











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroducción
La palma de aceite se cultiva debido a la producción de aceite (mesocarpio y almendra) contenido en los racimos de fruta fresca (RFF) (Corley y Tinker, 2015). El rendimiento de RFF lo determina el número de racimos y el peso promedio del racimo (Okoye et al., 2008), caracteres influenciados por factores genéticos y ambientales (Corley y Tinker, 2015; Okoye et al., 2008).
La interacción genotipo x ambiente (GxA) es una fuente de variación importante, porque según su magnitud, puede reducir la tasa de progreso de selección y también el uso de cultivares en áreas específicas (Kang y Gauch, 1996). Su análisis es útil para determinar “la expresión genotípica diferencial a través de los ambientes” (Fox et al., 1997), partiendo del conocimiento del efecto ambiental en la adaptación y el comportamiento varietal (Vallejo y Estrada, 2002). La interacción GxA es evaluada mediante diferentes metodologías, entre ellas: análisis de regresión conjunta (Eberhart y Russell, 1966), componentes de varianza (Shukla, 1972), método de efectos aditivos principales e interacciones multiplicativas (AMMI) (Gauch, 2006), modelo de regresión de sitios (SREG) (Crossa y Cornelius, 1997; Crossa et al., 2002) y GGE Biplot Analysis (Yan y Tinker, 2006). En Elaeis guineensis Jacq. la interacción GxA ha sido evaluada en el rendimiento y sus componentes (Obisesan y Fantula, 1983; Rosenquist, 1981; Rafii et al., 2001; 2012; 2013; Okoye et al., 2008), así como en la producción de materia seca en diferentes entornos con déficit de humedad (Ubara et al., 2017).
El objetivo del estudio fue estimar la interacción GxA a través del modelo AMMI, con respecto al rendimiento de RFF (kg/parcela) de siete genotipos Ténera y nueve del híbrido OxG de palma de aceite, evaluados durante tres años, en tres localidades de dos zonas palmeras de Colombia. Se evaluaron genotipos de la Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA) y materiales comercialmente disponibles, entre el segundo y cuarto año de su etapa productiva. Esto, en el marco del producto 384 “Caracterización de siete materiales Ténera y nueve híbridos con adaptación específica determinada”, financiado por el Ministerio de Agricultura y Desarrollo Rural de Colombia (MADR).
Materiales y métodos
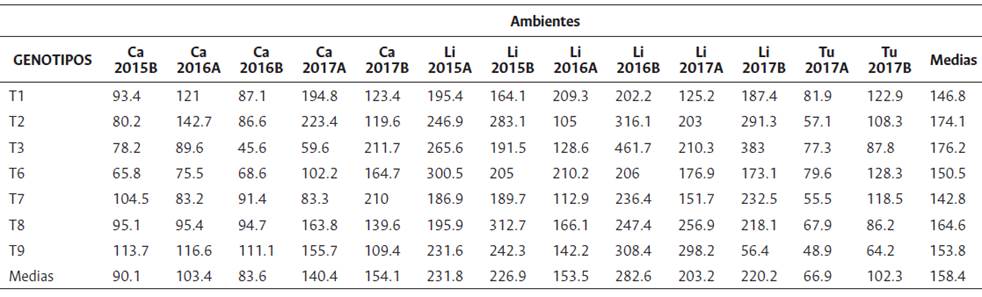
El estudio se realizó en tres localidades colombianas: 1) Plantación Palmas de Tumaco-Zona palmera occidental, Tumaco, Nariño, 2) Centro de Investigación La Libertad-Zona palmera oriental, Villavicencio, Meta y 3) Centro de Investigación Carimagua-Zona palmera oriental, Puerto Gaitán, Meta (Tabla 1). Por localidad se evaluaron siete genotipos Ténera (T1, T2, T3, T6, T7 T8 y T9) y nueve híbridos interespecíficos (H4, H5, H10, H11, H12, H13, H14, H15 y H16). Los genotipos Ténera corresponden a cruzamientos Dura (D) x Pisífera (P) de la especie Elaeis guineensis (G), mientras que los híbridos fueron obtenidos por cruzamiento entre palmas de las especies Elaeis oleifera (O) y Elaeis guineensis (G) en diferentes tipos: cruzamientos OxG para los híbridos H4, H5, H11, H12, H13, H14 y H15, retrocruzamiento (OxG)xG para el híbrido H16 y retrocruzamiento (GxO)xG para el híbrido H10.
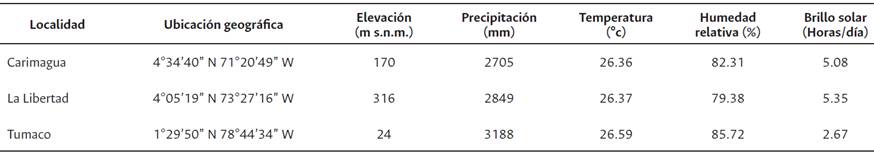
Los genotipos se sembraron en septiembre y octubre de 2010 (La Libertad y Carimagua) y enero de 2011 (Tumaco), en dos ensayos independientes, Ténera e híbridos. Se empleó una distancia de siembra en triángulo de nueve y diez metros entre palma y palma, para una densidad de 143 y 115 palmas/ ha (Ténera y OxG). En cada experimento se utilizó un arreglo de bloques completos al azar con cuatro repeticiones de 20 palmas cada una (cuatro hileras de cinco plantas por material). La producción (kg/ parcela) se estimó con base en los registros de cosecha por palma, tomados de la unidad experimental (seis palmas centrales), en ciclos de 15 y 20 días en promedio (Ténera y OxG). Durante el periodo 2015 a 2017 se contabilizó el número de racimos y su peso. La variable ambiente se definió como la combinación de localidades, años y semestres. Localidades: La Libertad (Li), Carimagua (Ca) y Tumaco (Tu). Años, 2015, 2016 y 2017. Semestres: A y B. Los ambientes Tu2015A, Tu2015B, Tu2016A, Tu2016B y Ca2015A, se excluyeron del análisis por falta de datos y/o datos incompletos.
Debido a diferencias en el número de palmas productivas por unidad experimental (UE) entre genotipos, se realizó un análisis combinado de covarianza por ambiente, para la variable rendimiento de RFF (kg/parcela), considerando el número de palmas como covariable; a través del procedimiento GLM de SAS (v9.4). La comparación de medias entre los genotipos se hizo mediante la prueba de Tukey (P=005). Estos análisis se realizaron independientemente para cada tipo de material genético. En los genotipos Ténera, el número de palmas productivas fue seriamente disminuido por la enfermedad pudrición del cogollo (PC) y por algunas palmas en ciclo masculino; mientras que, en los híbridos OxG fue debido a la cantidad de palmas en ciclo masculino, más que a la enfermedad.
El modelo estadístico utilizado fue el siguiente:
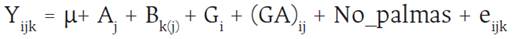
Donde:
Yijk = Rendimiento medio de fruto por palma para cada unidad experimental del i-ésimo genotipo obtenido en el j-ésimo ambiente y el k-ésimo bloque
μ = Efecto de la media general
Aj = Efecto del j-ésimo ambiente
Bk(j)= Efecto del k-ésimo bloque o repetición dentro de cada ambiente
Gi = Efecto del i-ésimo genotipo
(GA)ij. = Efecto de la interacción entre el i-ésimo genotipo y el j-ésimo ambiente
No_palmas = Número de palmas vivas por unidad experimental (Covariable)
eijk = Efecto aleatorio del error experimental asociado al i-ésimo genotipo en el j-ésimo ambiente y el K-ésimo bloque, según el modelo lineal aditivo.
Con base en las medias ajustadas por el número de palmas por UE mediante el análisis de covarianza (ANCOVA), se realizó un agrupamiento de ambientes y de genotipos, para Ténera e híbridos OxG, utilizando el análisis de conglomerados, mediante el procedimiento CLUSTER y el algoritmo de Ward en SAS v9.4. Este procedimiento agrupa jerárquicamente las observaciones (genotipos y ambientes) de un conjunto de datos. El método de varianza mínima de Ward minimiza la varianza dentro de cada grupo y la maximiza entre grupos (Statistical Analysis System [SAS], 2012).
La interacción GxA se descompuso con base en el modelo AMMI (Gabriel, 1971; Crossa et al., 1990; Crossa et al., 2000):
Donde:
Yij = Rendimiento observado del genotipo i-ésimo, en el ambiente j-ésimo, en el bloque k-ésimo
μ = Media general
Gi = Desviación del genotipo i-ésimo de la media general
Aj = Desviación del ambiente j-ésimo de la media general
λn = Valor singular para el eje n-ésimo del CP de la interacción
ϒin = Vector característico del genotipo i-esimo para el eje n-ésimo
δjn = Vector característico del genotipo j-esimo para el eje n-ésimo
ρij = Parte de la interacción que no es explicada por elj ACP
εijk = Desviación del genotipo i-ésimo, en el ambiente j-ésimo de la repetición k-ésima
Cornelius y Seyedsadr (1997), lo definen como modelos lineales-bilineales generales, porque tienen términos aditivos (lineales) y términos multiplicativos (bilineales). Los resultados AMMI se resumen en una gráfica de dos dimensiones (biplot), donde se ubican los efectos principales de los ambientes y los genotipos, y los efectos de la interacción GxA.
Resultados
Genotipos Ténera
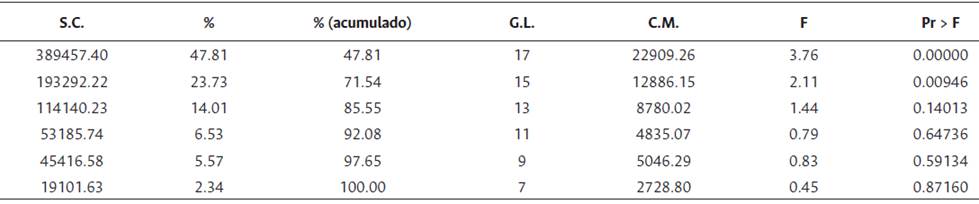
El ANCOVA indicó un efecto significativo del número de palmas productivas (en ciclo productivo) por UE, sobre el rendimiento de RFF (kg/parcela). Se detectaron diferencias significativas entre los ambientes (a < 0.0001), no se presentaron diferencias significativas entre los genotipos, pero hubo efecto significativo de la interacción GxA (a < 0.0001). El ambiente explicó el 33.1 % de la variabilidad total, los genotipos el 1.1 % y la interacción GxA el 17.3 % (Tabla 2). Las Figuras 1 y 2 representan el agrupamiento de los genotipos y de los ambientes.
Tabla 2 ANCOVA por ambiente (genotipos Ténera)
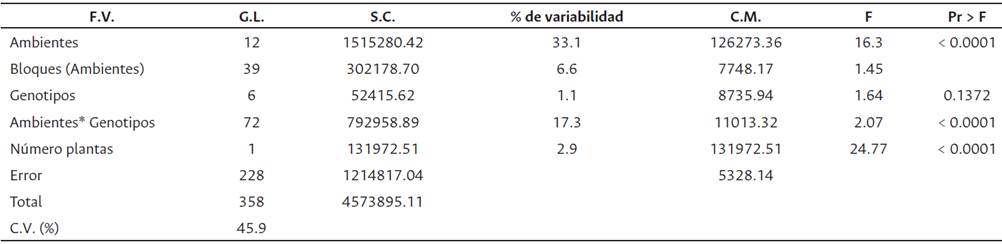
F.V.: Fuente de variación; G.L.: grados de libertad; S.C.: suma de cuadrados; C.M.: cuadrados medios; F: F calculado; Pr > F es la significancia.
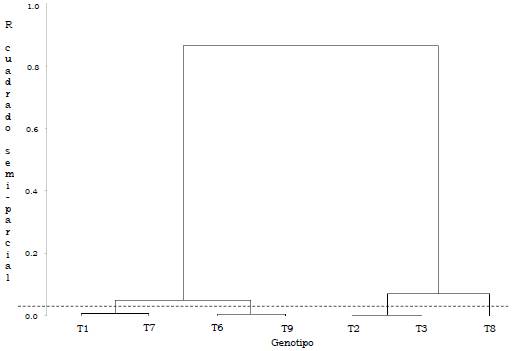
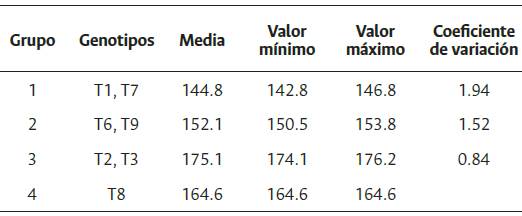
Los genotipos Ténera se clasificaron en cuatro conglomerados, al primero lo conforman T1 y T7, presentando el rendimiento medio más bajo (144.8 kg/parcela). Los genotipos T6 y T9 (152.1 kg/ parcela) constituyen el segundo grupo, mientras que T2 y T3 el tercer grupo, con mayor rendimiento medio de RFF (175.1 kg/parcela). El genotipo T8 representa el cuarto grupo (164.6 kg/parcela) (Tabla 3).
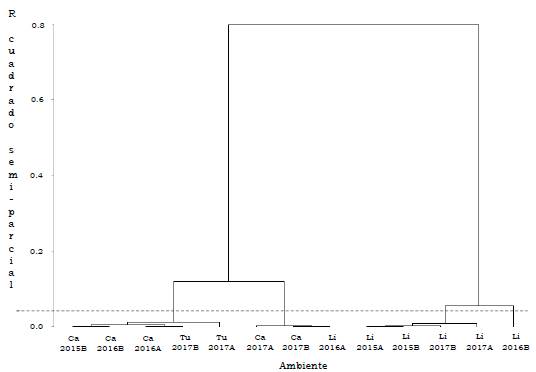
La clasificación general de los ambientes para genotipos Ténera, revela cuatro conglomerados, el primero lo conforman los ambientes Ca2015B, Ca2016A, Ca2016B, Tu2017A y Tu2017B con un rendimiento medio de 89.3 kg/parcela. Los ambientes Ca2017A, Ca2017B y Li2016A (149.3 kg/ parcela) constituyen el segundo grupo y al tercero lo conforman Li2015A, Li2015B, Li2017A y Li2017B (220.5 kg/parcela). El cuarto grupo lo representa Li2016B (282.5 kg/parcela), con mayor rendimiento medio (Tabla 4). De acuerdo con la prueba de Gollob (1968), para los genotipos Ténera evaluados, el primer componente representó el 47.8 % de la varianza total y el segundo el 23.7 %, para un acumulado del 71.5 % en los dos primeros términos AMMI (Tabla 5).
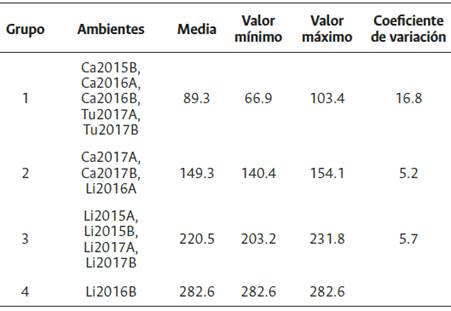
Tabla 5 Sumas de cuadrados y cuadrados medios para los términos AMMI (Prueba de Gollob)
F.V.: Fuente de variación; G.L.: grados de libertad; S.C.: suma de cuadrados; C.M.: cuadrados medios; F: F calculado; Pr > F es la significancia.
Los genotipos T1, T3 y T9 presentaron mayor contribución a la interacción GxA, con rendimientos medios de 146.8, 176.2 y 153.8 kg/parcela. T3 exhibe el mayor valor con 158.4 kg/parcela y T7 presentó bajo rendimiento (142.8 kg/parcela) (Figura 3 y Tabla 6). Los ambientes Li2016A, Li2016B, Li2017A, Li2017B, Ca2017A, Ca2017B y Tu2017B, fueron los que más contribuyeron en esta interacción, siendo Li2016B, el de mayor rendimiento medio (282.6 kg/ parcela).
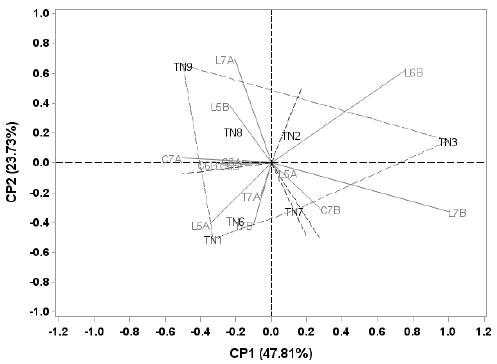
En la Figura 3 se identifican tres sectores delimitados por las líneas punteadas, en el primer sector se ubican los ambientes Li2015B, Li2017A, Ca2015B, Ca2016A, Ca2016B y Ca2017A, la mayoría correspondientes a Carimagua, cuyos rendimientos medios son inferiores a la media general, en cuanto ambientes se refiere. El genotipo T9 (153.8 kg/ parcela) fue el que mejor se adaptó en dichos ambientes, especialmente en Li2017A. En el segundo sector se ubican los ambientes Tu2017A, Tu2017B y Li2016A. En Li2016B y Li2017B se presentaron altos valores de rendimiento de RFF (282.6 y 220.2 kg/ parcela); el genotipo T3 (176.2 kg/parcela) fue el de mejor adaptación.
Híbridos interespecíficos OxG
El ANCOVA reveló un efecto significativo del número de plantas productivas por UE. Entre ambientes, entre genotipos y en la interacción GxA se detectaron diferencias significativas (a < 0.0001). El ambiente explicó el 35.5 %, de la variabilidad total, los genotipos el 7.0 % y la interacción el 13.6 % (Tabla 7). La Figura 4 y la Figura 5 representan el agrupamiento de los genotipos y de los ambientes.
Tabla 7 ANCOVA por ambiente (genotipos OxG)
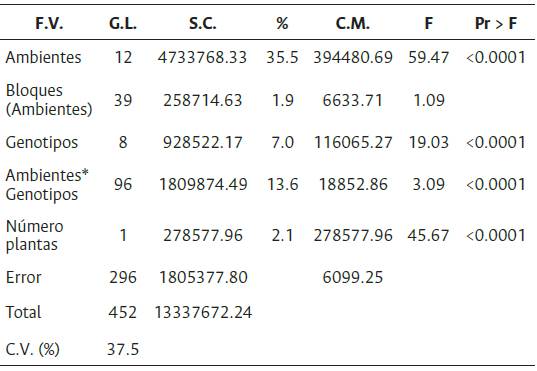
F.V.: Fuente de variación; G.L.: grados de libertad; S.C.: suma de cuadrados); C.M.: cuadrados medios; F: F calculado, Pr > F es la significancia.
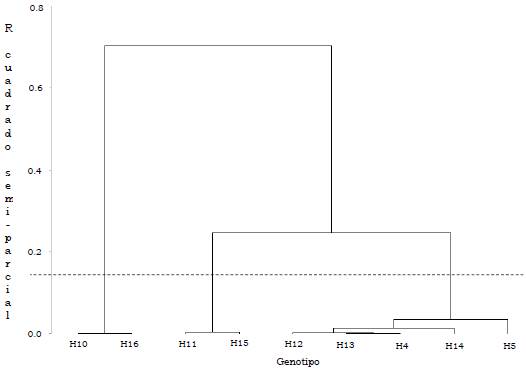
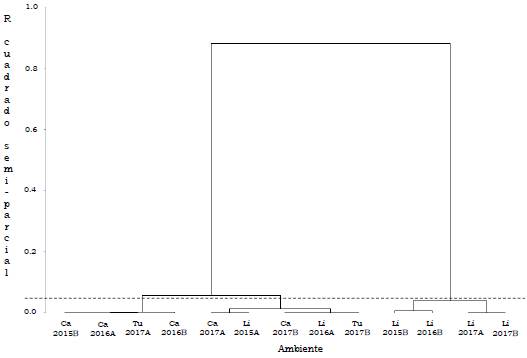
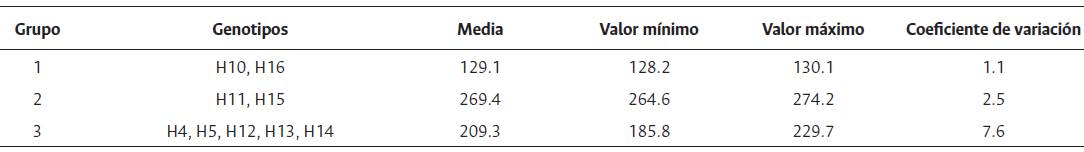
Los genotipos OxG se congregaron en tres grupos: el primero, conformado por H10 y H16, presentó el menor rendimiento medio (129.1 kg/parcela), mientras el segundo grupo, constituido por H11 y H15 el mayor rendimiento medio (269.4 kg/parcela). Los híbridos H4, H5, H12, H13 y H14 integran el tercer grupo, con 209.3 kg/parcela de RFF (Tabla 8).
Tabla 8 Estadísticas básicas de los grupos correspondientes a los genotipos OxG (valores en kg/parcela)
En la clasificación general de los ambientes para genotipos OxG se conformaron tres grupos: el primero constituido por los ambientes Ca2015B, Ca2016A, Ca2016B y Tu2017A, con un rendimiento medio de 99.7 kg/parcela, el valor más bajo. El grupo dos conformado por Ca2017A, Ca2017B, Li2015A, Li2016A y Tu2017B (163.9 kg/parcela) y el grupo tres por Li2015B, Li2016B, Li2017A y Li2017B (361.2 kg/parcela), que fue el de mayor rendimiento medio (Tabla 9). De acuerdo con la prueba de Gollob (1968), para los híbridos OxG evaluados, el primer componente representó el 45.2 % de la varianza total y el segundo el 32.9 %, para un acumulado del 78.2 % de los dos primeros términos AMMI (Tabla 10).
Tabla 9 Estadísticas básicas de los grupos de ambientes evaluados para los genotipos OxG (valores en kg/parcela)
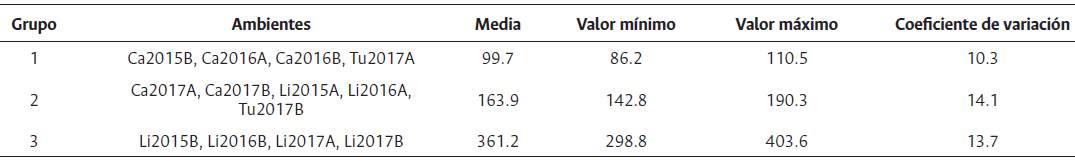
Tabla 10 Sumas de cuadrados y cuadrados medios para los términos AMMI (Prueba de Gollob)
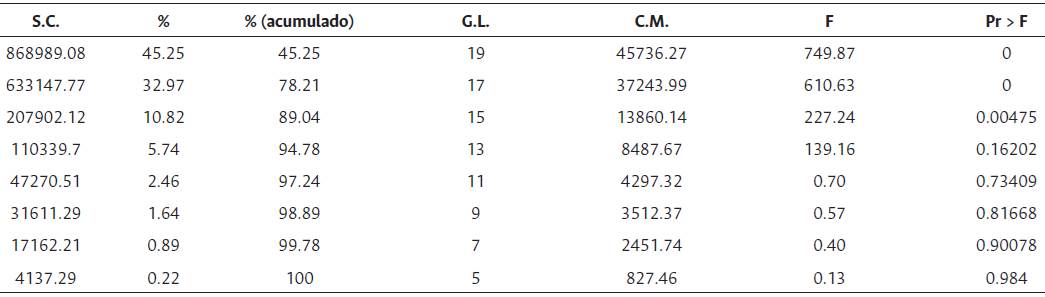
F.V.: Fuente de variación; G.L.: grados de libertad; S.C.: suma de cuadrados; C.M.: cuadrados medios; F: F calculado; Pr > F es la significancia.
Los híbridos H5, H10, H11, H13, H14, H15 y H16, presentaron la mayor contribución a la interacción GxA; H11 (264.6 kg/parcela), H13 (208.3 kg/parcela), H14 (229.7 kg/parcela) y H15 (274.2 kg/parcela), presentaron los mayores rendimientos medios (Figura 6 y Tabla 11). Los ambientes Li2015B, Li2016A y Li2017A, fueron los que más contribuyeron con esta interacción. En la Figura 6 se identifican tres sectores delimitados por las líneas punteadas, en el primero se ubican los ambientes Ca2015B, Ca2016A, Ca2016B, Ca2017B, Li2016A y Tu2017A; en este sector, los híbridos H10 y H16 presentaron los menores rendimientos medios (128.2 y 130.1 kg/parcela). En el segundo sector se ubicó Li2017A, en el cual, H14 presentó el mayor rendimiento medio (229.7 kg/parcela), demostrando buena adaptación. En el tercer sector, quedaron incluidos Li2015A, Li2015B, Li2016B, Li2017B, Ca2017A y Tu2017B. El híbrido H11 fue el de mejor adaptación en Li2017B, Li2015B y Li2016B por su mayor rendimiento, mientras que el híbrido H15 se adaptó mejor en Li2017B, con el segundo rendimiento más alto.
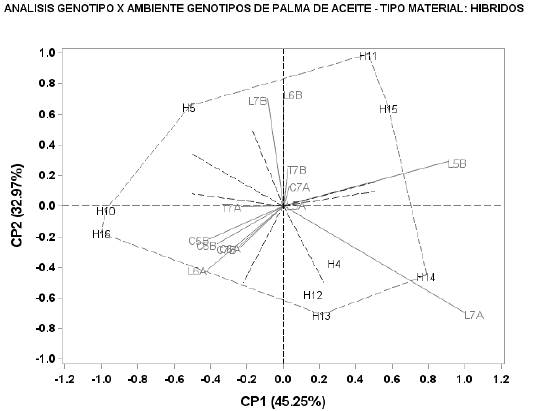
Tabla 11 Medias ajustadas del rendimiento de RFF para cada ambiente y para cada genotipo Híbrido OxG
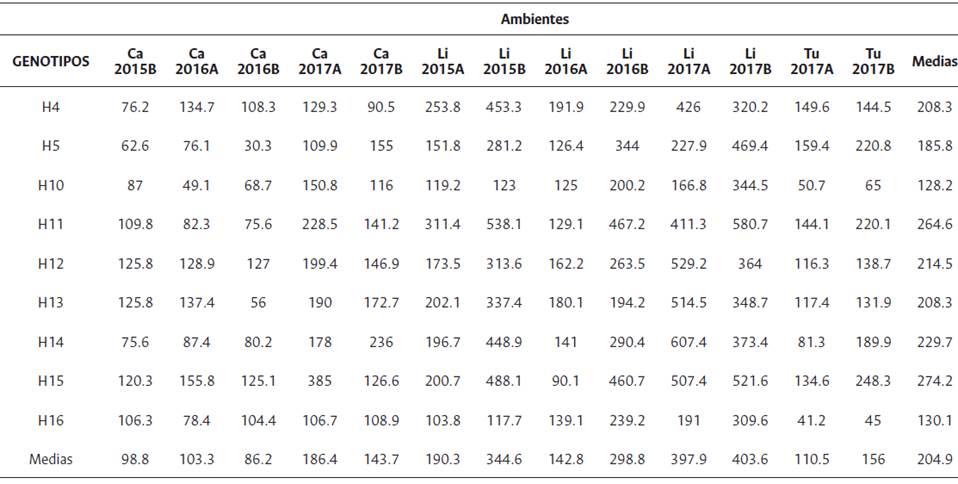
Entre los años 2015 y 2017, los híbridos H10, H11, H15 y H16 presentaron rendimientos variables. El híbrido H16 (retrocruzamiento) presentó los menores valores de rendimiento en las tres localidades. El híbrido H11 (F1) se destaca en La Libertad y el híbrido H15 (F1) lo es en Carimagua y Tumaco. En la Orinoquía, los mayores rendimientos (kg/parcela) para los híbridos OxG se obtuvieron en los ambientes Li2015B, Li2016A, Li2016B, Li2017A y Li2017B.
Discusión
Durante la investigación los genotipos Ténera e híbridos OxG presentaron comportamiento variable en cuanto a rendimiento de RFF (kg/parcela), debido a la interacción GxA (a < 0.0001) de tipo cualitativo; al respecto, Vallejo y Estrada (2002) aseguran que una interacción cualitativa genera cambios en el mérito de los genotipos en los diferentes ambientes. En híbridos OxG se observaron diferencias significativas (a < 0.0001) en ambientes, genotipos y en la interacción GxA para la variable rendimiento (kg/parcela), mientras que en materiales Ténera las diferencias significativas (a < 0.0001) se presentaron en ambientes y en la interacción GxA.
Estos resultados confirman los hallazgos en otras investigaciones, por ejemplo, Rafii et al. (2001); 2012) reportaron diferencias significativas en genotipos y ambientes para las variables rendimiento de aceite y sus componentes, en 40 y 38 progenies DxP, evaluadas en seis y cuatro localidades de Malasia. Okoye et al., (2008) obtuvieron diferencias significativas (p < 0,001) en el genotipo y el periodo de evaluación, para las variables rendimiento de RFF, número de racimos y peso promedio del racimo, en 15 genotipos (Deli x Ténera) evaluados en Nigeria durante 1999-2002. Bueraheng et al., (2018) observaron que las varianzas atribuidas al ambiente, el genotipo y a la interacción GxA, fueron significativas (p < 0,001) para seis progenies E. guineensis Ténera evaluadas en varias localidades del sur de Tailandia, con respecto a la estabilidad del rendimiento de aceite y sus componentes. Según los autores, el rendimiento de cada progenie evaluada depende del ambiente específico en el cual se establece.
Los genotipos Ténera e híbridos OxG presentaron variación en el rendimiento en algunos ambientes con respecto al periodo de registro, atribuible a las diferencias climáticas durante los procesos fisiológicos de formación de inflorescencias. Los ciclos de floración, masculinos o femeninos, obedecen a la influencia de factores externos como: temperatura, luminosidad, estado nutricional, precipitación y genotipo (Corley y Tinker, 2015; Bastidas et al., 2013).
Según Romero et al., (2007) el rendimiento de palma de aceite está determinado por la radiación solar fotosintéticamente activa, la fracción de esta interceptada por el dosel, la eficiencia fotosintética del cultivo, el índice de cosecha y la interacción entre la temperatura durante el periodo de crecimiento y la disponibilidad de agua (precipitación o riego). Al respecto, el comportamiento diferencial de los genotipos Ténera e híbridos OxG en el rendimiento de RFF, obedece a la variación presentada en precipitación, temperatura y brillo solar, entre las localidades Carimagua, La Libertad y Tumaco, durante el periodo de estudio (2015-2017). Según Bastidas et al., (2013), la producción de fruto en los híbridos OxG depende en mayor proporción de los factores ambientales (clima, suelo, nutrientes, plagas, enfermedades, manejo agronómico, riego, drenajes, etc.) y en menor proporción del genotipo.
Conclusiones
En genotipos Ténera e híbridos OxG el rendimiento de RFF fue alterado por la interacción GxA, la cual contribuyó con el 17.3 de la variabilidad de los genotipos y con el 13.6 % en híbridos OxG. En contraste, al ambiente se le atribuye el 33.1 % de la variabilidad de los genotipos Ténera y el 35.5 % en los híbridos OxG.
La variabilidad en cuanto a la producción de RFF fue mayor en los genotipos Ténera, puesto que los siete genotipos Ténera conformaron cuatro grupos diferentes, mientras que los nueve genotipos del híbrido OxG se agruparon en tres conglomerados.
El análisis de interacción GxA por el método AMMI denota que los genotipos Ténera T6 y T1 presentaron adaptación específica en los ambientes Tu2017B y Li2016A. Se destacan los genotipos T2 y T3 e híbridos OxG H11 y H15 por los mayores rendimientos de RFF (kg/parcela) en los ambientes evaluados.

