperatura promedio de 25.5 °C y humedad relativa de 84.3% Las muestras de suelo y macrofauna edáfica fueron recolectadas en cuatro arreglos agroforestales distribuidas aleatoriamente en el centro de investigaciones, con características propias en su distribución espacial y complejidad. Los arreglos fueron identificadas con base en la especie dominante en el estrato superior, así: abarco (AB) Cariniana pyriformis Miers, caucho (CH) Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg., Paricá (PA) Schizolobium amazonicum y Huito (HU) Genipa americana L. (Tabla 1).
Muestreo del suelo y macrofauna edáfica
Cada arreglo agroforestal consistió en un área de 20,000 m2, en los cuales se seleccionaron cuatro lotes de 2500 m2 cada uno, a una distancia entre ellos de 30 m, elegidos por características similares de: grado de pendiente, grado de erosión y tipo de vegetación.
Para el análisis físicoquímico, en cada lote se tomaron muestras compuestas por triplicado a una profundidad de 30 cm sobre el nivel del suelo. En los mismos puntos de muestreo se cuantificó la macrofauna siguiendo la norma ISO 23611-5 (ISO, 2011) para lo cual en un bloque de 25 x 25 x 30 cm se separaron en forma manual los especímenes de macrofauna y hojarasca los que fueron fijados en alcohol para su identificación posterior. En el laboratorio de Biología de la Universidad de la Amazonia los macroinvertebrados fueron separados de acuerdo a su morfología y clasificados en 13 grandes grupos. El material biológico fue depositado en el Museo de Historia Natural de la Universidad de la Amazonia.
Diseño y análisis estadístico
Para evaluar la relación entre comunidades de macroinvertebrados y propiedades del suelo se utilizaron tres parcelas distribuidas en diferen tes arreglos agroforestales con tres repeticiones distribuidas al azar en cada parcela. Los datos de densidad y riqueza de macroinvertebrados fueron sometidos a un análisis de varianza y comparación de medias utilizando la prueba LSD de Fisher (P < 0.05). Los taxones (Blattodea, Der- maptera, Homoptera, Orthoptera, Raphidioptera) que presentaron baja densidad en los arreglos agroforestales fueron agrupados en la categoría ‘Otros’. Sin embargo, fueron tenidos en cuenta en el cálculo de la riqueza específica.
Para cada matriz de datos i = Comunidad de macroinvertebrados e ii = Propiedades edáficas se realizó un Análisis de Componentes Principales (ACP) para determinar similitud entre arreglos agroforestales y explorar la relación entre variables de un mismo conjunto de datos. Previo al ACP de macrofauna, los datos fueron transformados por Log10(x+1) para reducir el efecto de grupos dominantes. La separación de los arreglos agroforestales se hizo con un test de Monte Carlo. Se realizó un análisis de Co-inercia para probar covariaciones entre matrices de datos (Dray, Dufour & Chessel, 2007); la significancia de los valores del Co-inercia se evaluó con un test de Monte Carlo. Los análisis multivariados se realizaron en el software R 3.4.0 (R Core Team, 2016) mediante el paquete estadístico Ade4.
Resultados
Macrofauna edáfica
Fueron recolectados 1727 individuos, siendo los grupos más abundantes Oligochaeta con 990 y Hymenoptera con 189, que representan respec tivamente 52.3% y 10.9% del total de individuos. La densidad máxima se presentó en el arreglo AB (945 ± 382 individuos/m2) y la mínima en el arreglo CH (487 ± 80 individuos/m2) aunque no presentaron diferencias significativas entre ellos (Tabla 2). La riqueza estimada fue de 13 grupos taxonómicos, clasificados en 11 órdenes, 1 clase y 1 subclase. El arreglo PA presentó la mayor riqueza con 6.6 ± 0.8 taxones, en promedio, por cada monolito. El arreglo AB con 4.8 ± 0.8 taxones presentó la menor riqueza (Tabla 2), aunque la diferencia entre ambos igualmente no fue significativa. En resumen, no se observó un efecto definido del arreglo agroforestal sobre la densidad y riqueza de macroinvertebrados (Tabla 2).
Tabla 2 Abundancia de macroinvertebrados presentes en diferentes arreglos agroforestales de la Amazonia colombiana.
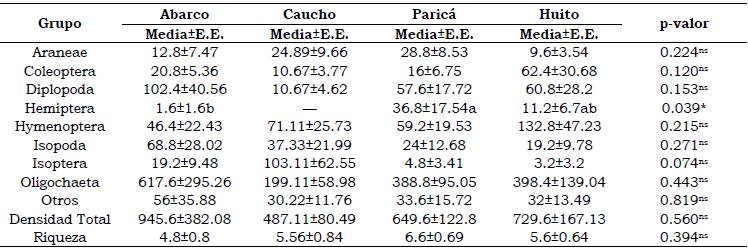
Medias con letras iguales en las filas no difieren entre sí mediante la prueba LSD Fisher con una probabilidad < 0.05. ns no significativo, * significativo al 5% de probabilidad. E.E.: error estándar.
El ACP con los datos de la comunidad de macrofauna explicó 46.7% de la variabilidad total de los datos con los dos primeros ejes. El eje 1 explicó el 29.6%, representando un gradiente de abundancia de los diferentes macroinvertebrados ubicados en lado izquierdo del círculo de correlación (Figura 1). El eje 2 explicó el 17.1% y separó los sitios con mayor abundancia de Araneae e insectos sociales (Isoptera e Hymenoptera) de sitios con mayores densidades de Diplopoda (Figura 1). Las diferencias en las comunidades de macroinvertebrados según el arreglo agroforestal no resultan significativas (test de Monte Carlo) (Figura 1).
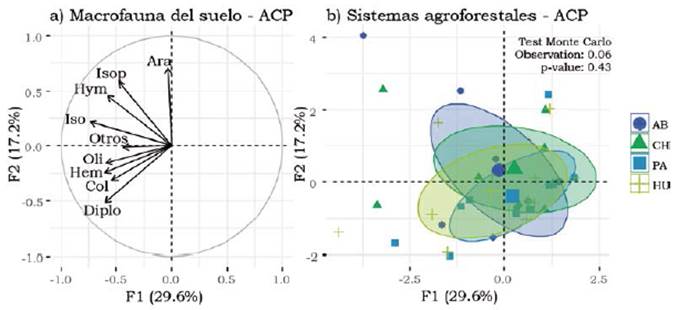
Figura 1 Proyección en el plano factorial F1/F2 de un análisis de componentes principales (ACP) de variables (macroinvertebrados) y de los puntos de muestreo agrupados según el tipo de arreglo agroforestal. (a) macroinvertebrados, círculo de correlación. Ara: Araneae, Col: Coleoptera, Oli: Oligochaeta, Hem: Hemiptera, Hym: Hymenoptera, Iso: Isopoda, Isop: Isoptera, Diplo: Diplopoda y Otros. b) Ordenación de los arreglos agroforestales incluidas en la muestra del plano factorial de un PCA. AB= Abarco, CH= Caucho, PA= Paricá y HU= Huito.
Propiedades del suelo
Los dos primeros ej es del ACP de las propiedades del suelo explicaron el 77% de la variabilidad total de los datos. El eje 1 explicó el 63.8%, separando aquellos arreglos agroforestales como AB y HU que presentaron los niveles más altos de fósforo, pH, calcio y magnesio, de los arreglos CH y PA que presentaron mayores niveles de arena y aluminio (Figura 2). El eje 2 explicó el 13.2% y separó los sitios que presentaron mayor carbono orgánico (CO) de aquellos con mayor contenido arcilla (Arc) (Figura 2). En general, en el plano factorial se encontró un agrupamiento de algunos arreglos agroforestales como AB y HU por presentar altos contenidos de bases intercambiables (P < 0.05, Figura 2). Por la ubicación espacial de los arreglos, se presentaron diferencias texturales separando los arreglos AB y HU de CH y PA; no obstante, cuando se exploran datos a nivel de parcela por cada arreglo agroforestal, no se encontraron diferencias significativas (P > 0.05).
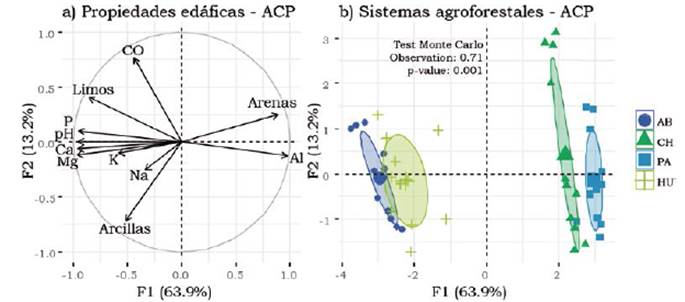
Figura 2 Proyección en el plano factorial F1/F2 de un análisis de componentes principales (ACP) de variables (propiedades del suelo) y de los puntos de muestreo agrupados según el tipo de arreglo agroforestal. (a) Propiedades del suelo, círculo de correlación. Al: aluminio, pH, CO: carbono orgánico, Ca: calcio, Mg: magnesio, K: potasio, Na: sodio y P: fósforo. (b) ordenación de los arreglos agroforestales inclui das en la muestra del plano factorial de un ACP. AB= Abarco, CH= Caucho, PA= Paricá y HU= Huito
Análisis de Co-inercia
La relación entre comunidades de macroinvertebrados y propiedades del suelo bajo los diferentes arreglos agroforestales estudiados, utilizando el análisis de Co-inercia, mostró un coeficiente de correlación significativo (RV: 0.13, P < 0.05 -Figura 3). Los coleópteros presentaron una asociación positiva con Ca, Mg, P y pH, pero negativa con Al a lo largo del eje 1; los taxones Araneae, Hymenoptera, Hemiptera e Isoptera se asociaron positivamente con el K en el lado izquierdo del eje 1. El eje 2 relacionó los diplopodos con el contenido de arena y también a las lombrices e isópodos con el carbono orgánico.
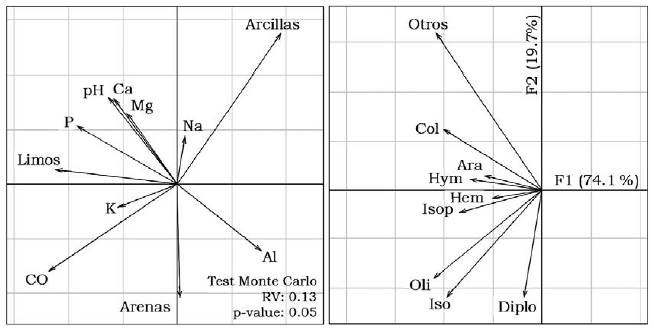
Figura 3 Proyección en el plano factorial F1/F2 de un análisis de Co-inercia de variables propiedades edáficas y variables macrofauna. Ara: Araneae, Col: Coleoptera, Oli: Oligochaeta, Hem: Hemiptera, Hym: Hymenoptera, Iso: Isopoda, Isop: Isoptera, Dipl: Diplopoda y Otros; Al: aluminio, pH, CO: carbono orgánico, Ca: calcio, Mg: magnesio, K: potasio, Na: sodio y P: fósforo.
Discusión
La densidad de la macrofauna edáfica en los arreglos agroforestales estudiados presentó valores más bajos que los reportados para los bosques amazónicos (5301 individuos/m2), pero superiores a los de pasturas degradadas (21.3 individuos/m2) (Marichal et al., 2014), lo que demuestra que los efectos negativos de la intensificación del uso de la tierra sobre las comunidades de macroinvertebrados en el suelo pueden ser reducidos por prácticas agroforestales.
La riqueza de la comunidad de macrofauna no presentó diferencias entre los arreglos agroforestales, posiblemente por las condiciones tróficas y de microclima similares entre ellos, las cuales se obtienen al desarrollar prácticas comunes como la inclusión de árboles de sombra, el mantenimiento de la cobertura y la ausencia de prácticas como labranza y quema, que favorecen la abundancia y diversidad de la macrofauna (Pauli, Barrios, Conacher, y Oberthür, 2011). En este sentido, Lavelle et al. (2016) demostraron que los arreglos agroforestales en paisajes deforestados de la Amazonia son una alternativa que permite obtener una producción sostenible y mantener la biodiversidad y servicios ecosistémicos del suelo; asimismo, Marichal et al. (2014) encontraron que estos arreglos pueden favorecer la densidad de la macrofauna en paisajes amazónicos, la cual disminuye a lo largo de un gradiente de deforestación y la intensificación del uso del suelo.
Los resultados del análisis Co-inercia confirmaron la hipótesis que los grupos de macrofauna son afectados por las propiedades del suelo, pero estos, a su vez, también pueden influir en estas propiedades. Esta relación ha sido estudiada por investigadores como Velásquez, et al. (2012); Vasconcellos, Segat, Bonfim, Baretta y Cardoso (2013); Marichal et al. (2014); y Oliveira, Gama-Rodrigues, Gama-Rodrigues y Sales (2018).
Según Vasconcellos et al. (2013) la relación negativa del Al con la abundancia de Coleópteros puede ser empleada como indicador de la macrofauna, debido al efecto de este elemento sobre los nutrientes y la calidad del suelo; Geissen y Brümmer (1999) concluyeron que un mayor suministro de nutrientes y una disminución del contenido de Al presente en suelos forestales, conducen a una mayor actividad biológica relacionada con la alta descomposición de la materia orgánica.
Los nutrientes Ca, Mg y P, así como sitios con menor nivel de acidez (pH) estuvieron relacionados con la alta presencia de coleópteros. Al respecto, Auclerc et al. (2012) encontraron que la aplicación de cal en el suelo favorece la abundancia de Coleópteros, principalmente la familia Curculionidae, como resultado de los cambios en las concentraciones de Ca, Mg y pH. Li, Ji, Schaffer y Brune (2006) encontraron un aumento significativo de P en el suelo como resultado de la solubilización y la hidrólisis enzimática del P orgánico por la actividad de los coleópteros.
La relación entre Isópodos y carbono orgánico se explica por la actividad de estos organismos en el proceso de descomposición de la materia orgánica, ya que fraccionan la hojarasca (Marques, Silva, Silva, Moreira y Pinto, 2014), estimulan y consumen hongos y bacterias (Frasson, Rosado, Elias y Harter-Marques, 2016), procesos que son fundamentales para el ciclaje de nutrientes.
La distribución de carbono orgánico influye de manera positiva en la presencia y abundancia de lombrices (Kuntz et al., 2013); por su parte, la forma y los mecanismos como estos macroorganismos influyen sobre las concentraciones de este compuesto se explican por la asignación del carbono en microagregados (250 - 253 ^m) recién formados en el horizonte orgánico (Knowles, Ross y Gorres, 2016) y su participación activa en la descomposición de la materia orgánica (Filser et al., 2016).
Moutinho, Nepstad & Davidson (2003) al evaluar la influencia de las colonias de hormigas cortadoras sobre las propiedades del suelo en la Amazonia, encontraron que los desechos orgánicos de Atta sp. (Linnaeus, 1758) incrementaron significativamente la concentración de K, Ca y Mg. Sanabria, Dubs, Lavelle, Fonte y Barot (2016) encontraron que el N total en el suelo se relacionó positivamente con Crematogaster longispina (Emery, 1890), lo que es debido al aumento de la concentración de C y minerales por la presencia de hormigas.
Rousseau, Deheuvels, Rodrigues y Somarriba (2012) encontraron que las arañas del género Pardosa (Koch, 1847) están asociadas con suelos de buena fertilidad, alto contenido de materia orgánica y macronutrientes. De Bruyn & Conacher (1990) y Mujinya, Ranst, Verdoodt, Baert y Ngongo (2010) encontraron mayores concentraciones de nutrientes, en especial K, y mayor capacidad de intercambio catiónico, debido a la concentración y posterior descomposición de la materia orgánica en montículos de termitas. Estos macroorganismos poseen un tracto digestivo con diferentes procesos químicos y biológicos que pueden alterar la carga de iones (Sarcinelli et al., 2009) y favorecer la acumulación de K en forma de carbonato de potasio (Brune yKühl, 1996).
Conclusiones
Las propiedades del suelo bajo los arreglos agroforestales estudiados afectaron la diversidad de macroinvertebrados y son potenciales reguladores de la abundancia de estos organismos en el suelo. En el estudio, los arreglos agroforestales evaluados no afectaron la densidad de individuos/m2 en el suelo.

