











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
En Colombia, la producción de flores en el año 2019 alcanzó las 243 548 t; Cundinamarca y Antioquia fueron los departamentos de mayor participación, lo que generó un empleo rural para 193 764 personas y ubicó al país como el segundo exportador de flores del mundo (MADR, 2020), con casi 1500 variedades comerciales cultivadas, de las cuales la especie más producida es la rosa debido al gran potencial en el mercado internacional (Albornoz et al., 2016).
El aumento en el rendimiento requiere de la implementación de nuevas tecnologías para compensar el costo de producción. Dentro de estas se destaca la siembra de flores de corte en hidroponía bajo condiciones de invernadero, ya que tienen una mayor productividad (Castellanos, 2010). Sin embargo, la implementación de los cultivos de flores en sustratos ha provocado la reducción de la capacidad buffer que tiene normalmente el suelo, lo cual disminuye la asimilación de nutrientes minerales y genera menores producciones (Argo y Fisher, 2003).
Una planta con un suministro adecuado de nutrimentos tiene un óptimo crecimiento, desarrollo, calidad y rendimiento (de Mello, 2021). En ese sentido, el Si es uno de los elementos que desempeña un papel importante en la obtención de altas productividades, y es considerado un elemento benéfico para las plantas con funciones, como proteger cultivos, brindar resistencia a diferentes tipos de estrés biótico y abiótico (Wiese et al., 2007), mejorar la síntesis de metabolitos secundarios antifúngicos, como las fitoalexinas, y reducir el daño oxidativo celular a causa de la toxicidad con metales pesados o un estrés por sequía (Bhat et al., 2019).
El Si puede mejorar el crecimiento y rendimiento de los cultivos al incrementar la disponibilidad de los nutrimentos bajo condiciones de campo y la distribución de estos dentro de la planta, al controlar la absorción de metales en la raíz (Wiese et al., 2007; Hernández-Apaolaza et al., 2020; Pavlovic et al., 2021). Se ha demostrado que la absorción de Si afecta los niveles de macro y micronutrientes dentro de la planta (Greger et al., 2018), por lo que la fertilización con silicio estaría directamente relacionada en la calidad nutricional, aumentado la productividad de los cultivos bajo una agricultura sostenible.
La interacción del Si con los demás nutrimentos aún no se ha esclarecido del todo. Wiese et al. (2007) plantean que los métodos en que la planta toma el Si, sea vía foliar o radicular, pueden afectar la concentración de este elemento, por ende, es posible que la aplicación del Si modifique la absorción de nutrientes de manera diferente en especies de interés comercial, como la rosa sembrada en sustratos. Algunos antecedentes en la aplicación del silicio en el cultivo de rosa han demostrado resultados como hojas más fuertes, gruesas y verdes, mayor longitud de los tallos y un incremento en la resistencia natural al mildiu polvoso (Sphaerotheca pannosa) y a Botrytis cinerea (Albornoz et al., 2016).
Existen pocos estudios que muestren los efectos de diferentes dosis de Si sobre el nivel y la interacción con otros nutrientes minerales en el cultivo de rosa en sustratos. Por lo anterior, el objetivo de esta investigación fue evaluar el efecto de diferentes métodos de aplicación de silicio en la concentración foliar de nutrientes en rosa hidropónica.
Materiales y métodos
Localización
El experimento se llevó a cabo en la finca comercial Flores las Acacias, en Madrid, Cundinamarca (Colombia), ubicada a 2557 m s.n.m., y presenta una temperatura máxima promedio anual de 19° C, y una mínima de 8° C, precipitación promedio anual de 625 mm, con brillo solar promedio de 4.5 horas por día y humedades relativas del 80 %.
Material vegetal
Se utilizaron rosas tipo spray de la variedad “Snowflake” injertadas sobre el patrón “Nathal Briar”, sembradas en camas hidropónicas con sustrato de cascarilla de arroz quemada, el cual no contiene minerales solubles que puedan ser asimilados por la planta. Se sembraron 40 tallos por unidad experimental dispuestas a doble surco, con una distancia entre plantas de 0.20 m y entre surcos de 0.35 m. Las parcelas tenían unas distancias entre ellas de 1 m, con un área de 3 m2 cada una.
Diseño experimental
Se realizó un diseño completamente al azar con cuatro tratamientos, los cuales consistieron en la aplicación de silicio al sustrato (fertirriego, vía foliar, sustrato + foliar y un testigo). Cada tratamiento contó con cuatro repeticiones para un total de 16 unidades experimentales (UE) y cada UE correspondió a una cama con 40 tallos injertados, para un total de 640 plantas.
El manejo del cultivo se llevó a cabo con la instalación del fertirriego por goteo (pH = 6.1), en el cual se suministraron todos los macro y micronutrientes para las plantas de rosa, como son nitrógeno (N), fósforo (P), potasio (K+), calcio (Ca2+), magnesio (Mg2+), azufre (S), hierro (Fe), zinc (Zn), boro (B), manganeso (Mn) y cobre (Cu). Las aplicaciones de Si se realizaron justo después del pinch (corte de yema apical para eliminar dominancia y permitir el desarrollo de tallos laterales) con un fertilizante a base de Si y K (Tabla 1). Para el tratamiento de fertirriego se utilizó una dosis de 0.33 ml/l. En el tratamiento vía foliar, se asperjó una dosis de 1.5 ml/l del mismo compuesto. Para la aplicación de Si al sustrato y vía foliar se manejaron las mismas concentraciones mencionadas anteriormente. Las aplicaciones se realizaron a los 7, 14, 28, 42, 56 y 70 días después del pinch (ddp).

Variables de respuesta
La longitud de tallo floral se midió durante 14 semanas después del pinch y se ajustó a un modelo logístico doble sigmoide, el cual es uno de los modelos que mejor describe el crecimiento de las variables biológicas (Álvarez- Herrera et al., 2017). Este modelo se determinó mediante la ecuación (1):
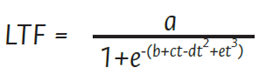 Ec. 1
Ec. 1
donde LTF es la longitud de tallo en cm según el tiempo (t); a es el valor máximo de la variable en el tiempo en cm; b, c, d y e son constantes relacionadas con la velocidad relativa de crecimiento (VRC) y los puntos de inflexión de la curva de crecimiento. La VRC se calculó con la ecuación (2) , al derivar la fórmula de LTF.
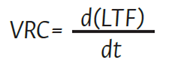 Ec. 2
Ec. 2
La concentración foliar de N, P, K, Si, Ca2+, Mg2+, S, Na, Fe, Zn, B, Mn y Cu se midió en el laboratorio AGRILAB® con muestras foliares de rosa tomadas en la etapa fenológica “rayando color” (83 ddp). La técnica para cuantificar los elementos fue la de espectroscopía de absorción atómica, a excepción del P, S y B, los cuales se evaluaron mediante el método colorimétrico, acorde con Gómez (2013).
Análisis estadístico
El manejo estadístico se realizó con el programa SAS v.9.2 (SAS Institute Inc, Cary, NC). Luego de la prueba de normalidad de Shapiro- Wilk, se realizó un análisis de varianza (ANOVA) con el procedimiento GLM y se clasificaron los tratamientos mediante la prueba de Duncan (P<0.05).
Resultados y discusión
Longitud del tallo floral
La LTF no presentó diferencias significativas entre los tratamientos de Si, por lo que, con los promedios, se generó una curva de crecimiento en la que el modelo logístico doble sigmoideo fue el que mayor coeficiente de correlación presentó (R2= 0.996), y se aprecia en la Figura 1 y en la ecuación (3), lo que significa que este modelo es el que mejor describe el crecimiento de la LTF para el cultivo de rosa de variedad “Snowflake” en las La LTF no presentó diferencias significativas entre los tratamientos de Si, por lo que, con los promedios, se generó una curva de crecimiento en la que el modelo logístico doble sigmoideo fue el que mayor coeficiente de correlación presentó (R2= 0.996), y se aprecia en la Figura 1 y en la ecuación (3), lo que significa que este modelo es el que mejor describe el crecimiento de la LTF para el cultivo de rosa de variedad “Snowflake” en las condiciones de la sabana de Bogotá. El crecimiento en altura se caracterizó por tener cinco fases. La fase I se destacó por un crecimiento lento hasta los 215 GDC (2.96 cm); la fase II por un crecimiento rápido desde los 215 hasta los 558 GDC (47.49 cm); luego se ralentiza el crecimiento de la planta hasta los 721 GDC (51.10 cm), lo que corresponde al final de la fase III; sigue una IV fase donde vuelve a presentarse un ligero incremento desde los 721 hasta 932 GDC (57.83 cm); y finalmente la fase V, en la que se produce el corte del tallo floral.
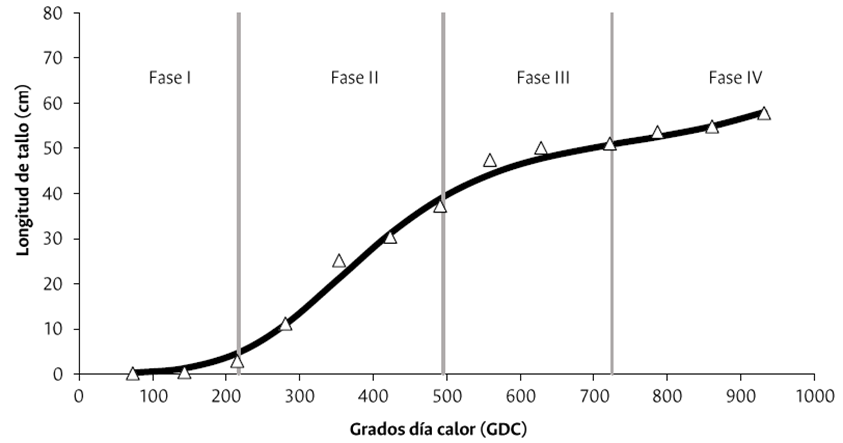
Figura 1 Curva de crecimiento de la longitud del tallo floral de rosa “Snowflake” cultivada en la sabana de Bogotá.
La VRC alcanzó los máximos valores a los 353 GDC (34 ddp) con 0.148 cm día-1 (Figura 2). A partir de este momento, la VRC disminuyó rápidamente hasta los 721 GDC (69 ddp), instante en el que inicia el estadio “garbanzo”, y comienza una etapa de muy bajo crecimiento en la LTF hasta los 787 GDC (estadio fenológico de “rayando color”), punto en el cual nuevamente se acelera el crecimiento en la altura del tallo hasta el punto de corte (923 GDC).
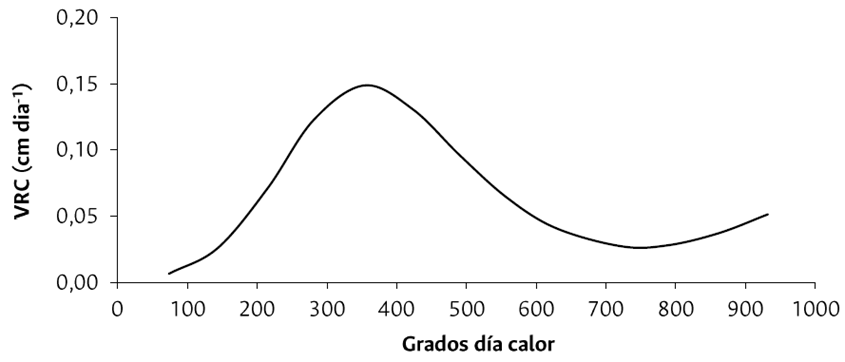
Figura 2 Curva de velocidad relativa de crecimiento de la longitud del tallo floral en rosa “Snowflake” cultivada en la sabana de Bogotá.
En este sentido, el estadio fenológico denominado “palmiche” (558 GDC) consiste en la diferenciación del punto floral (inducción floral) y debido a que la variedad “Snowflake” es una rosa tipo spray, esta genera varios brotes (3 a 6 puntos florales), lo que disminuye el crecimiento de la LTF. A partir de allí inicia el desarrollo del pedúnculo y punto floral, lo que se traduce en una disminución en la VRC hasta llegar al estadio “garbanzo” (721 GDC). Por otro lado, entre el estadio “garbanzo” y “rayando color” (787 GDC), el aumento del LFT se ralentiza considerablemente como consecuencia del incremento en el crecimiento del diámetro de la flor. Finalmente, se produce un leve aumento de la VRC que constituye el factor diferencial en la calidad de las rosas sembradas en la sabana de Bogotá, generando tallos tipo exportación.
Nitrógeno, fósforo y potasio
No hubo diferencias significativas entre las aplicaciones de Si realizadas para el nitrógeno, fósforo y potasio (Tabla 3). El N mostró un valor promedio de 4.24% ± 0.26 de materia seca (MS) para todos los tratamientos a los 83 ddp, mientras que el P y el K mostraron valores de 0.36% ± 0.03 y 2.08% ± 0.09, respectivamente. Estos elementos se encontraron dentro del rango recomendado por Cabrera (2003). Los valores para la concentración de N son superiores a de Pascale y Paradiso (2001), quienes reportan datos de 3.33 % para rosas variedad “Dallas”. En cuanto al fósforo y el potasio, estos autores reportan 0.29 % y 2 %, respectivamente, que también resultan inferiores a los encontrados en el presente estudio. Al respecto, Franco et al. (2013) reportaron valores nutricionales en rosa de variedad “Charlotte” cultivada en la sabana de Bogotá de 3.70 %, 0.29 % y 2.02 %, para N, P y K, respectivamente, los cuales también fueron inferiores a los comparados con este estudio. Ello podría indicar que la variedad “Snowflake” demanda mayores requerimientos nutricionales.
Es difícil que el Si afecte la concentración de los principales macronutrientes en las plantas, ya que esta depende del crecimiento y desarrollo de cada especie. Esto concuerda con Greger et al. (2018), quienes mencionan que la aplicación de Si no aumentó la absorción de N, P y K para las plantas de lechuga, zanahoria, trigo y maíz. Asimismo, se ha reportado que existe un antagonismo entre la interacción de Si y N en arroz (Wu et al., 2017).
En la relación de P y Si, estos resultados no concuerdan con otras investigaciones realizadas, en las que la aplicación de Si aumentó en la raíz la exudación de ácidos orgánicos que movilizan el P y regulan genes transportadores en trigo (Kostic et al., 2017) y en arroz (Ma y Takahashi, 1990). Se ha reportado que existe una relación compleja entre el P, Si y Zn dentro del apoplasto de la hoja, ya que al aplicar dosis elevadas de Zn más Si, se ve reducido el transporte de P en el apoplasto de la hoja en plantas de pepino y cebada (Wiese et al., 2007). Este fenómeno explicaría la mayor acumulación de Zn que tuvo la rosa “Snowflake” en este experimento bajo diferentes métodos de aplicación de Si (Tabla 3).
Por otra parte, la aplicación de Si no mejoró la absorción de K en las hojas. Estudios recientes han demostrado que la aplicación de Si mejoró la absorción de K en varias plantas forrajeras tanto en deficiencia de K como en una nutrición completa (Buchelt et al., 2020). Liang (1999) y Liang et al. (2005) comentan que el Si puede mejorar la absorción de K, ya que el Si está directamente relacionado con la activación enzimática de la H+-ATPasa y H+K+-ATPasa, que son la fuente principal del gradiente de potencial electroquímico H+.
Calcio, magnesio y azufre
La concentración de Ca2+, Mg2+ y S a nivel foliar mostró diferencias significativas entre las distintas aplicaciones de Si realizadas al cultivo de rosa (Tabla 2). Los elementos de Ca2+ y Mg2+ estuvieron dentro del rango normal de concentración foliar recomendada para rosa sembrada en la sabana de Bogotá (Tabla 2). Las plantas que presentaron las mayores concentraciones de Ca estuvieron sometidas a la aplicación foliar de Si y superaron en un 3 % y 14 % al Ca registrado tanto en las plantas con aplicación de Si en el sustrato+foliar como en el testigo, respectivamente (Tabla 2). Estos valores foliares son superiores a los reportados por Cabrera (2002) en rosas “Bridal White” injertadas en el portainjerto Rosa x “Nathal Briar” con un Ca de 1.60 %. Esto indicaría que la aplicación de Si al sustrato+foliar mejora la absorción de Ca2+ en plantas de rosa “Snowflake”. Con respecto a la interacción de Si y Ca2+, se ha reportado que, en plantas de tomate hidropónico, al aplicar K2 SiO3 a la solución nutritiva estándar reportaron Mg de 0.26 % de MS para la variedad de rosa “Charlotte” sembrada en la sabana de Bogotá, lo que indicaría una mayor concentración y absorción de este elemento en la variedad “Snowflake”. Por otro lado, al analizar la interacción del Si y el Mg, Greger et al. (2018) reportaron que la aplicación foliar de Si promueve la absorción y translocación de Mg2+ en brotes y hojas de algunas leguminosas, gramíneas y hortalizas. Sin embargo, otras investigaciones en plantas forrajeras (Buchelt et al., 2020) y en maíz (Hosseini et al., 2019) mencionan que el Si no tiene influencia en la absorción, traslocación y acumulación de Mg2+ en los brotes con o sin déficit de este elemento. Del mismo modo explican que el Si no mejora el Mg dentro de la planta, pero sí favorece el uso eficiente del elemento al mejorar las características fisiológicas de las plantas bajo el estrés inducido de Mg gracias al efecto indirecto del Si en el aumento significativo de los niveles de clorofila y en la regulación del metabolismo de los azúcares y el equilibrio hormonal (Hosseini et al., 2019).
Tabla 2 Concentraciones foliares de macronutrientes en plantas de rosa hidropónica “Snowflake” en el estado fenológico “rayando color” bajo diferentes métodos de aplicación de Si
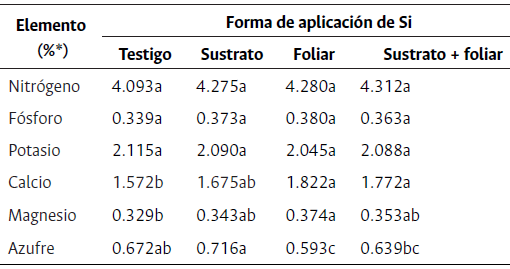
* Expresado en porcentaje de masa seca. Letras distintas en la misma fila indican diferencias estadísticas significativas entre tratamientos según la prueba de Duncan (P≤0.05).
En cuanto al S, las plantas con aplicaciones de Si al sustrato mostraron las mayores concentraciones de S, con un 17.2 % más que las plantas sometidas a la aplicación de Si foliar. Este valor de S es mayor que las concentraciones foliares recomendadas en Rosa spp. por Cabrera (2003), aun así, no se apreciaron síntomas de toxicidad por S en las plantas estudiadas. En comparación con los datos nutricionales analizados por Franco et al. (2013), el rango de S a nivel foliar en rosa cultivada en la sabana de Bogotá oscila entre 0.34 % y 0.44 %, el cual es menor a lo reportado por todos los tratamientos de aplicación de Si evaluados. Al ser el primer reporte de concentraciones nutricionales de rosa variedad “Snowflake” se podría inferir que este cultivar acumula una gran cantidad de S, el cual aumenta con la aplicación de Si. Con respecto a la interacción del Si y el S, los estudios indican que la absorción y la acumulación de S no se vieron afectados por el suministro de Si en pastos forrajeros (Buchelt et al., 2020). Asimismo, otras investigaciones mencionan que la adición de Si en cebada bajo un estrés por déficit de S indujo la expresión del transportador de S (HvST1;1) en la raíz, pero esto no aumentó los niveles de S en los brotes, por el contrario, disminuyó durante el crecimiento (Maillard et al., 2018).
Micronutrientes
La aplicación de Si no aumentó la concentración de Fe ni de Cu en las plantas de rosa. Por el contrario, la aplicación foliar de Si disminuyó estas concentraciones en un 9.44 % y 13.63 %, respectivamente. A pesar de que el Si foliar redujo el Fe y Cu, los valores de estos elementos estuvieron dentro del rango normal recomendado y no alcanzaron valores de deficiencia (Tabla 2).
El Fe en rosa “Snowflake” fue mayor al reportado en las variedades “Bridal White” y “Charlotte” en la etapa floración con 51 y 76 mg kg-1 de MS, respectivamente (Cabrera, 2002; Franco et al., 2013).
En la relación Si y Fe, Greger et al. (2018) encontraron que al aplicar Si en plantas comerciales, el Fe aumentó en un 10 % a 20 % en brotes y en un 20 % a 40 % en las raíces. Al respecto, la adición de Si influye y regula fuertemente la disponibilidad de Fe en el apoplasto de la raíz, así como la expresión de genes involucrados en el transporte de Fe, tanto a nivel de la raíz como de hoja, lo que influye de manera positiva (Pavlovic et al., 2021) o negativa (Hernández-Apaoloza et al., 2020) en la absorción y translocación de Fe dentro de diferentes órganos y tejidos vegetales. De tal forma, Hernández-Apaoloza et al. (2020) reportan que al aplicar Si por vía radicular a plantas de pepino, la absorción de Fe disminuyó, mientras que, la aplicación vía foliar aumentó el Fe en las hojas.
La acumulación neta de Cu en rosa variedad “Snowflake” registró valores promedio menores (5.23 mg kg-1) a los reportados por Franco et al. (2013) quienes obtuvieron valores de 7.2 mg kg-1 en promedio para los cultivares de rosa en la sabana de Bogotá. En la presente investigación, todos los tratamientos de Si disminuyeron el Cu en las hojas de rosa, mientras que este efecto no se vio en las plantas testigo. Estos resultados concuerdan con Greger et al. (2018), quienes encontraron una reducción en la acumulación neta de Cu al aplicar Si en brotes de hortalizas, leguminosas y gramíneas. Al respecto, se ha encontrado que el Si, al depositarse en la pared celular, inmoviliza los iones de Cu y sintetiza moléculas de unión de Cu (metalotioneínas), tanto en los tejidos de la raíz como en los brotes (Li et al., 2008), por lo que disminuye el Cu, y en casos de toxicidad por Cu ayuda a mitigar el estrés.
Las concentraciones foliares de Mn, Zn y B presentaron diferencias significativas entre los diversos métodos de aplicación de Si en rosas variedad “Snowflake” (Tabla 3). El Mn y el Zn se mantuvieron dentro de los rangos normales de concentración foliar (Tabla 2) a diferencia del B, el cual mostró valores elevados al compararse con los datos obtenidos de cultivares de rosas en la sabana de Bogotá. Aun así, este exceso no generó síntomas de toxicidad por B en las plantas de rosa del estudio, por lo que se podría inferir que el cultivar de rosa “Snowflake” acumula una mayor cantidad de B que otras variedades.
Tabla 3 Concentraciones foliares de micronutrientes y elemento benéficos en plantas de rosa hidropónica “Snowflake” en el estado fenológico “rayando color” bajo diferentes métodos de aplicación de silicio
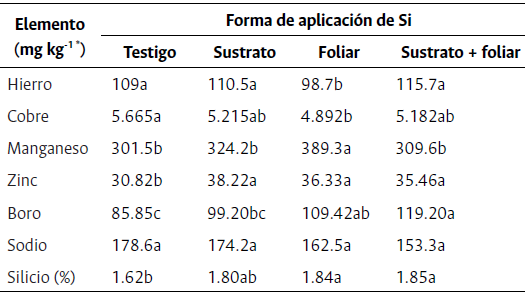
* Expresado en mg por kg de materia seca. Letras distintas en la misma fila indican diferencias estadísticas significativas entre tratamientos según la prueba de Duncan (P≤0.05).
Las plantas que presentaron las mayores concentraciones de Mn fueron tratadas con Si vía foliar, con lo que superaron en un 29.13 % a las plantas testigo. En la variedad “Charlotte”, Franco et al. (2013) encontraron Mn foliar de 155 mg kg-1 de MS, lo que es un valor bajo respecto al promedio para “Snowflake” (331.18 mg kg-1). En cuanto al efecto del Si sobre el Mn, de Oliveira et al. (2019) reportaron que la aplicación vía foliar o radicular de Si en plantas de sorgo no aumentó el Mn en las hojas. Además, el Si tiene un efecto muy fuerte sobre la capacidad de intercambio y formas de unión del Mn en el apoplasto de la hoja (90 % de Mn) a diferencia del simplasto (10 % de Mn) (Wiese et al., 2007), por lo que el Si no está directamente relacionado con la absorción del Mn, pero sí con su acumulación dentro de la planta. Esta fijación en las hojas del Mn podría explicar la mayor concentración que se obtuvo cuando el Si se aplicó vía foliar.
Por otro lado, todas las aplicaciones de Si aumentaron la concentración de Zn un 19 % con respecto al testigo independientemente del método de aplicación (Tabla 3). Los valores de Zn en rosa “Snowflake” son similares a los encontrados por Cabrera (2002) para la variedad “Bridal White”, con un promedio de 38 mg kg-1. En cuanto a la influencia positiva de las aplicaciones de Si sobre el Zn foliar, Soliman et al. (2021) reportaron resultados similares en durazno, ya que al aplicar aspersiones foliares de K2SiO3 a 3000 mg kg-1 incrementaron en un 21.3 % el Zn. Sin embargo, otros estudios muestran que la aplicación de Si reduce entre un 10 % y 20 % el Zn en hojas y brotes de arroz (Gu et al., 2012).
Al respecto, se ha encontrado que el Si inicialmente retrasa el transporte de Zn hacia los brotes debido a un aumento en los sitios de unión en la pared celular de la raíz, mejorando la adsorción de Zn en los depósitos de silicato y disminuyendo su concentración en los brotes (Hernández-Apaolaza et al., 2020). No obstante, Gu et al. (2012) afirman que este incremento en los sitios de unión también ocurre en las hojas, lo que podría finalmente aumentar el Zn foliar.
La concentración de B foliar en las plantas de rosa se vio altamente beneficiada con la aplicación de Si en comparación con el testigo. El mejor tratamiento fue la aplicación de Si al sustrato + foliar, seguido de la aplicación foliar y al sustrato, con aumentos del B en un 38.8 %, 27.4 % y 15.5 % en comparación con el testigo, respectivamente. Los valores obtenidos de B son mayores a los reportados para la variedad de rosa “Charlotte” en el estadio fenológico de “Rayando color”, con 68.2 mg kg-1 sin aplicaciones de Si (Triana et al., 2006). De forma similar, Kamenidou et al. (2010) en flores de Gerbera sp. encontraron que la aplicación de Si directamente al sustrato mejoró el B foliar (36 µg g-1 MS) frente al testigo (29 µg g-1 MS). Asimismo, Wiese et al. (2007) explican que, en plantas de pepino tratadas con Si, este unió más B a la pared celular y generó un menor transporte en el floema de las hojas.
Sodio y silicio
La concentración foliar de Na no tuvo diferencias significativas (Tabla 3) y mostró un promedio de 167.18 ±12.08 mg kg-1 de MS para todos los tratamientos a los 70 ddp. Estos valores son inferiores a los reportados por Cabrera (2002) quien en la variedad “Bridal White” injertada sobre “Natal Briar” (el mismo patrón utilizado en este experimento) encontró Na de 303 mg kg-1 sin la aplicación de Si. En cuanto al efecto de la aplicación de Si sobre el Na foliar, se ha obtenido que, bajocondiciones salinas, el Si puede actuar como una barrera mecánica para la traslocación de Na+ de la raíz a los brotes a través del bloqueo de la ruta apoplástica (Pavlovic et al., 2021), evitando así la acumulación de Na+ en las hojas.
La concentración foliar de Si en las plantas tratadas con distintos métodos de aplicación de K2SiO3 mostraron diferencias significativas con respecto al testigo (Tabla 3). Las plantas de todos los tratamientos con aplicación de Si aumentaron el Si foliar en un 13 % con relación al testigo. De igual forma, Kamenidou et al. (2010) reportaron resultados similares en flores de Gerbera sp., en las que los métodos de aplicación (sustrato o aspersión) y diferentes fuentes de Si (K2SiO3 o Na2SiO3) aumentaron el Si. Así mismo, se podría clasificar a las rosas “Snowflake” injertadas sobre “Natal Briar” como plantas moderadamente acumuladoras de Si (0.5 - 2 %) según la clasificación propuesta por Takahashi et al. (1990).
Conclusiones
El modelo logístico doble sigmoide es el que mejor describe el crecimiento de la longitud del tallo floral de rosa variedad “Snowflake” cultivada en la sabana de Bogotá. El Si, independientemente del método de aplicación, no mejoró la concentración foliar de N, P y K+, pero sí la de Ca, Mg y S. Específicamente, el suministro de Si por aspersiones foliares aumentó la concentración de Ca y Mg. Asimismo, aplicar Si de forma edáfica generó mayor traslocación de S a las hojas. La rosa “Snowflake” acumula una mayor cantidad de Fe, Mn y B en comparación con otras variedades sembradas en la zona. Las aplicaciones de Si favorecen la asimilación de Mn, B, Si y Zn, mientras que disminuyen el Cu. La rosa “Snowflake” se podría clasificar como una planta acumuladora de Si.

