











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
In Colombia, there are about 175,000 hectares cultivated with cacao trees that produce 60 535 t of cocoa annually (Portafolio, 2017). Losses caused by fungal diseases, especially by Monilia (Moniliophthora roreri [Cif.]), are over 40 % (Rodríguez, 2006; Tirado-Gallego et al., 2016). Furthermore, the black pod rot (BPR) of cacao (Theobroma cacao L.), caused by several species of Phytophthora, is one of the most limiting diseases for the production of cocoa and is present in all the cacao-producing regions worldwide (Cilas & Despreaux, 2004). Different species of this oomycete have been reported to cause significant losses, e.g. Phytophthora capsici Leonian can cause yield losses of up to 10 %, Phytophthora palmivora Butler can be responsible for up to 30 % of the losses (Tahi et al., 2006), while Phytophthora megakarya Brasier and Griffin can cause up to 80 % of the losses (Mfegue, 2012). Therefore, P. megakarya, which is endemic to Africa, is considered the most aggressive Phytophthora species that causes BPR on cacao plants throughout the world (Tahi et al., 2006). P. palmivora is currently present in all continents where cacao is produced, contrary to P. capsici that has only been found in the Americas (Bowers et al., 2001). These two species can attack many other tropical plants; however, the only known host for P. megakarya is cacao. In Colombia, only P. palmivora has been reported as the causal agent of BPR disease in cacao (Rodríguez & Vera, 2015).
Promotion of cocoa crops in the country started in the year 2000, leading to an increase of newly planted areas at altitudes above 800 meters above the sea level (m.a.s.l.) where lower temperatures prevail, favoring the development of the BPR disease. With these favorable edaphoclimatic conditions, BPR exceeds in importance and economic losses diseases caused by Monilia, and positions P. palmivora as the main pathogen for cocoa production in these areas (Rodriguez & Vera, 2015). This situation, together with an increase in the use of the cacao genotype CCN51, which is highly susceptible to BPR when establishing new plantations, increased production losses up to 25 % caused by P. palmivora, while Monilia (Moniliophthora roreli [Cif.]) only caused up to 4.2 % of the losses (Ramírez, 2016; Rodríguez & Vera, 2015).
Traditionally, BPR management strategies have been based mainly on cultural practices, such as phytosanitary pruning, and early elimination of diseased fruits. These are efficient and potential control methods that seem to be less expensive and more environmentally sustainable compared to the application of chemical products (Djocgoue et al., 2006; Ndoumbe-Nkeng et al., 2004). Although the use of copper-based fungicides has been effective in reducing the level of infection by Phytophthora, the high costs of these products make them in many cases inaccessible for small farmers, who produce more than 50 % of the cocoa around the world (de Oliveira & Newman, 2005). Furthermore, biological control is another alternative explored (Deberdt et al., 2008).
Due to the considerable decrease in crop yield caused by the attack of different species of Phytophthora, genetic resistance is considered the most economical and effective alternative for the control of Phytophthora sp. (Surujdeo-Maharaj, Umaharan, & Iwaro, 2001). The programs of genetic improvement of cocoa are commonly oriented to get cultivars resistant to diseases that allow better crop quality and productivity (Dias, 2001; Phillips-Mora, Arciniegas-Leal, Mata-Quirós, & Motomayor-Arias, 2012). Different methods for assessing the degree of genetic resistance in cocoa planting materials include observation of yield in the field under natural infection conditions; likewise, artificial inoculation in fruits and leaves has been tested and adopted (Blaha, 1974; Iwaro & Sreenivasan, 1997; Nyassé, Cilas, Herail, & Blaha, 1995; Saul-Maora, Namaliu, Cilas, & Blaha, 2003).
The reliability of artificial inoculation tests depends on their correlation with the resistance rates observed in the field (Amponsah & Asare-Nyako, 1973; Nyassé, 1997). Therefore, a significant and positive correlation between the results of artificial inoculation of fruits and the total incidence of BPR in the field (including ripe and immature fruits) indicates that artificial inoculation of attached and detached fruits is a reliable method to evaluate the resistance to the BPR disease (Pokou et al., 2008).
However, correlations with field resistance have not always been significant, suggesting the influence of uncontrolled environmental factors affecting field observations or results in screening tests (Efombagn, Bieysse, Nyassé, & Eskes, 2011). These authors concluded that, if leaf disc and detached pod tests are applied under standardized conditions, these can be of great value to speed up the selection of cacao clones resistant to BPR.
Studies on the genetic control of cocoa resistance to three species of Phytophthora (P. palmivora, P. megakarya, and P. capsici) have shown quantitative trait loci (QTL) located in many genomic regions (Flament et al., 2001; Lanaud et al., 2009), with some of these being common to the three Phytophthora species. This situation offers the possibility of improving resistance in cocoa by a possible accumulation of many different resistance genes located in different chromosome regions using marker-assisted selection. Nyadanu et al. (2012) reported that insoluble sugar, flavonoid, tannin, and lignin were the most reliable biochemical factors that can be used as marker traits to screen and select to improve the resistance to the BPR disease of cocoa. The expression of resistance in the host is observed as a decrease in the rate of disease development since one or more stages are delayed or completely blocked. Each resistance factor that plays a role in a step of the life cycle of the pathogen may be considered as a horizontal resistance component (Vanderplank, 1966).
According to what was mentioned above, the aim of this study was to establish the response of detached fruits from six cocoa clones to the artificial inoculation with five isolates of P. palmivora, and to discuss the degree of resistance in the materials evaluated.
Materials and methods
Two artificial inoculation experiments on cacao fruits were carried out in the Laboratory of Phytopathology at the Nataima Research Center (C.I. Nataima) of the Colombian Agriculture Research Corporation (AGROSAVIA), located in the municipality of Espinal, Tolima, Colombia. The center is located in the following geographic coordinates: 40° 12’ Latitude N, and 74° 56’ Longitude W, at an altitude of 430 m.a.s.l. The laboratory had an average temperature of 22 °C and a relative humidity of 70 %.
Phytophthora palmivora isolates
Five isolates of P. palmivora were selected from the C.I. Nataima collection, obtained from different cacao producing regions of Colombia (Table 1). The isolates used were selected for showing the highest levels of aggressiveness in screening tests, previously performed on detached fruits of the clone IMC67.
Table 1 Phytophthora palmivora isolates collected from different cacao producing regions and materials of Colombia in 2014.
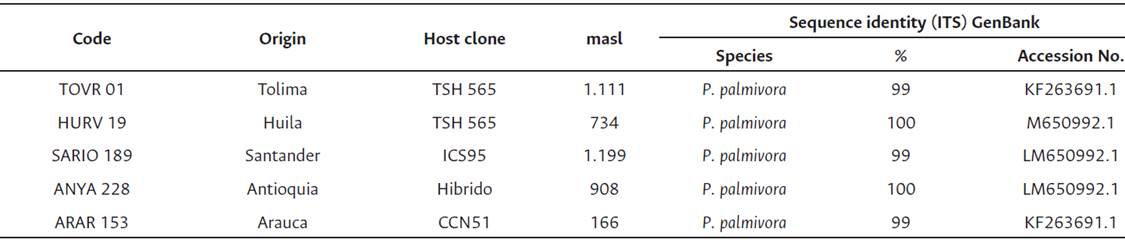
Note. [a ] masl: meters above sea level
Isolates were previously characterized by colony morphology (texture, aerial development of mycelia and color), and biometric characteristics of sporangia and chlamydospores (length, width, and length/width ratio of sporangia, diameter of chlamydospores, length of the pedicel and presence or absence of papilla and chlamydospores, and sporangia dehiscence). The identification of the isolates was supported using molecular markers (ITS4, ITS6, and COX). Isolates were conserved in Eppendorf tubes with sterile distilled water at 10° C. These were reactivated on cacao fruits of the cacao clone IMC67 to minimize any epigenetic influence on the virulence of the pathogen. Besides, the environment in which the experiments were conducted was maintained constant with a relative humidity of 90 % and a temperature of 28° C to standardize the environmental influence on the virulence of the pathogen.
Plant material
Healthy fruits were obtained from commercial cacao plantations located in Palocabildo, department of Tolima, Colombia, where two-month-old fruits were bagged to avoid infection in the field. Once fruits reached an age of 4.5 months, they were cut and taken to the laboratory for artificial inoculation with P. palmivora. Cacao fruits of the clones CCN51, ICS95, EET8, TSH565, PA46, and IMC67 were used. The cacao clones selected for this study represent the two major types of cacao planted in most cacao- growing regions of Colombia, i.e., PA46 (cluster marañon-PA series) and IMC67 (cluster Iquitos-IMC series) (Motamayor et al., 2008), and Trinitarian (CCN51, ICS95, EET8, and TSH565). Clone PA46 was used as a resistant control (R) and clone CCN51 as a susceptible control (S) (Iwaro, Bekele, & Butler, 2003; Phillips-Mora et al., 2012).
Inoculation of fruits and qualification of resistance reaction
Ten fruits of approximately 4.5 months old from each of the clones selected were inoculated with a suspension of 1.5 x 105 zoospores mL-1 from Petri dishes with 12-day-old cultures with each of the isolates selected, impregnated in two discs of filter paper of 0.5 cm diameter, and placed at equidistant points in the equatorial zone of the cacao pod. Fruits inoculated with sterile distilled water corresponded to the absolute control treatment (Phillips-Mora et al., 2012; Rodríguez & Vera, 2015). Subsequently, the fruits were incubated individually in a humid chamber (airtight plastic bag with a paper towel moistened with sterile distilled water) at 28 °C. Subsequently, six and ten days after inoculation (DAI), the incidence, i.e., the presence or absence of injuries, and the severity, measured as the average lesion diameter (ALD) obtained from two perpendicular sides of the pod, were evaluated. With the ALD, the response to the infection in each clone was made, and the degree of genetic resistance was graded according to the scales published by Phillips-Mora & Galindo (1989) (Table 2). The level of aggressiveness of the isolates was also established according to the diameter of the lesion in the clones evaluated.
Table 2 Reaction scale to BPR disease based on mean diameter of lesion (cm) caused by P. palmivora in cacao fruits (Phillips-Mora & Galindo, 1989).
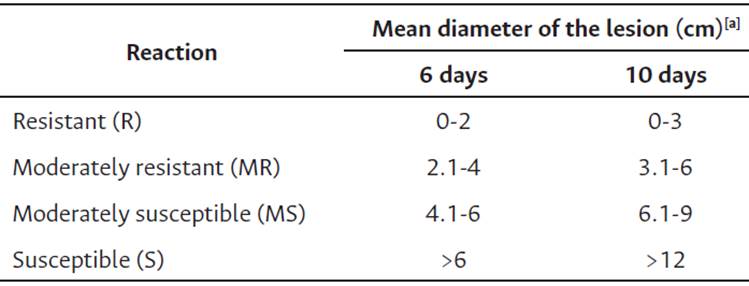
Note. [a ]Mean diameter of cacao pod lesion calculated from length and width measurements.
Expression and development period of BPR symptoms in cacao
The periods of symptom expression of P. palmivora were identified in an artificial inoculation experiment established in the field of an experimental plot in La Isla (C.I. La Suiza, department of Santander, Colombia). Growing cacao fruits of approximately 1.5 months selected from the clones ICS39, ICS40, ICS60, and ICS95 were bagged in order to guarantee their health until they reached an age of approximately 4.5 months. Subsequently, 10 fruits of each of the clones were artificially inoculated with a suspension of 1.5 x 105 zoospores mL-1, as described above. The absolute control consisted of inoculation with sterile distilled water. Observations were made every 24 hours from the moment inoculation was performed until the time of symptom onset was recorded. Then, these were categorized according to their development as necrotic points (first symptoms), evident spots (> 2 cm in diameter), presence of mycelium (white mycelium), and sporulation (formation of sporangia). In this trial, the P. palmivora isolate SARIO189 was used due to its high level of aggressiveness that was previously established.
Statistical design and analysis
Fruit inoculation and symptom observation: Two independent experiments were carried out in a completely randomized design (CRD) with six clones and five isolates to evaluate the average lesion diameter (ALD), with ten repetitions per treatment. Data from ADL of the two experiments were combined after establishing the homogeneity of the variance using Cochran’s Q test (Gomez & Gomez, 1984), the normality of the residuals employing Shapiro-Wilk’s test, and the homogeneity of the variances using Levene’s test. ALD data were subjected to a two-way variance analysis with clone and isolate as factors, to assess the significance of the treatment effect and the interaction between treatments; finally, the means were compared through the t-test (p < 0.05) using the software Infostat (di Rienzo et al., 2012).
Periods of expression and development of BPR symptoms: This experiment was carried out using a completely randomized design with ten repetitions per treatment. The data were assessed using an analysis of variance (ANOVA). The normality of the residuals and the homogeneity of the variances were verified employing the tests Shapiro-Wilk and Levene’s, respectively. ALD data were subjected to an analysis of variance, and the treatment means were compared through the t-test (p < 0.05) using the software InfoStat (di Rienzo et al., 2012).
Results
Fruit inoculation, resistance evaluation and virulence reaction of isolates
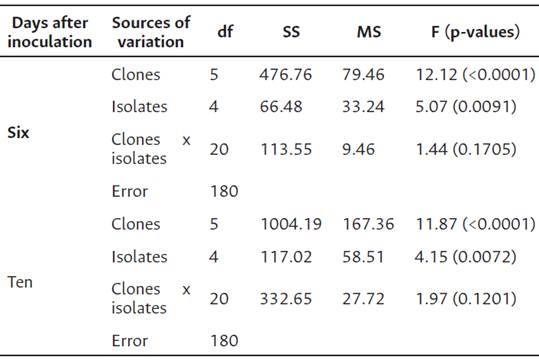
The analysis of variance of ADL was statistically significant (p < 0.001), influenced by the cacao genotypes and the P. palmivora isolates. Host genotype x isolate interaction effects were, however, not significant (p < 0.005) (Table 3).
Table 3 Analysis of variance of severity levels in six cocoa clones (CCN 51, ICS 95, EET 8, TSH 565, IMC 67 and PA 46), six and ten days after inoculation (DAI) with five isolates (ANYA 228, SARIO 189, ARAR 153, TOVR 01 and HURV 19) of Phytophthora palmivora.
The results of artificial inoculation in detached cacao pods showed significant differences (p < 0.05) in the lesion size for the clones evaluated at 6 DAI (Table 4). Clone CCN51 showed the highest significant value in lesion size and was superior in 46.08 % and 46.39 % compared to the clones IMC 67 and PA 46, respectively. Clone CCN51 was classified as susceptible (S), and clones PA 46 and IMC 67 were classified as moderately susceptible (MS). Clones ICS95, EET8, and TSH565 did not show significant differences between them and were classified as susceptible (S) according to their response to infection. In all clones, incidence reached a value of 100 % (Table 4).
Table 4 Reaction to BPR disease in detached fruits of six cocoa clones, six and ten days after artificial inoculation to Phytophthora palmivora (isolates ANYA 228, SARIO 189, ARAR 153, TOVRO1, HURV19 ). C.I. Nataima, Espinal, Tolima, Colombia.
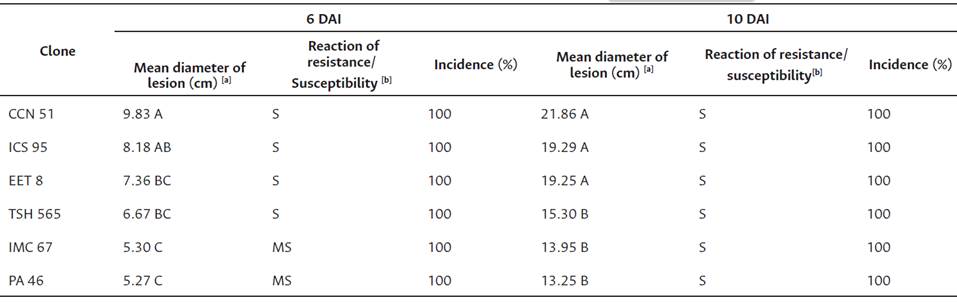
Note. [a ] Different letters in the same column represent significant differences identified by the Tukey test (p < 0.05). [b ] S: susceptible, MS: moderately susceptible.
The results of the analysis of variance of artificial inoculation in detached cacao pods indicated significant statistical differences (p < 0.05) in the size of the lesión for clones evaluated at 10 DAI (Table 4). Clones CCN51, ICS95, and EET8 showed the highest values with 21.86 cm, 19.29 cm, and 19.25 cm, respectively, and were statistically similar between them (p < 0.05). The lowest values for lesion size corresponded to the clones TSH565, IMC67, and PA46, with average values of 15.30 cm, 13.95 cm, and 13.25 cm, respectively. All clones tested reached values higher than 12 cm in lesion size, so they were classified as susceptible (S) (Phillips-Mora et al., 2012).
Aggressiveness of P. palmivora isolates
The aggressiveness variability of P. palmivora isolates was established as the average size of the lesion measured in two perpendicular directions of the cacao pod in five clones evaluated at six DAI. Significant differences (p < 0.05) were found among the five isolates used (Table 5). The most aggressive isolate corresponded to HURV19 with a value of 13.79 cm and was 45.98 % higher compared to ANYA228, which exhibited the lowest aggressiveness amongst all isolates evaluated. Isolates SARIO189 and ARAR153 did not show significant differences between them, with lesion size values of 10.84 and 12.11 cm, respectively, showing an intermediate behavior in aggressiveness (Table 5).
Table 5 Virulencie of selected isolates of P. palmivora in detached fruits of five cocoa clones (CCN51, ICS95, EET8, TSH 565, IMC 67, PA 46). C.I. Nataima, Espinal, Tolima, Colombia.
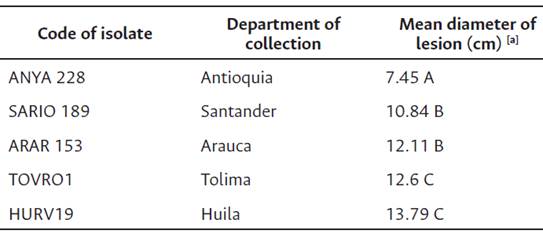
Note. [a ] Different letters in the same column represent significant differences identified by the Tukey test (p < 0.05). Means were calculated from 10 independent measurements for each pod in five clones, six days after inoculation (DAI).
Periods of expression and development of P. palmivora symptoms
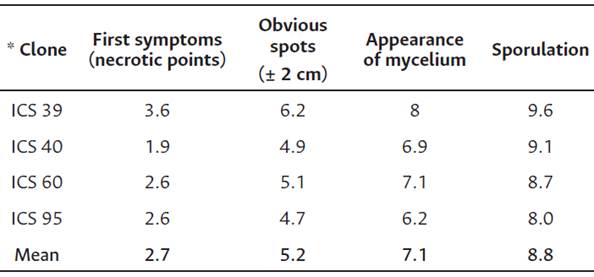
Symptom development indicated that, on average, the clones inoculated showed the first symptoms (necrotic points) in the inoculation zone at 2.7 DAI. Necrotic spots appeared at 5.2 DAI and increased in size as the infection process in the cortex tissue progressed until the appearance of mycelia at 7.1 DAI. Finally, the formation of sporangia occurred at 8.8 DAI (Table 6). Clone ICS95 had the pathogen isolate with the shortest periods of symptom expression: on average, it lasted 8 days until sporulation. The opposite behavior was observed in clone ICS39, where P. palmivora required 9.6 DAI for the formation of reproductive structures. In all inoculated clones, the incidence of P. palmivora was 100 % at 6 DAI.
Discussion
Disease symptoms were observed in cacao pods of all genotypes inoculated with P. palmivora at 6 DAI, showing that none of them was immune to the pathogen. This indicates the existence of variation in the reaction of the evaluated plant material, suggesting that, in this pathosystem, there is a resistance gradient, quantitative resistance, or that it is a non-specific race, as has been indicated in previous studies (Blaha, 1974; Legavre et al., 2015; Surujdeo-Maharaj et al., 2001). This type of resistance is controlled by many genes that interact delaying the development of the pathogen, and its multigene character makes it more challenging to overcome (Eulgem, 2005).
This study showed a different response to infection in the clones, which can provide evidence of the existence of quantitative resistance or a non- specific race of P. palmivora in cocoa. Symptoms of the disease were observed in all genotypes inoculated with each of the five isolates of P. palmivora collected in geographically distant cacao-producing regions in the country. These experiments indicate that resistance is expressed with similar patterns, although with different values when pods are inoculated with each of the five isolates of P. palmivora.
The lesion size values obtained showed significant differences (p < 0.05), confirming the differentiated response of each cacao genotype to infection by P. palmivora (Table 3). A broad genetic variation resistant to BPR has been reported in cacao by Iwaro et al.(1997), Nyadanu et al. (2010), and Tahi et al. (2006). Previous studies indicated that six days of incubation are not enough to accurately separate resistance from susceptible genotypes, whereas, after ten days, the lesion size (LS) observed in each fruit tends to stabilize (Phillips-Mora & Galindo, 1989).
The melon-shaped genotype PA46 showed the smallest lesion size, and this agrees with the fact that resistance to BPR has been found predominantly associated with high Amazonian Forastero genotypes (Bartley, 2005; Kébé et al. 1999). Among these, SCA6, PA150, and P7 have shown lower susceptibility to the pathogen, and some lower Amazonian Forasteros (amelonado or melon-shaped) are included in this group. In general, Trinitarian materials are considered more susceptible to the BPR disease (Paulin et al., 2008), as it was confirmed with the genotypes evaluated in this study, with the Trinitarian genotype CCN51 exhibiting the highest lesion size values, followed by ICS95, EET8, and TSH565 (Table 4). This observation agrees with previous research reports, where this material has been classified as highly susceptible in several regions of Colombia (Phillips-Mora & Galindo, 1989; Rodríguez & Vera, 2015; Ramirez, 2016). Iwaro & Sreenivasan (1997) estimated that 90 % of the known commercial cacao genotypes are susceptible to this disease. In fact, clone CCN51 may be used as a control in tests that involve the evaluation of resistance to black pod rot in cacao genotypes (Lisboa, Cerqueira-Silva, Clement, & Newman, 2011).
As mentioned, genotype ICS95 was the most susceptible after CCN51. Moreover, the values obtained coincided with the observation of the life cycle of P. palmivora in the field, where this genotype showed the lowest duration value for the infection stages, with 1.6 days less compared to genotype ICS39, which showed the most extended infection period.
A decrease in the life cycle of the pathogen may indicate the expression of some level of resistance, as one or more stages are delayed or completely blocked. Each of several factors that reflect specific affectation of a step in the life cycle of a pathogen may be considered as a component of horizontal resistance (Vanderplank, 1966). Horizontal or quantitative resistance is based on four fundamental actions: resistance to penetration, resistance to pathogen growth, resistance to sporulation, and resistance to lesion growth (Vander Plank, 1982).
This study identified a significant variation (p < 0.05) in the aggressiveness of P. palmivora isolates in Colombia, with a difference of 45.98 % in the ALD between the more aggressive (HURV19) and the less aggressive (ANYA228) isolates. Under semi-controlled experimental conditions in which this study was conducted, this variation may be predominantly attributed to genetic causes. According to Agrios (2004), the development of the disease is a function of the host, the pathogen, and the environment. In the current study, the selection of fruits was performed in the same development stage, and plants were grown under similar environmental conditions, minimizing the influence of environmental factors. The fact that there was no interaction between the isolate and the cacao genotype is vital in the generation of quantitative resistance to P. palmivora. In this sense, this characteristic allows the identification of different levels of resistance in cacao genotypes using any isolate. However, considering that pathogen population may be highly variable in aggressiveness, as observed in this work, Lessa et al. (2020) emphatically use a mixture of isolates that can reflect the variability of the population and, therefore, the identification of more significant levels of resistance against this pathogen.
Furthermore, if the resistance level present in a genotype is consistent in all the evaluations with several isolates in different regions of the world, the varieties created with this resistance to P. palmivora in one location would also express resistance in another locations (Surujdeo-Maharaj et al., 2001). Indeed, some similarities have been observed in the classification of cacao genotypes regarding the resistance to P. palmivora and P. megakarya in Cameroon, Ivory Coast, and Togo; where the Trinitario parents were generally more susceptible to the disease, lower Amazon (LA) and some upper Amazon (UA) amelonado type parents, such as Sca 6, P 7, Pa 150 or T85/799, should help develop less susceptible cultivars (Nyassé et al., 2007). A significant difference (p < 0.05) was also observed among clones for pod resistance to pathogen invasion based on lesion size. Relatively smaller lesions were recorded for SCA 6 and ICS 95. Lesion sizes for ICS 6, ICS 40, ICS 84, SCA 12, and PA 46 were moderate, whereas relatively large lesions were produced on other clones (Iwaro & Sreenivasan, 1997).
Development and refining of the artificial inoculation method of detached cacao fruits for the identification and characterization of the level of resistance to black pod rot has shown respectability and reliability, finding a positive correlation between the data obtained with the test on detached pods and natural infection in the field (Iwaro & Sreenivasan, 1997; Kébé et al., 1999). Early resistance detection methods using detached leaves and pod segments have correlated well with observations of incidence and severity of BPR disease in the field, so they are currently used to evaluate segregating progenies to detect and discard highly susceptible materials (Iwaro & Sreenivasan, 1997; Iwaro, Thévenin, Butler, & Eskes, 2005).
The use of these methods allows a rapid and early evaluation of resistance levels of a large number of materials. The percentage of mature fruits infected in the field is determined not only by the resistance of the fruit to the infection, but also by escape phenomena, such as the fructification cycle (production outside the main epidemic period), and fructification intensity (Cilas & Despréaux, 2004). The fructification cycle usually explains about 43 % of the variation in infection level (Kébé et al., 1999), and the use of an artificial inoculation technique in detached fruits may allow reducing this difficulty and reflect variation in the genetic resistance of cacao tissue to infection, as the main parameter.
The existence of polygenic or quantitative resistance to different species of Phytophthora in cacao indicates that this can be used for genetic improvement by recurrent selection processes (Nyassé et al., 2007), where the search for resistance to Phytophthora sp. should be expanded. This expansion should include selected genotypes in regions with high disease pressure and early detection of resistant progenies using artificial inoculation methods. Recent molecular studies on the genetic regulation of resistance in Theobroma cacao to three species of Phytophthora (P. palmivora, P. megakarya, and P. capsici) have shown that QTL are located in many genomic regions (Flament et al., 2001; Lanaud et al., 2009), and some of them are common to all three Phytophthora species.
This situation offers the possibility of improving resistance to Phytophthora in cacao through a possible accumulation of many diverse resistance genes located in different chromosomal regions through the marker-assisted selection and rapid selection methods (Legavre et al., 2015).
Conclusions
The use of a method for early detection of resistance allowed identifying PA 46 as a tolerant clone to the BPR disease; this suggests the possibility of selecting genotypes with durable resistance to the pathogen, allowing the use of these materials in breeding programs in search of resistance to Phytophthora sp.

