











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroducción
La transformación de ecosistemas naturales en modelos productivos agrícolas, la deforestación, el sobrepastoreo, la urbanización y la industrialización alteran notablemente los procesos biológicos, físicos y químicos del suelo (Zia-ur-Rehman et al., 2016). Para la zona andina, la agricultura intensiva de cultivos anuales y la ganadería extensiva en zonas de ladera se asocian a la degradación de la estructura del suelo, simplificación de hábitats y pérdida del suelo por procesos erosivos (Galindo et al., 2017; 2022). La disminución de la diversidad de la vegetación en los sistemas agropecuarios también conduce a una disminución notable de macrofauna y microorganismos del suelo, lo cual modifica los procesos biológicos relacionados con la regulación de ciclo de nutrientes (carbono y nitrógeno) y la agregación del suelo (Lavelle et al., 2020).
Los hongos formadores de micorriza arbuscular (HMA) pueden formar simbiosis con más del 80 % de las plantas terrestres, por ello se les considera como los organismos más importantes involucrados en la estabilización de los agregados (Six et al., 2004). Los HMA pueden intervenir directamente en la agregación del suelo por medio de mecanismos biofísicos, bioquímicos y biológicos tales como la acción mecánica de las hifas que sirven para enredar las partículas de menor tamaño y mediante la secreción de sustancias adherentes (e.g. glomalina) e interacciones con otros organismos, además del incremento de la superficie de absorción de las raíces de las plantas hospederas (Rillig y Mummey, 2006).
Muchos de los esfuerzos para comprender las interacciones entre los HMA y la calidad del suelo en las últimas dos décadas, se han enfocado en suelos de zonas templadas y de importancia agrícola, bajo condiciones de laboratorio, con suelos estériles y plantas inoculadas (Wu et al., 2014; Muchane et al., 2019). En el trópico, las HMA son el foco de estudios, especialmente en regiones agrícolas, porque intervienen en los procesos ecológicos de rehabilitación de suelos con limitaciones de nutrientes, puesto que los filamentos de las hifas mejoran la eficiencia en el uso del agua y la absorción de nutrientes en las plantas cultivadas, y porque intervienen en la agregación y la estabilidad estructural del suelo (Picone, 2000; Lavelle et al., 2020).
Debido a que gran parte de los trabajos sobre HMA se enfocan en la relación con la formación de macroagregados en áreas agrícolas, este trabajo busca evaluar el cambio en las principales características de los HMA que intervienen en la formación de micro y macroagregados y su relación con las coberturas de vegetación y los principales modelos agropecuarios presentes en la cuenca del río Cali, Colombia, donde es importante el control de la erosión. Esta investigación aporta información para contribuir a comprender los procesos edáficos, en especial las interacciones entre el suelo y los HMA.
Materiales y métodos
Zona de estudio
El estudio se realizó en la zona de Reserva Forestal de la cuenca del río Cali, en el Valle del Cauca, Colombia, que cuenta con 7481 ha, y entre 1300 y 2300 m s. n. m. Está situada en la transición entre las zonas de vida Bosque Húmedo Premontano (bh-PM) y Bosque muy Húmedo Premontano (bmh-PM). El río Cali lo conforman las subcuencas del río Felidia y Pichindé, allí presomina un relieve montañoso con valles de poca profundidad, además de una densa red de drenajes de primer y segundo orden. A partir del análisis de coberturas de vegetación predominante, planos y mapas se seleccionaron cuatro coberturas vegetales (Tabla 1).
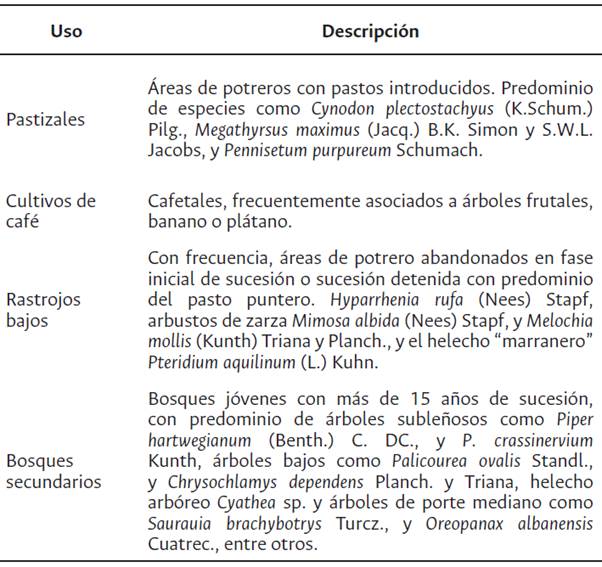
Diseño experimental
Se realizó un diseño estratificado donde se seleccionaron cinco parches en cada una de las cuatro coberturas vegetales, para un total de 20 unidades experimentales (UE), con una separación mínima de 100 m entre cada unidad de muestreo.
Cálculo de las variables de respuesta en cada UE
Para la densidad de esporas, se tomó una muestra de 80 g de suelo de hasta 10 cm de profundidad y se extrajeron las esporas siguiendo el procedimiento de Sieverding (1983) y Sánchez de Práguer et al. (2010). Las muestras se pasaron en tamices superpuestos con 2 mm, 450 μm, 120 μm y 40 μm de tamaño de poro. Luego los contenidos de los tamices fueron centrifugados a 3350 rpm por 4 min, utilizando solución de sacarosa (70 %) y agua. La cuantificación de esporas extraídas se calculó siguiendo la metodología sugerida por International Culture Collection of Vesicular Arbuscular Mycorrhizal Fungy INVAM (2017). El procedimiento para la extracción y cuantificación de micelio externo total se realizó con base a una muestra de 15 g de suelo en un beaker con 250 ml de agua desionizada y 31 ml de hexametafosfato de sodio por 12 horas. La muestra se dispersó en un agitador y se tomaron seis alícuotas de 1 ml mezcladas con 250 ml de agua destilada y 35 ml de hexametafosfato de sodio. Cuatro alícFuotas de 5 ml se depositaron en un equipo de filtración por membrana y una adición de tinta negra en vinagre por diez minutos. Después de retirar el filtro se midió la longitud de micelio por medio del método de Tennant (1975):
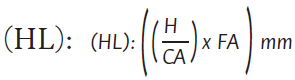
Donde:
H: longitud total de hifas sobre área de filtro
cubierta por 70 campos leídos.
CA: área del filtro cubierta por 70 campos leídos.
FA: área neta de filtrado (210 mm2).
Para calcular la concentración de glomalina fácilmente extraíble (EE-GRSP), glomalina difícilmente extraíble (DE-GRSp) y glomalina total (T-GRSP), se realizó la extracción de las fracciones de las proteínas del suelo relacionadas con la glomalina, por medio del método Bradford, sugerido por Wright y Upadhyaya (1998). Se tomaron muestras de 1 g de suelo mezcladas con 8 ml de citrato de sodio 20 mM pH 7, y fueron luego sometidas a autoclave a 120° C y 15 lb presión/in2 por 30 minutos. Las muestras fueron centrifugadas a 5000 rpm por 15 minutos para extraer el sobrenadante. Para glomalina difícilmente extraíble, se repitieron los pasos anteriores siete veces (citrato de sodio, autoclave y centrífuga). En cada repetición se almacenó el sobrenadante en el mismo frasco y se midió el volumen total recolectado. La glomalina total se calculó sumando la glomalina fácilmente extraíble y difícilmente extraíble.
La glomalina se midió en un espectrofotómetro a 595 nm, preparando previamente una curva estándar con suero albúmina bovina (BSA) factor V en tampón fosfato salino (PBS) pH 7.4 y reactivo Bradford como colorante. El BSA se reemplazó por la solución de glomalina en cantidades entre 1 y 50 μL y se calculó la concentración proyectando el valor sobre la curva estándar. La cuantificación de agregados estables al agua se realizó con el método de Yoder, descrito por Jaramillo (2001). Los parámetros físicos y químicos del suelo se realizaron en cada sitio de muestreo; se tomaron cinco submuestras de suelo de 200 g equidistantes a una profundidad de 0-20 cm, para una muestra final de 1 kg de suelo. Se siguieron los procedimientos de Motsara y Roy (2008), como se describe en Galindo et al. (2022).
Análisis de datos
Se realizaron análisis de varianza y post-Anova Tukey para evaluar las diferencias entre las características químicas y físicas del suelo, el estado de agregación del suelo, y los rasgos de los hongos formadores de micorriza arbuscular entre las coberturas de vegetación. Se usaron análisis de correlación de Pearson y Spearman para variables con distribución normal sin distribución normal, respectivamente. Se realizó un análisis factorial múltiple (AFM) para evaluar la relación entre las variables que describen la formación de macro y microagregados, los mecanismos fisicoquímicos de los HMA y la composición de elementos orgánicos del suelo en diferentes coberturas vegetales. Igualmente, se conformaron cuatro grupos de variables: (1) grupo cobertura, que incluye las coberturas evaluadas. Esta se tomó como variable suplementaria, dado que la cobertura es una variable cuantitativa; (2) grupo rasgos de los HMA; (3) grupo con las variables de los agregados del suelo; y (4) grupo de variables Fisicoquímicas. Los análisis se realizaron con el programa estadístico R (RStudio Team, 2021).
Resultados y discusión
En general, las diferentes variables de respuesta tuvieron tendencias diferentes en las cuatro coberturas de vegetación estudiadas, con implicaciones importantes para la conservación y la prevención de la erosión aguas abajo en esta importante cuenca del río Cali.
Densidad de esporas, micelio externo total y glomalinas
La densidad de esporas, en general, fue tres veces mayor en rastrojo y potrero con respecto a los bosques (P < 0.001) (Tabla 2). Es posible que la mayor densidad de esporas en suelos de potrero esté relacionada con la alta densidad de raíces finas en las gramíneas que, según Picone (2000), es hasta nueve veces mayor que las raíces presentes en los bosques tropicales, y esto puede favorecer la esporulación de HMA. También se conoce que, a mayor temperatura y luminosidad en la superficie de suelos, como la presente en rastrojos y potreros, se incrementa la producción de esporas, especialmente de hongos asociados a hospederos intolerantes a la sombra (Picone, 2000). En los pastizales, la mayor esporulación puede significar estrés (Picone 2000), aunque la alta presencia de esporas permite suponer que el inóculo de HMA no sería limitante para la restauración de pasturas antiguas.
Por otro lado, la longitud de micelio externo (LME) fue mayor en la cobertura de bosque (P = 0.019) y menor en cultivo de café y rastrojo (Tabla 2). Este resultado se asocia a la baja incidencia de agentes físicos (manejo agrícola) que llevan a la perturbación y ruptura de las redes de micelio fúngico y a la ausencia de agentes químicos que restringen el crecimiento de los HMA (Sally y David, 2010). En los suelos de ladera de la cuenca del río Cali se realiza un manejo convencional de control de arvenses con herramientas como azadas, que afectan los diez primeros centímetros de suelo, con la consecuente disrupción de la estructura del micelio. La ruptura del micelio puede afectar negativamente la diversidad de hongos micorrícicos, pues la cantidad de micelio externo generado por los HMA varía según las especies (Muchane et al., 2019).
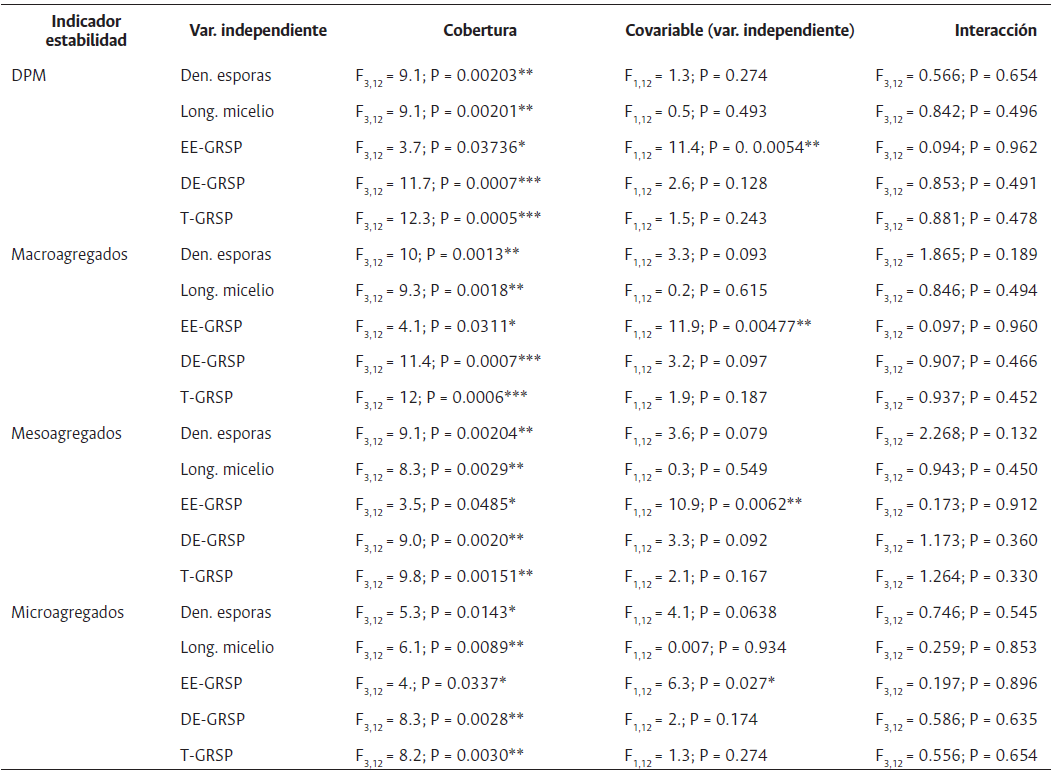
Tabla 2 Efecto de Las coberturas de suelo sobre variables físicas y químicas evaluadas: densidad de esporas (N.° esporas/100g ss), LME (longitud de micelio externo m/g ss), glomalina fácilmente extractable (EE-GRSP), glomalina difícilmente extractable (DE-GRSP), glomalina total (T-GRSP), DPM (diámetro ponderado medio), DGM (diámetro geométrico medio), % microagregados, % macroagregados, % mesoagregados, (Ca, K, mg, Na, Al, CICE (meq/100g)), pH, % CO, % NT, (P, NO3, NH4, (mg/kg))
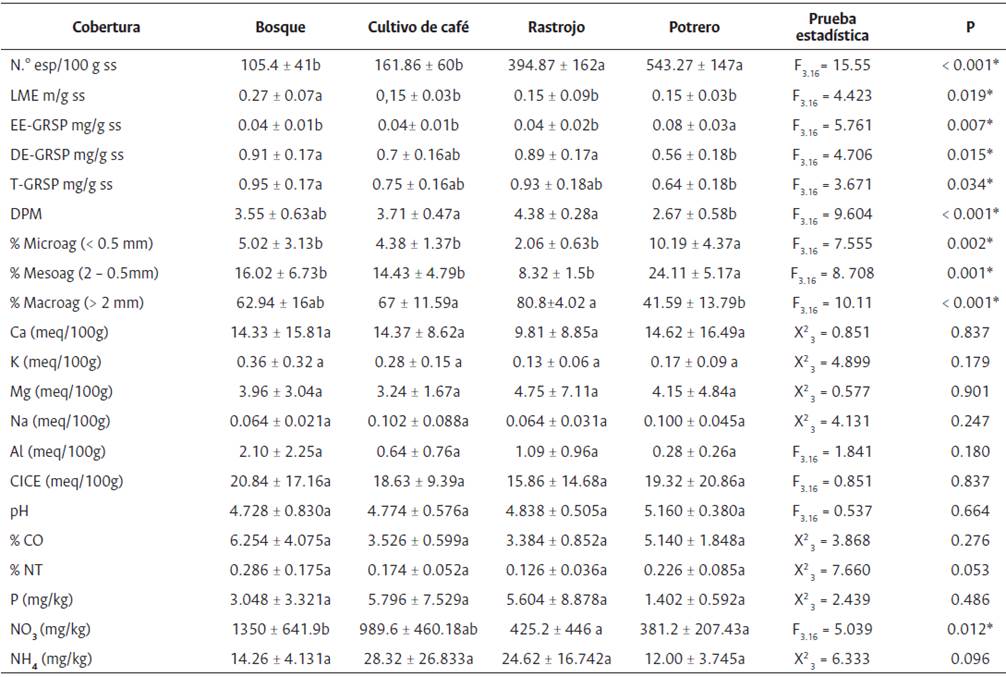
Datos (media ± desviación entre coberturas mediante estándar n = 5), seguidos por letras diferentes son significativamente diferentes (p < 0.05) entre valores p de la misma variable prueba de Tukey
La concentración de glomalina fácilmente extractable (EE-GRSP) fue más alta en los suelos de potrero que en bosque, cultivo de café y rastrojo (P = 0.007) (Tabla 2). Esta fracción se relaciona con la glomalina de más reciente formación, por lo que puede variar según las condiciones climáticas y las prácticas agrícolas de movimiento de suelo, con una mayor degradación asociada a la incidencia directa del sol en suelos con baja cobertura de vegetación leñosa (Holátko et al., 2021). En contraste, la glomalina difícilmente extractable (DE-GRSP) y la glomalina total (T-GRSP) presentaron las concentraciones más altas en los suelos de bosque y rastrojo (Tabla 2 Se interpreta que estas fracciones de proteína (DE-GRSP y T-GRSP) cuentan con mayor tiempo de consolidación y están más reguladas por la acción microbiana y la química del suelo (Koide y Peoples, 2013) lo que concuerda con los resultados.
Cuantificación de agregados estables al agua
El índice de diámetro ponderado medio (DPM) fue más alto en rastrojo, seguido de cultivo de café, bosque y potrero (Figura 1a; Tabla 4). Por otro lado, el porcentaje de los microagregados y mesoagregados fue significativamente mayor en potreros que en las demás coberturas de vegetación (Figuras 1b, 1c, 1d). Las tendencias de los DPM y macroagregados fueron contrarias con respecto a las tendencias con mesoagregados y microagregados, es decir, las pasturas tuvieron menores valores en los dos primeros y significativamente mayores en los dos últimos. La agregación en suelos de bosques, rastrojos y cultivos de café se asemejan por las relaciones favorables para la formación de agregados, tales como la disminución de prácticas de manejo intensivo del suelo y la presencia de vegetación leñosa (árboles y arbustos asociados) (Verbruggen y Toby Kiers, 2010). En contraste, la mayor fracción de microagregados y mesoagregados en suelos de potrero puede asociarse a procesos de degradación de la estructura del suelo, compactación e incidencia de procesos erosivos por efecto del pisoteo continuo del ganado (Mulholland y Fullen, 1991), lo que conduce a la disminución del tamaño de los agregados, al incremento de la escorrentía superficial y al lavado de nutrientes (Pietola et al., 2005, Galindo et al., 2022).
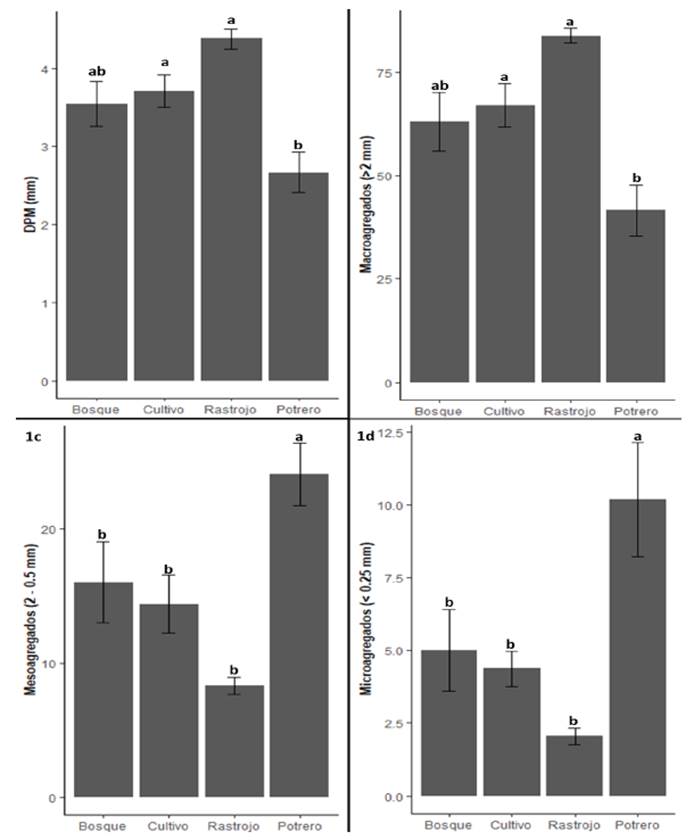
Figura 1 Valores promedios y desviación estándar de diámetro ponderado medio (DPM) (1a), macroagregados (2b), mesoagregados (2c), microagregados (2d). Las letras diferentes indican diferencias significativas entre coberturas.
Parámetros físicos y químicos del suelo.
Los parámetros físicos y químicos del suelo no presentaron diferencias significativas entre las coberturas evaluadas, a excepción de los nitratos (NO3), los cuales fueron significativamente mayores en los bosques y más bajos en los potreros (Tabla 2). Teniendo en cuenta que los nitratos son la principal forma de absorción de nitrógeno por las plantas, estos tienen mucha movilidad en los suelos y se pueden perder fácilmente por escorrentía si no se cuenta con mecanismos de absorción que los retengan (Erickson et al., 2001), lo que podría explicar los resultados obtenidos.
Relaciones entre los mecanismos de los HMA, la estabilidad de agregados y los parámetros físicos y químicos del suelo.
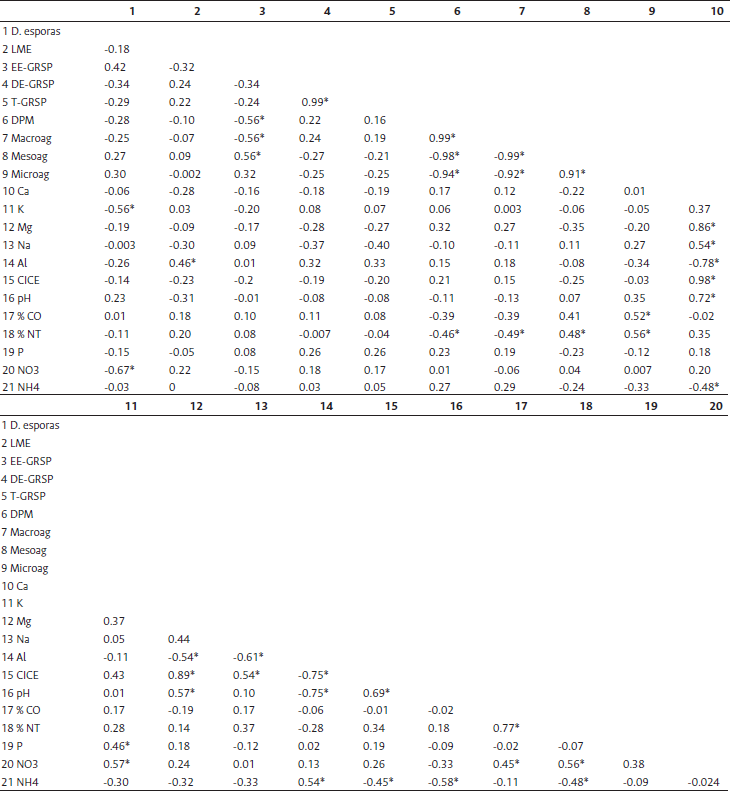
Diferentes estudios han mostrado que los HMA, al igual que otras micorrizas, son agentes importantes en la absorción de nutrientes por las plantas, al incrementar la superficie de contacto de sus raíces, y contribuyen en la estructuración física del suelo formando agregados estables (Cardoso y Kuyper, 2006; Rillig y Mummey, 2006; Delavaux et al., 2017). En el análisis de correlaciones Pearson, la glomalina fácilmente extractable (EE-GRSP) presentó una relación negativa estadísticamente significativa con el índice de estabilidad de agregados DPM y el porcentaje de macroagregados, y una relación positiva con el porcentaje de mesoagregados (Tabla 3). La mayor longitud de micelio externo (LME) y de dos variables glomalina (T-GRSP, DE-GRSP) en los suelos de bosque y rastrojo puede relacionarse con los altos índices de agregación, ya que esta proteína secretada durante el desarrollo de las hifas aglutina las partículas de suelo en forma de cadenas de pequeñas bolsas pegajosas (Stíckíng string bags) (Cardoso y Kuyper, 2006; Rillig y Steinberg, 2002). Por las características insolubles de la glomalina, la materia orgánica contenida atrapada en los agregados queda protegida del efecto erosivo y de la degradación bacteriana (Cardoso y Kuyper, 2006). Los HMA juegan un papel importante en la estructura del suelo y en el desempeño de las plantas. Con el incremento del micelio externo se incrementa la superficie de absorción de nutrientes y, a su vez, como efecto simbiótico, el aumento del flujo de carbono de la planta a los HMA puede también explicar el incremento de la longitud del micelio externo (Delavaux et al., 2017), es decir, es un sistema que se retroalimenta, y también es modulado por la actividad de micro y macroorganismos del suelo (Lavelle et al., 2020).
Tabla 3 Matriz de correlaciones de variables evaluadas. Los valores de coeficiente con asterisco son estadísticamente significativos
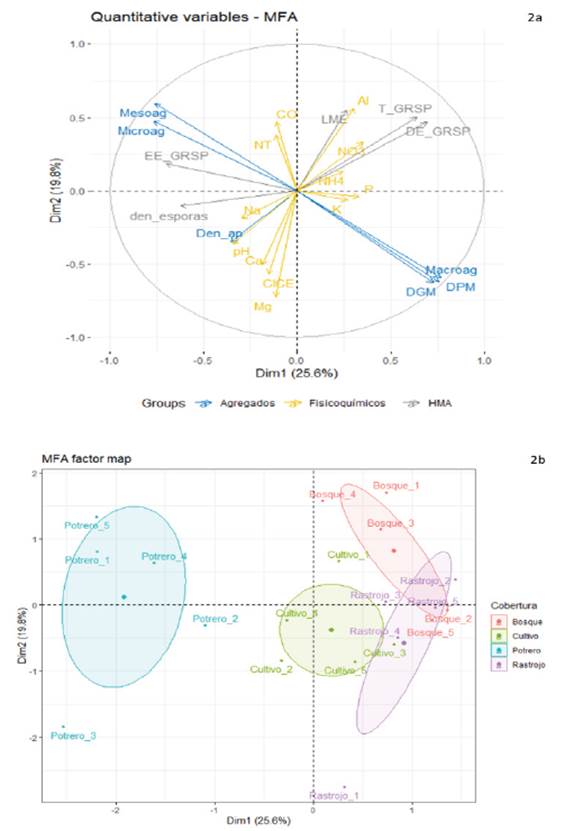
En el análisis factorial múltiple (Figura 2) las componentes uno y dos explicaron el 26 % y 20 % de la varianza total, respectivamente. En el componente uno se destaca la importancia de los HMA en la agregación (Figuras 2a y 2b), mientras que en el componente 2 se representaron mejor variables fisicoquímicas y de los agregados. Con relación a los microagregados del suelo, no se observó una relación directa asociada a los rasgos de los HMA. Este resultado (Figura 2) es congruente con la función de los HMA, que se reconoce más por la formación de los macroagregados (las hifas y la glomalina como agente aglutinante: Rillig y Mummey, 2006; Ji et al., 2019), mientras que la formación y estabilización de los microagregados está relacionada más estrechamente con las comunidades de bacterias y el carbono orgánico del suelo (Six et al., 2004). Este resultado muestra una razonable pero limitada relación entre las características de las HMA y la formación de agregados.
Conclusiones
Los bosques con mayor agregación en términos de macroagregados presentaron también mayor longitud de micelio, glomalina difícilmente extractable (DE-GRSP) y glomalina total (T-GRSP), mientras que los potreros, que tuvieron mayor fracción de micro y mesoagregados, presentaron mayor densidad de esporas y EE-GRSP, lo que sugiere que los HMA contribuyen a mejorar las condiciones del suelo durante la aglutinación de partículas en macroagregados estables por la glomalina. Los suelos con escasa cobertura vegetal tuvieron la tendencia a presentar valores bajos en los índices de agregación, y están relacionados con una baja presencia de rasgos de HMA, lo que limita la contribución al mantenimiento de la estabilidad del suelo. Este, por tanto, puede perderse fácilmente por lixiviación y procesos erosivos que arrastran las partículas del suelo en potreros. Concluimos que el efecto de las micorrizas en la agregación puede afectarse por el tipo de actividad agropecuaria, la intensidad y el laboreo del suelo.

