










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Facultad de Medicina Veterinaria y de Zootecnia
Print version ISSN 0120-2952
Rev. Med. Vet. Zoot. vol.57 no.1 Bogotá Jan./Apr. 2010
REGIONES DEL CROMOSOMA 5 ASOCIADAS A CARACTERÍSTICAS
DE CRECIMIENTO EN GANADO CRIOLLO ROMOSINUANO
CHROMOSOMAL 5 REGIONS ASSOCIATED WITH GROWTH TRAITS
IN THE ROMOSINUANO CREOLE CATTLE
R. M. Ríos1, S. L. Castro2, D. J. Moreno3, J. S. Moncaleano4,
M. O. Santana5*, R. Barahona6**, B. F. Ariza7
Grupo de Investigación Genética Molecular Animal,
Facultad de Medicina Veterinaria y de Zootecnia,
Universidad Nacional de Colombia sede Bogotá
1mriosr@unal.edu.co, 2susancastr09@hotmail.com
3dianamoga@gmail.com, 4jsmoncaleanov@unal.edu.co
7mfarizab@bt.unal.edu.co
C. I. Turipaná, Investigador*
5mosantanar@gmail.com
Universidad Nacional sede Medellín**
6rolandobarahona2002@yahoo.com
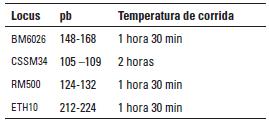
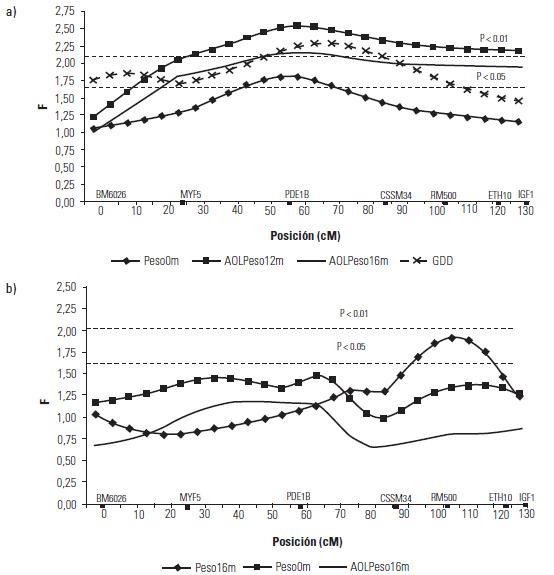
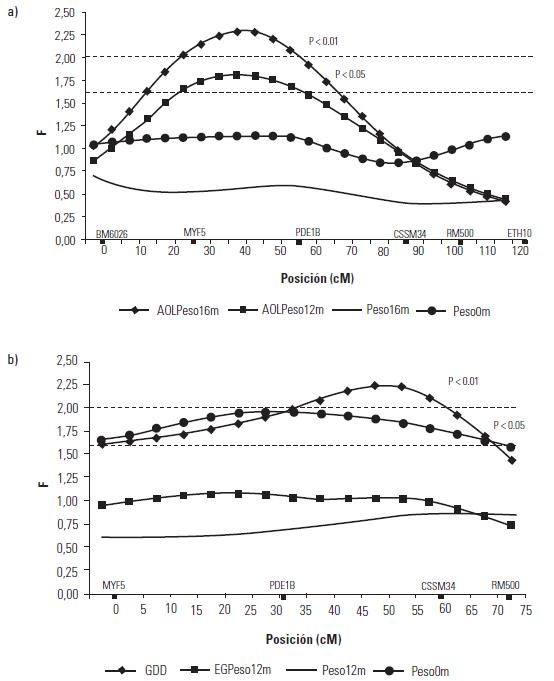
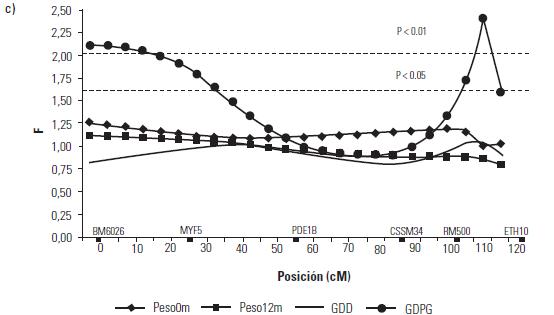
Artículo recibido: 10 de octubre de 2009; Aprobado: 5 de marzo de 2010 RESUMEN El crecimiento es una de las características de naturaleza cuantitativa que influye en la calidad de la canal. En el presente estudio se identificaron las regiones cromosomales responsables para la variación del crecimiento en el cromosoma 5 en una población de ganado criollo romosinuano. Se evaluaron en 72 progenies las características de peso al nacimiento, peso al destete, peso a los 12 meses y a los 16 meses, área de ojo del lomo a los 12 y 16 meses, espesor de grasa dorsal a los 12 y 16 meses, espesor de grasa del anca a los 12 y 16 meses, ganancia diaria predestete y posdestete, al igual que los genotipos de tres polimorfismos de nucleótido simple de los genes MYF5, PDE1B e IGF1 y de cuatro microsatélites BM6026, CSSM34, RM500, ETH10, distribuidos a lo largo del cromosoma. Se realizó un análisis de regresión linear el cual mostró el efecto de seis loci de rasgos cuantitativos (QTL) asociados a características de crecimiento. Cinco QTL fueron significativos (p≤0,05) para las características de peso al nacimiento, peso a los 16 meses, área del ojo de lomo a los 12 y 16 meses y ganancia diaria posdestete y un QTL se encontró con una significancia de p≤0,01 para la característica ganancia diaria predestete. Los resultados demostraron que los genes MYF5, PDE1B, IGF1 pueden ser genes candidatos posicionales que inciden en la variación del crecimiento para la calidad de la canal en el ganado romosinuano. Palabras clave: IGF1, ligamiento, marcadores microsatélite, MYF5, PDE1B, ultrasonido. Growth is a quantitative trait that influences the carcass quality. In the present study, bovine chromosomal 5 regions responsible for growth variations have been identified in a romosinuano creole cattle population. The traits birth weight, weaning weight, 12 months weight, 16 months weight, the rib eye area at 12 and 16 months, back fat thickness at 12 and 16 months, rump fat thickness at 12 and 16 months, preweaning and postweaning daily gain were evaluated in 72 progenies, as well as the genotypes of 3 SNPs from the MYF5, PDE1B, IGF1 genes and 4 microsatellites (BM6026, CSSM34, RM500, ETH10) distributed along the chromosome. A linear regression analysis showed the six QTL effect associated with growth traits. Five QTL were significant at p≤0.05 for the triait birth weight, weight at 16 months, rib eye area at 12 and 16 months, and preweaning daily gain interval between 45 and 105 cM and a QTL showed a level significance of p ≤ 0.01 for the postweaning daily gain. It was demostrated according to the position of the QTLs identified in the present study, that the MYF5, PDE1B and IGF1 genes might be positional candidate genes affecting the growth variation for carcass quality in romosinuano cattle. Key words: IGF1, genes, linkage, microsatellite marker, MYF5, PDE1B, ultrasound. INTRODUCCIÓN El objetivo de los métodos del análisis genómico modernos para especies de importancia en la ganadería se fundamenta en la identificación de regiones cromosomales llamadas loci de rasgos cuantitativos (QTL) responsables por la variación en el fenotipo tanto de la producción como de la calidad de las canales en las especies de importancia zootécnica. Esta metodología se ha convertido en una herramienta de apoyo a los programas de mejoramiento tradicionales ya que puede ser aplicada a edades tempranas en los animales (1). Las características cuantitativas más importantes en producción animal están bajo el control de varios genes y algunos de ellos ejercen un mayor o menor efecto sobre estos rasgos bajo la influencia del medioambiente. Además, otros aspectos importantes para tener en cuenta son las tasas de mutación de los genes tanto dentro y entre poblaciones y sus mecanismos de acción (2). Varios estudios han identificado QTL para el peso al nacimiento en cinco cromosomas bovinos: BTA2 (3), BTA3 (3, 4), BTA5 (3, 4, 5, 6), BTA6 (4, 7, 8) y BTA21 (3, 4, 8). En el cromosoma 5 también se han identificado QTL con efectos significativos para otras características productivas como son el porcentaje de ovulación (9, 10), la calidad de la canal (4, 7, 11) y el crecimiento. En el estudio reportado por Li et ál. (5) en dos líneas sintéticas de ganado Beefbooster se identificaron tres regiones cromosomales del BTA5 (0 a 30 cM, 55 a 70 cM, y 70 a 80 cM) asociados con peso al nacimiento, ganancia diaria predestete y ganancia diaria durante la alimentación (eficiencia alimentaria) utilizando 16 marcadores; en este trabajo alcanzaron un cubrimiento del 93% del cromosoma con una extensión de 114 cM. En un trabajo posterior estos mismos investigadores (12), analizando características de crecimiento en las regiones previamente identificadas, en dos líneas comerciales de individuos Bos taurus encontraron asociación entre polimorfismos de nucleótido simple (SNP) de dos genes candidatos, el factor miogénico 5 (MYF5) y el factor de crecimiento insulinico 1 (IGF1). De la misma manera, Casas et ál. (3), en cruces de ganados Bos indicus x Bos taurus, identificaron varios QTL en el BTA5 relacionados con características de crecimiento (peso al nacimiento) en la región de 30 a 70 cM y de la composición de las canales (área del ojo del lomo y peso de la canal en caliente) a los 29-74 cM. Para la característica de peso al nacimiento varios autores han reportado QTL en este mismo cromosoma como Kim et ál. (4), en una población de angusxBrahman en la región de 45,5 cM a 71,6 cM; Machado et ál. (6), en la raza Canchim a los 82,6 cM, y Gasparin et ál. (13), en un cruce de GyrxHolandés a los 69 cM. En el BTa5 también se han reportado dos QTL para la característica de peso a los doce meses a los 45,5 a 71,6 cM (4) y 76,18 cM (6). En la literatura sólo se encuentra un estudio de asociación entre marcadores y peso al nacimiento llevado a cabo por ariza et ál. (14), quienes en el cromosoma cinco de la raza romosinuano identificaron dos marcadores tipo microsatélite (BMS772 y BM2830), y encontraron una asociación significativa con un efecto positivo para el marcador BMS772 con peso al nacimiento en los homocigotos mientras que el marcador BM2830 mostró un mayor peso promedio al nacimiento en los heterocigotos. El objetivo del presente estudio fue identificar QTL que tengan efecto sobre las características de crecimiento a partir de la construcción de un mapa de ligamiento para determinar el orden de los marcadores de los genes factor miogénico 5 (MYF5), fosfodiesterasa calmodulin estimuladora de los nucleótidos cíclicos (PDE1B) y el factor de crecimiento insulínico 1 (IGF1), y los microsatélites BM6026, CSSM34, RM500, ETH10 para el cromosoma 5 bovino (BTA5) en una población de la raza romosinuano. MATERIALES Y MÃTODOS El estudio se realizó en el Centro de investigación Turipaná de Corpoica (Cereté, Córdoba, Colombia), con una altura de 17 msnm. Se evaluaron 72 progenies de la raza romosinuano, dentro de un diseño de medios hermanos paternos distribuidos en 12 familias (12 padres). Los registros fenotípicos de peso se tomaron de las bases de datos de la granja teniendo en cuenta peso y fecha al nacimiento, peso y edad al destete, peso a los 12 meses, y peso y edad al día de la evaluación ecográfica. Además, se tuvo en cuenta la identificación del padre (número del padre), la identificación de la madre (número de la madre y número de partos), el año de nacimiento del ternero (2003 ó 2004), y la época de nacimiento del ternero (verano o invierno). Las medidas fenotípicas evaluadas por ultrasonido fueron: el área de ojo del lomo (AOL), el espesor de grasa dorsal (EG) y el espesor de grasa en el anca (P8). Las medidas se tomaron previo ayuno de 12 horas con un ecógrafo marca aquila equipado con sonda de 18 cm, asP 3,5 MHz (Pie-medical vet., Holanda) y con una almohadilla de acople anatómico. Las imágenes se interpretaron con el software Open Data Transfer (ODT, University of Tennessee USA). Identificación de los genotipos A partir de las muestras de sangre se aisló y purificó el ADN genómico según el protocolo descrito por sambrook et ál. (15). Los genotipos de los genes MYF5, PDE1B, IGF1, y los microsatélites BM6026, CSSM34, RM500, ETH10, se amplificaron utilizando los iniciadores que se presentan en la tabla 1. TABLA 1. Secuencia de los iniciadores Forward y Reverse de los marcadores que se utilizaron en el presente estudio y su temperatura de anillamiento * Corresponde a un marcador de polimorfismo de nucleótido simple (SNP). Los iniciadores para amplificar el segmento del gen MYF5 se seleccionaron a partir de la información pública suministrada por Li et ál. (12); para el gen PDE1B se escogieron de acuerdo con lo reportado por stone et ál. (16), y para el gen IGF1 se seleccionaron a partir de la información del Molecular animals Genetics, CSIRO (australia, comunicación personal) (17) (tabla 1). Las condiciones de la reacción de cadena de la polimerasa (PCR) se ajustaron para un volumen final de 12,5 µl, que contenían 4 µl de ADN genómico (20ng-µl), 1,5 µl de MgCl2 (1,5 mM), 2,5 µl de dnTP (12,5 mM cada uno, mezclados con Buffer 1x), 0,5 µl de Taq polimerasa preparada en el laboratorio (0,2U), 1 µl de cada iniciador (20 ng-µl), 2 µl de agua grado molecular y una gota de aceite mineral. Las condiciones de amplificación incluyeron 95°C por 3 minutos durante 1 ciclo, 95°C por 30 segundos. Las temperaturas de anillamiento variaron entre 54-60°C por 1 minuto de acuerdo con el marcador utilizado (tabla 1), 72°C por 1 minuto 30 segundos durante 30 ciclos, seguido de 1 ciclo a 72°C por 10 minutos, y finalmente 1 ciclo a 30°C por 1 minuto. Para la detección de los genotipos se empleó la metodología indicada para cada marcador, así: para el marcador MYF5 se usó la técnica de polimorfismos de longitud de fragmentos digeridos (RFLP), usando la enzima Taqi y los productos de digestión se separaron en geles de agarosa al 1,5%; para el marcador PDE1B se empleó el polimorfismo conformacional de cadena sencilla (SSCP) / Heterodúplex, y para el IGF1 se utilizó SSCP. Los microsatélites se genotipificaron en geles denaturantes de poliacrilamida (29:1 6X con urea 7,5 M); el tamaño de los fragmentos en pares de bases (pb) de los microsatélites, y el tiempo de corrida del gel se describen en la tabla 2. Los geles se corrieron a 1500 voltios a 50°C en buffer TBE 1X. Posteriormente, los geles fueron teñidos con nitrato de plata y los genotipos registrados en una base de datos después de haber sido confrontados por dos personas diferentes al lector inicial. TABLA 2. Tamaño de los fragmentos y temperatura de corrida de los geles de acrilamida Construcción del mapa de ligamiento Para la construcción del mapa de ligamiento se empleó el programa Animap (19) el cual utiliza la función de Kosambi para calcular las frecuencias de recombinación entre los marcadores considerando un grado intermedio del fenómeno de interferencia entre los alelos o de la probabilidad de entrecruzamiento determinando el ligamiento entre los marcadores y el orden de los mismos a lo largo del cromosoma con un valor de LOd mayor a 3,0 (p ≤ 0,001) para cada par de marcadores. Detección de QTL para características de crecimiento El análisis para los QTL se realizó mediante el programa QTL express (20) utilizando la opción de "análisis para un sólo QTL". Se evaluaron los efectos entre familias (n=12), con un total de 84 individuos (72 progenies y 12 padres) para las características de crecimiento: peso al nacimiento (peso 0m), peso al destete (peso9m), peso al año (peso12m) , peso a los 16 meses (peso16m), área de ojo del lomo a los 12 y 16 meses (AOL peso12m y AOL peso16m), espesor de grasa dorsal a los 12 y 16 meses (EG peso12 m y EG peso16m), espesor de grasa del anca a los 12 y 16 meses (P8 peso12m y P8 peso16m), ganancia diaria predestete y posdestete (GDD y GDPG) y los genotipos de los genes MYF5, PDE1B, IGF1, y de los microsatélites BM6026, CSSM34, RM500, ETH10. Para estimar el efecto del QTL se realizaron análisis que cubrieron la extensión del BTA5 a intervalos de 5 cM de acuerdo con el mapa de ligamiento para estos marcadores en la raza romosinuano, estableciendo como estrategia de análisis el agrupamiento de los marcadores en diferentes grupos de ligamientos como pares, tríos, tétradas y así sucesivamente, hasta analizar todos los marcadores como un solo grupo. El F estadístico se determinó por comparación de mínimos cuadrados obtenidos de la regresión dentro de la familia para el mínimo cuadrado residual. Los grados de libertad del numerador (S) corresponden al número de padres y el denominador corresponde a N-2S donde N es el número de la pro-genie. Los umbrales de significancia se calcularon utilizando el método de permutación de Churchill et ál. (21). Los test de significancia para la presencia de un QTL se diseñaron a un intervalo de 10 cM y se repitieron 1000 veces para la construcción de la distribución del test estadístico bajo la hipótesis nula. Método estadístico Las medidas de peso y ultrasonido fueron ajustadas antes de los análisis. Se utilizó el siguiente modelo estadístico de regresión para la identificación de los QTL: Yij = ai+biXij+eij, donde Yij: es el valor de la característica del individuo j, descendiente del padre i; ai: es el efecto poligénico para la familia de medios hermanos paternos i; bi: es el coeficiente de regresión en la familia i, como es el efecto de la sustitución alélica para un QTL dado; Xij: es la probabilidad condicional para el individuo j de heredar el primer haplotipo del padre i; eij: es el efecto residual. El análisis de regresión se realizó entre familias y los valores de la raza son valores ponderados por su fiabilidad (esto es el número de hijos). El análisis provee relaciones de F entre los grupos de ligamiento con el máximo valor que es la posición más probable del QTL. RESULTADOS Todos los marcadores mostraron segregación mendeliana y demostraron ser informativos para ser incluidos dentro del grupo de ligamiento correspondiente al cromosoma 5 bovino. En la figura 1 se muestra el orden y las distancias de mapa en cM de Kosambi. Los errores generados por dobles recombinantes fueron removidos. Identificación de QTL En la tabla 3 se indican las características evaluadas, el agrupamiento de los marcadores para el BTA5 en cada uno de los ensayos efectuados en donde se identificaron QTL significativos. Además, se describe la posición estimada del QTL, los valores del F estadístico, el valor del F calculado, el porcentaje de verosimilitud (LR) y la familia significativa con su valor absoluto t (Ab(t)) que indica la evidencia del QTL en un análisis entre familias; estos resultados fueron dados por el programa QTLexpress. El valor de significancia de p es la diferencia del F estadístico en relación con el F calculado e indica la significancia entre p ≤ 0,05 y p ≤ 0,01 para los QTL. TABLA 3. QTL detectados por el programa QTLExpress para el BTA5 utilizando diferentes grupos Como se observa en los resultados obtenidos en la tabla 3, para la característica de peso0m se detectó un QTL (p≤0,05) para el grupo de marcadores BM6026MYF5-PDE1B-CSSM34-RM500ETH10-IGF1 a los 45 cM delimitado por los marcadores MYF5 y PDE1B (véase figura 2a), siendo las familias significativas la 4 con un valor de Ab(t) de 2,23, lo cual evidenció la presencia de un QTL en el análisis entre familias. Para la característica de peso 16 se identificó un QTL en la posición de 105 cM para el grupo de marcadores BM6026-PDE1B-CSSM34-IGF1 con un p≤0,05. El QTL se encontró delimitado por los marcadores CSSM34-IGF1 (véanse tabla 3 y figura 2b). La familia 6 fue significativa con un Ab(t) de 2,63. En la posición de 40 cM se identificaron dos QTL para las características de AOL Peso12m y AOL Peso16m, para los grupos de marcadores BM6026MYF5-PDE1B-ETH10 con un p≤0,05 delimitado por los marcadores MYF5 (24 cM) y PDE1B (55 cM). Las familias informativas fueron la 4 y 1 con un ab(t) de 2,44 y 3,06 respectivamente (véanse tabla 3 y figura 3a). Para la característica GDD se identificó un QTL a los 50 cM (p≤0,01) para el grupo de marcadores MYF5-PDE1BCSSM34-RM500 siendo la familia significativa la 12 con un ab(t) de 3,46 (véase tabla 3 y figura 3b). Se identificó un QTL para la característica GDPG con una significancia de (p≤0,05) para los grupos de marcadores BM6026-CSSM34-ETH10 a los 110 cM (véase tabla 3 y figura 3c). La familia 8 se observó como la más significativa con un Ab(t) de 3,62. DISCUSIÓN DE RESULTADOS En este trabajo se generó el segundo mapa de ligamiento para el BTA5 en la raza criolla romosinuano después del reportado por ariza et ál. (14). La certeza en la ubicación del orden de los marcadores a lo largo del mapa es considerada de gran importancia para la ubicación de los genes responsables de características de importancia zootécnica. El orden de los marcadores descrito en el presente estudio coincide totalmente con los ma-pas reportados por otros autores como Barendse et ál. (22), Ozawa et ál. (23) y donato et ál. (24) lo que permitiría comparar el mapa de ligamiento de la raza romosinuano con los mapas de otras razas ya estudiadas facilitando la detección de QTL o genes candidatos relacionados con características de crecimiento. Además, este es el primer estudio de QTL asociados a características de crecimiento para la raza criolla romosinuano que se reporta en la literatura. Por tan-to, los resultados obtenidos son de gran valor y servirán de base para futuros estudios de genes candidatos relacionados con características de producción. Así, para la discusión de estos resultados no se tiene un punto de referencia y por consiguiente esta comparación se hace con trabajos semejantes en otras razas de origen europeo y cruces de estas con Bos indicus. El QTL para peso0m identificado en este estudio se ubicó a los 45 cM, de modo similar al QTL reportado por Casas et ál. (3) (30 a 70 cM) y Kim et ál. (4) (45,5 a 71,6 cM) para esta misma característica. Las posición de este mismo QTL es cercana a las regiones cromosomales reportadas por Li et ál. (5) y Li et ál. (12) quienes ubicaron un QTL entre 19 a 28,6 cM, pero se encuentra más lejano al QTL reportado por otros auto-res como Machado et ál. (6) (82,9 cM) y Gasparin et ál. (13) (69 cM). Estos hallazgos confirman la existencia de un QTL para peso0m en el BTA5 en la raza romosinuano; aunque su posición varía con respecto a algunos estudios, las diferencias pueden atribuirse a la población estudiada y a los marcadores empleados en cada análisis. Para la característica de peso16m se reporta por primera un QTL en el BTA5 ubicado a los 105 cM y delimitado por los marcadores RM500 y ETH10, los cuales están relacionados con crecimiento, como lo reportaron previamente Machado et ál. (6) y Li et ál. (12). Aunque este QTL no ha sido identificado para otras razas bovinas es importante destacar su efecto debido al crecimiento más lento que se observa en el ganado criollo colombiano. Para las características de AOL peso12m y AOLpeso16m en este estudio se detectaron dos QTL ubicados a los 40 cM, de modo similar al análisis realizado por Casas et ál. (3) para estas mismas características. Esta similitud permitiría posicionar genes candidatos ya identificados en otras razas para estos rasgos y extrapolar estos datos a la raza romosinuano. Los QTL detectados para ganancias de peso diarias GDD y GDPG se ubicaron a los 50 cM y 110 cM respectivamente. La ubicación del primer QTL coincide con la región reportada por Li et ál. (5) y Li et ál. (12) a los 73 cM. Para la característica GDPG, también se reporta por primera vez un QTL en el BTA5 en la raza romosinuano. Este QTL serviría de punto de partida para realizar el seguimiento durante esta etapa de crecimiento, lo que permitiría seleccionar animales con mejor comportamiento para su uso como parentales. CONCLUSIONES Se reporta por primera vez un mapa de ligamiento en el cromosoma 5 bovino utilizando los marcadores BM6026, MYF5, PDE1B, CSSM34, RM500, ETH10, IGF1 para la raza romosinuano. El orden de los marcadores encontrados en este estudio es similar a otras publicaciones recientes de los mapas de ligamiento para el cromosoma 5 bovino, aunque se presentan variaciones en las distancias entre los marcadores, lo que se puede deber a la raza y los genes y/o marcadores analizados. Se reportan por primera vez seis QTL asociados a características de crecimiento en la raza criollo romosinuano. En el análisis entre familias se detectaron QTL en el BTA5 con significancia p ≤ 0,05 para las características de crecimiento relacionadas con peso: peso0m, peso16m, GDPG, para las características de crecimiento medidas por ecografía: AOL peso12m, AOL peso16m y con significancia de p ≤ 0,01 para la característica GDD. Las características de crecimiento: peso0m, AOL peso12m, AOL peso16m se asocian con la presencia de los marcadores MYF5-PDE1B debido a que el umbral del QTL estuvo cercano a estos marcadores. La selección de la progenie que heredará los rasgos analizados se facilitará debido al análisis de los QTL lo que per-mite identificar la familia y, por tanto, el parental que presenta el mayor efecto sobre la característica de interés. Se recomienda ampliar los estudios utilizando los toros de las familias 6, 8 y 12 para incrementar el número de progenies dentro de éstas y así poder evaluar la segregación de los rasgos en la población. Con el propósito de aumentar la eficacia en la construcción de un mapa de ligamiento genético y determinar las regiones que presentan QTL asociados a características de crecimiento en bovinos, se recomienda realizar estudios incluyendo individuos producto del cruzamiento del F2 debido a que poseen mayor poder para detectar efectos aditivos entre los alelos y permite una mayor información de los eventos recombinantes entre los gametos parentales. FIGURA 2. Perfil del F estadístico para las características de a) QTL para peso0m, b) QTL para peso16m. En las líneas horizontales se indican los umbrales de significancia para un 5% (línea interrumpida) y 1% (línea y punto) definidos por permutaciones. Los triángulos negros en el eje X indican la posición de los marcadores a lo largo del mapa. FIGURA 3. Perfil del F estadístico para las características de a) QTL para AOL peso12m y AOL peso16m. b) QTL para GDD y c) QTL para GDPG. En las líneas horizontales se indican los umbrales de significancia para un 5% (línea interrumpida) y 1% (línea y punto) definidos por permutaciones. Los triángulos negros en el eje X indican la posición de los marcadores a lo largo del mapa).
ABSTRACT
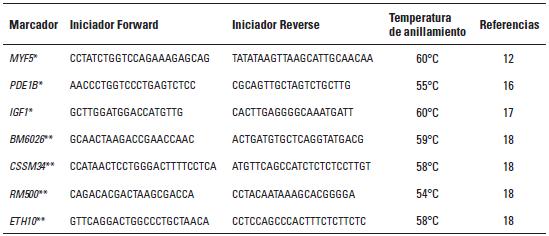
** Corresponde a un marcador de tipo microsatélite.
para los microsatélites BM6026, CSSM34, RM500, ETH10
Mapa de ligamiento 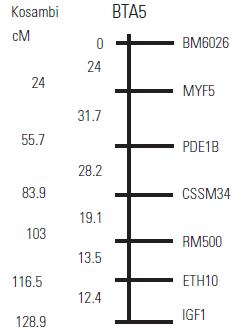
FIGURA 1. Mapa de ligamiento generado por Animap para el BTA5.
Las fracciones de recombinación, el valor LOD y la distancia de mapa entre marcadores fueron generados utilizando la opción de ligamiento de dos puntos (two point). Las fracciones de recombinación y el valor de LOD entre pares de marcadores fueron: BM6026-MYF5 (lod 0,007, θ = 0,4), MYF5-PDE1B (lod 0,036, θ = 0,3), PDE1BCSSM34 (lod 1,58, θ = 0,3), CSSM34-RM500 (lod 2,96, θ = 0,2), RM500-ETH10 (lod 2,95, θ = 0,2), ETH10-IGF1 (lod 4,14, θ = 0,2).
de marcadores para las características fenotípicas evaluadas. 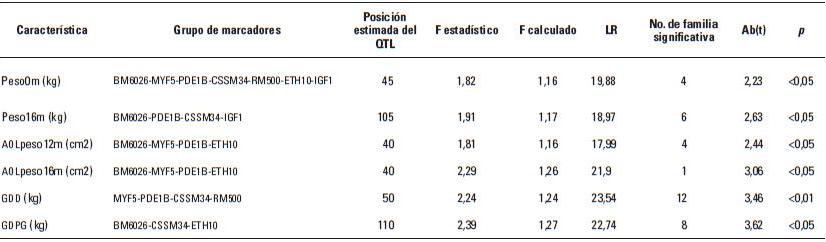
REFERENCIAS
1. Bryne PF, McMullen MD. Defining genes for agricultural traits: QTL análisis and the candidate gene approach. Probe 1996; 7: 24-27. [ Links ]
2. Moro J, Hayes J. Métodos de mapeo de loci de rasgos cuantitativos y sus aplicaciones potenciales en la industria lechera. Téc Pec Méx 2006; 44 (3): 329-350. [ Links ]
3. Casas E, Shackelford SD, Keele JW, Koohmaraie M, Smith TP, Stone RT. Detection of quantitative trait loci for growth and carcass composition in cattle. J Anim Sci 2003; 81: 2976-2983. [ Links ]
4. Kim J, Farnir F, Savell J, Taylor J. Detection of quantitative trait loci for growth and beef carcass fatness traits in a cross between Bos taurus (angus) and Bos indicus (Brahman) cattle. J Anim Sci 2003; 81: 1933-1942. [ Links ]
5. Li C, Basarab J, Snelling W, Benkel B, Murdoch B, Moore S. The identification of common haplotypes on bovine chromosome 5 within commercial lines of Bos taurus and their associations with growth traits. J Anim Sci 2002; 80: 1187-1194. [ Links ]
6. Machado M, Alencar M, Pereira A, Oliveira H, Casas A, Coutinho L, Regitano L. QTL affecting body weigth in a candidate región of cattle chromosome 5. Genetics and Molecular Biology 2003; 26 (3): 259-265. [ Links ]
7. Casas E, Shackelford S, Keele J, Stone R, Kappes S, Koomaraie M. Quantitative trait loci affecting growth and carcass composition of cattle segregating alternate forms of myostatin. J Anim Sci 2000; 78: 560-569. [ Links ]
8. Kneeland J, Li C, Basarab J, Snelling WM, Benkel B, Murdoch B, Hansen C, Moore SS. Identification and fine mapping of quantitative trait loci for growth traits on bovine chromosome 2, 6, 14, 19, 21 and 23 within one comercial line of Bos Taurus. J Anim Sci 2004; 82: 3405-3414. [ Links ]
9. Kirkpatrick B, Becky M, Gregory K. Mapping quantitative trait loci for bovine ovulation rate. Mamm Genome 2000; 11: 136-139. [ Links ]
10. Arias J, Kirkpatrick B. Mapping of bovine ovulation rate QTL: An analytical approach for three generation pedigree. Anim Genet 2004; 35: 7-13. [ Links ]
11. Stone R, Keele J, Shackelford S, Kappes S, Koomaraie M. A primary screen of the bovine genome for quantitative trait loci affecting carcass and growth traits. J Anim Sci 1999; 77: 1379-1384. [ Links ]
12. Li C, Basarab J, Snelling WM, Benkel B, Murdoch B, Hansen C, Moore SS. Assessment of positional candidate genes MYF5 and IGF1 for growth on bovine chromosome 5 in commercial lines of Bos taurus. J Anim Sci. 2004; 82: 1-7. [ Links ]
13. Gasparin G, Miyata M, Lehmann L, Martinez M, Vinícius M, Barbosa da Silva G, Machado M, Campos A. Quantitative trait locus affecting birth weight on bovine chromosome 5 in a F2 Gyr x Holstein population. Genetics and Molecular Biology 2005; 28: 670-676. [ Links ]
14. Ariza F, Guerrero A, Fajardo C, Franco L, Vargas C, Ayala C, Peña L, Roa L, Barrera G, Martínez R. Asociación de marcadores moleculares con peso al nacimiento en el ganado criollo romosinuano. V Simposio Iberoamericano sobre la conservación y utilización de recursos zoogenéticos. Universidad Nacional de Perú; 2004. [ Links ]
15. Sambrook F, Fritsch EF, Maniatis T. Molecular cloning; a laboratory manual, Second Editión. Cold Spring Harbor Laboratory Press; 1989. [ Links ]
16. Stone RT, Casas E, Smith TP, Keele JW, Harhay G, Bennett GL, Koohmaraie M, Wheeler TL, Shackelford SD, Snelling WM. Identification of genetic markers for fat deposition and meat tenderness on bovine chromosome 5: development of a low-density single nucleotide polymorphism map. J Anim Sci 2005; 83: 2280-2288. [ Links ]
17. Molecular Animals Genetics, CSIRO (australia, comunicación personal); 2000. [ Links ]
18. Barendse W, Armitage SM, Kossarek LM, Shalom A, Kirkpatrick BW, Ryan AM, Clayton D, Li L, Neibergs HL, Zhang NL. A genetic linkage map of the bovine genome. Nat Genet 1994, 3: 227-235. [ Links ]
19. Georges M, Nielsen D, Mackinnon M. Mapping quantitative trait loci controlling milk production in dairy cattle by exploiting progeny testing. Genetics 1995; 139: 907-920. [ Links ]
20. Seaton G, Haley SA, Knott M, Kearsey PM, Visscher D. QTL express: mapping quantitative trait loci in simple and complex pedigrees. Bioinformatics 2004; 18: 339-340. [ Links ]
21. Churchill GA, Doerge RW. Empirical threshold values for quantitative trait mapping. Genetics 1994; 138: 963-971. [ Links ]
22. Barendse W, Vaiman D, Kemp SJ, Sugimoto Y, Armitage SM, Williams JL, Sun HS, Eggen A, Agaba M, Aleyasin SA. A medium-density genetic linkage map of the bovine genome. Mamm. Genome 1997; 8: 21-28. [ Links ]
23. Ozawa A, Band M, Larson J, Donovan J, Green C, Womack J, Lewin H. Comparative organization of cattle chromosome 5 revealed by comparative mapping by annotation and sequence similarity and radiation hybrid mapping. Proc Natl Acad Sci 2000; 97 (8): 4150-4155. [ Links ]
24. Donato M, Brenneman RA, Stelly DM, Womack JE, Taylor JF. A methodological approach for the construction of a radiation hybrid map of bovine chromosome 5. Genetics and Molecular Biology 2004; 27 (1): 22-32. [ Links ]

