











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkINTRODUCCIÓN
Durante el periodo de transición a la lactancia (3 semanas antes y 3 semanas después del parto), la cantidad de ácidos grasos no esterificados (AGNES) que llega al hígado sobrepasa su capacidad de oxidación. El hígado utiliza los ácidos grasos (AG) principalmente como fuente energética y los excedentes que no realizan oxidación mitocondrial deben ser esterificados e integrados a la lipoproteína de muy baja densidad (VLDL, por sus siglas en inglés) para ser transportados a los tejidos demandantes de energía. Durante el periodo de transición a la lactancia ambos procesos, oxidación mitocondrial y ensamblaje de los triglicéridos (TG) a VLDL, pueden verse disminuidos debido a la baja disponibilidad de componentes claves como la metionina, colina y carnitina (Drackley et al. 2001; Piepenbrink et al. 2004; Piepenbrink y Overton 2003).
La disponibilidad de fosfatidilcolina puede ser un factor limitante en el hígado de las vacas durante la lactancia temprana debido a la depresión en el consumo de materia seca, la cual limita la ingestión adecuada de nutrientes entre los que se destacan, sus precursores, colina y metionina (Piepenbrink y Overton 2003). Otro factor importante para la eficacia de la suplementación con colina es el nivel de metionina en la dieta; algunas investigaciones que estudiaron la eficacia en la suplementación con colina reportaron niveles de metionina dietaría de 2,1% de la proteína digestible a nivel intestinal (Zom et al. 2011), similares a las recomendaciones hechas por la NRC (2001) y Rulquin et al. (2001) para este tipo de animales. Entre tanto, se ha estimado que para el pasto kikuyo (Cenchrus clandestinus) los aminoácidos esenciales con menor concentración relativa son, en su orden, lisina y metionina (Clark et al. 1992; Correa 2006). En este sentido, la composición de aminoácidos del forraje puede ser utilizada como un indicador del perfil de aminoácidos de la proteína que no es degradada en el rumen (Correa at al. 2008; Tedeschi et al. 2001); basado en lo cual, es de esperar un bajo aporte de los aminoácidos lisina y metionina para la absorción intestinal en dietas basadas en pasto kikuyo. Bajo esta situación se podría incrementar la capacidad exportadora de TG desde el hígado hacia otros tejidos si durante el periodo de transición se complementan las vacas con niveles bajos de metionina y colina conjuntamente, o si se complementan únicamente con niveles altos de colina. El objetivo de esta investigación fue evaluar el efecto del nivel de complementación dietaría con metionina y colina sobre la concentración de triacilglicéridos hepáticos.
MATERIAL Y MÉTODOS
La propuesta para la realización de este estudio fue avalada por el comité de ética en investigación de la Universidad Nacional de Colombia, sede Medellín, en su comunicación CEMED 200. La fase experimental del estudio se realizó en el centro Paysandú de la Universidad Nacional, sede Medellín, correspondiente a la formación bmh-MB, según la clasificación de Holdridge (Espinal 1977).
Animales y dieta
El estudio se desarrolló con vacas Holstein de dos a seis partos, que se encontraban entre el día 260 de gestación y el 20 posparto, las cuales se pesaron y posteriormente se calificó su condición corporal al inicio y al final del experimento siguiendo metodologías previamente descritas por Edmonson et al (1989) y Hady et al. (1994).
La descripción de los animales utilizados en el experimento se detalla en la Tabla 1. Las vacas pastorearon praderas de kikuyo (Cenchrus clandestinus) y recibieron un complemento alimenticio, el cual, acorde con el tratamiento, recibía la adición de la respectiva fuente de cada factor lipotrópico: para metionina, Aminoshure-M® (75% L-Metionina, Balchem USA) y para colina, Reashure-M® (25% de Colina, Balchem USA). La composición de los suplementos alimenticios se describe en la Tabla 2. Se utilizaron 6 animales por tratamiento, los cuales fueron T0: grupo control, T1: 20 g/d de Aminoshure-M®, T2: 20 g/d de Aminoshure-M® + 60 g/d de Reashure-M® y T3: 20 g/d de Aminoshure-M® + 120 g/d de Reashure-M® La caracterización nutricional de los forrajes y de los suplementos alimenticios se describe en Tabla 3. La cinética de la degradación ruminal del Reashure-M® y del Aminosshure-M® fue descrita previamentre por Montoya et al. (2015) quienes reportaron una degradabilidad ruminal a las 48 horas de 3,2% para el Reashure® y de 55,7% para el Aminoshure®.
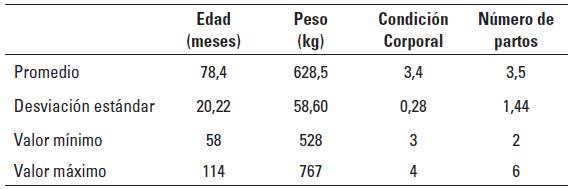
TABLA 2 Porcentaje de ingredientes utilizados en la formulación de los complementos alimenticios.
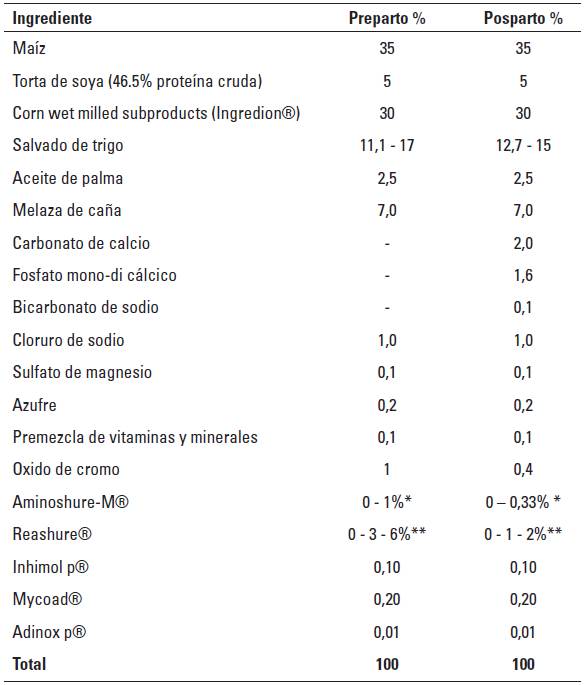
* Estas concentraciones garantizan un consumo de 20 g/d de Aminoshure-M® (15 g/d de metionina), con consumos de complemento alimenticio de 2,0 y 6,0 kg/d durante el preparto y el posparto respectivamente.
** Estas concentraciones garantizan un consumo de 30 o 60 g/d de Reashure-® (15 o 30 g/d de colina) según el nivel de complementación (bajo o alto), con consumos de complemento alimenticio de 2,0 y 6,0 kg/d durante el preparto y el posparto respectivamente.
TABLA 3 Composición química del pasto kikuyo (Cenchrus clandestinus) y del complemento alimenticio utilizados en la investigación.
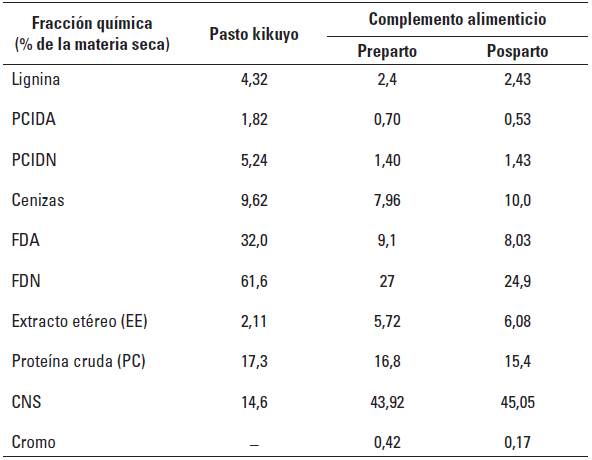
PCIDA: Proteína cruda insoluble en detergente acido, PCIDN: Proteína cruda insoluble en detergente neutro, FDN: Fibra en detergente neutro, FDA: Fibra en detergente ácido, CNS: Carbohidratos no estructurales = 100 - (PC + FDN + Cenizas + EE) + PCIDN (NRC 2001).
Toma de muestras y determinaciones
De cada vaca se tomaron tres muestras de sangre en tubos con y sin anticoagulante el día 270 de gestación, el día del parto y los días 10 y 20 de posparto, respectivamente. Adicionalmente, en los días 270 de gestación y 10 y 20 de posparto se tomaron biopsias de hígado utilizando el procedimiento descrito por Galvis et al. (2016). Las muestras de sangre se centrifugaron a 5000 rpm durante 5 min, se les separó el suero o el plasma (según el caso) y se envasaron en alícuotas de 2,0 ml. Para su transporte las muestras se almacenaron a -10°C y se mantuvieron a -72°C hasta su análisis.
En las muestras de suero se cuantificaron las concentraciones de AGNES, β-Hidroxibutirato (BHB) y TG. Para estimar la concentración de AGNES se utilizó el kit espectrofotométrico Randox NEFA Fa-115 (Randox®, UK). Para estimar la concentración de β-hidroxibutirato se utilizó el kit Ranbut RB 1008 (Randox®, UK). Para TG se utilizó el kit Triglycerides (Biosystems®, España). Por su parte, las determinaciones espectrofotométricas se realizaron utilizando un espectrofotómetro Genesys UV-VIS 10S Thermoscientific. Para las muestras de hígado se determinaron las concentraciones de carnitina total, carnitina libre, acilcarnitina y triglicéridos utilizando métodos descritos previamente (Prieto et al. 2006) y colina-fosfolípidos (Galvis et al. 2016). Por último, se estimó el balance de energía neta de lactancia (ENL), el cambio de peso y los balances de proteína metabolizable (PM) y de proteína degradable en rumen (PDR) utilizando el software (NRC 2001).
Análisis estadístico
Se realizaron análisis de medidas repetidas en el tiempo utilizando el PROC MIXED del paquete estadístico SAS® (SAS® 1988). Se modelaron las estructuras de covarianza: simétrica compuesta y auto-regresiva de primer orden. Según el criterio de información de Akaike, la estructura de covarianza más adecuada para todas las variables fue la auto regresiva de primer orden. El número de partos, la producción de leche corregida (305d, 2x, EM) en la lactancia anterior y la condición corporal al inicio del experimento fueron involucrados como covariables. La comparación de medias entre tratamientos se realizó mediante la prueba LSD. Se aceptaron diferencias estadísticas cuando p < 0,05. El modelo estadístico incluyó los efectos fijos de: tratamiento, periodo de medición y la interacción entre el tratamiento y el periodo de medición.
Yhijk =U +β1 +β2 +β3 +Th +I(T) +Pj+T*Phj+E(hijk)
Dónde:
Yijk = Variable de respuesta del k-ésimo animal en el h-ésimo tratamiento dentro del j-ésimo periodo de medición. U = Media general; β1 = Efecto de la covariable 1; β3 = Efecto de la covariable 2; β3 = Efecto de la covariable 3; Th = Efecto del h-ésimo tratamiento (1-n); I(T). = Efecto del i-ésimo individuo anidado en el tratamiento (1-n); Pj. = Efecto del j-ésimo periodo de medición (1-n); T*Phj. = Efecto de la interacción entre el h-ésimo tratamiento y el j-ésimo periodo de medición; E(hijk) = Error experimental.
RESULTADOS
No se presentaron interacciones entre los tratamientos y los periodos de muestreo para ninguna de las variables analizadas. Ninguna de las covariables incluidas tuvo efecto significativo (p > 0.05).
En la Tabla 4 se presentan los indicadores metabólicos y el balance nutricional para cada uno de los tratamientos.
TABLA 4 Medias por tratamiento para los indicadores metabólicos y el balance nutricional en vacas Holstein sometidas a diferentes niveles de inclusión de colina y metionina.
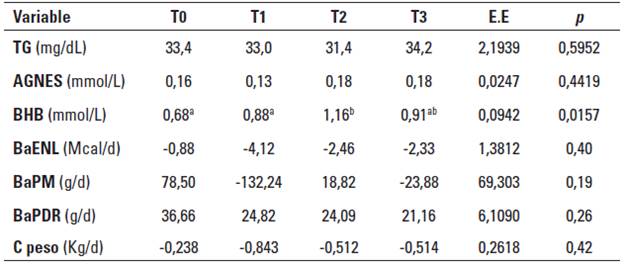
T0: control, T1: 20 g/d Aminoshure®, T2: 20 g/d de Aminoshure® + 60 g/d Reashure®, T3: 20 g/d de Aminoshure® + 120 g/d Reashure®, EE: máximo valor del error estándar de la medía, p: probabilidad de cometer el error Tipo I.
BaENL: balance de energía neta de lactancia, BaPM: balance de proteína metabolizable, BaPDR: balance de proteína degradable en rumen, C peso: cambio estimado de peso vivo.
La Tabla 5 presenta las los indicadores metabólicos y el balance nutricional por periodos de muestreo.
TABLA 5 Medias por periodos de muestreo para los indicadores metabólicos y el balance nutricional en vacas Holstein sometidas a diferentes niveles de inclusión de colina y metionina.
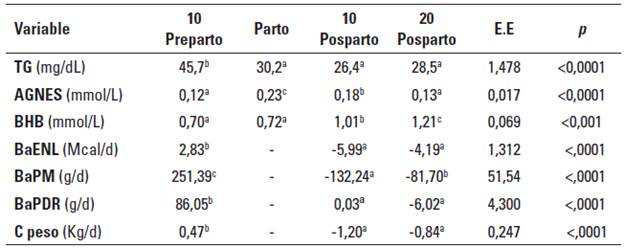
EE: máximo valor del error estándar de la medía, p: probabilidad de cometer el error Tipo I, BaENL: balance de energía neta de lactancia, BaPM: balance de proteína metabolizable, BaPDR: balance de proteína degradable en rumen, C peso: cambio estimado de peso vivo. Medias con letra distinta dentro de la misma fila son estadísticamente diferentes (p < 0.05).
La Tabla 6 describe el efecto del suministro de colina y metionina sobre las concentraciones hepáticas de TG, colina y carnitina para cada uno de los tratamientos.
TABLA 6 Medias por tratamiento de las concentraciones hepáticas de triglicéridos, colina y carnitina de vacas Holstein sometidas a diferentes niveles de inclusión de colina y metionina.
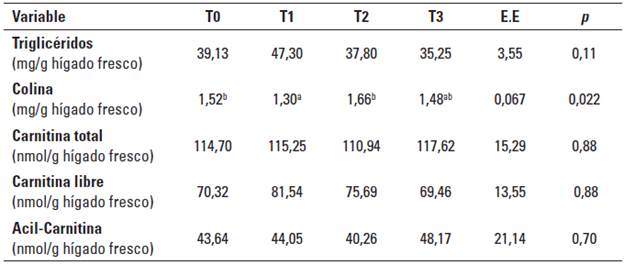
T0: control, T1: 20 g/d Aminoshure®, T2: 20 g/d de Aminoshure® + 60 g/d Reashure®, T3: 20 g/d de Aminoshure® + 120 g/d Reashure®, EE: máximo valor del error estándar de la medía, p: probabilidad de cometer el error Tipo I. Medias con letra distinta dentro de la misma fila son estadísticamente diferentes (p < 0.05).
La Tabla 7 muestra las concentraciones hepáticas de TG, colina y las diferentes formas de carnitina por periodos de muestreo. No se presentaron diferencias estadísticamente significativas entre periodos.
TABLA 7 Medias por periodo de muestreo de las concentraciones hepáticas de triglicéridos, colina y carnitina de vacas Holstein sometidas a diferentes niveles de inclusión de metionina y colina.

Medias con letra distinta dentro de la misma fila son estadísticamente diferentes (p < 0.05). EE: máximo valor del error estándar de la medía, p: probabilidad de cometer el error Tipo I.
DISCUSIÓN
Los tratamientos no tuvieron efecto sobre las concentraciones plasmáticas de AGNES, estos resultados concuerdan con los de investigaciones previas (Davidson et al. 2008; Hartwell et al 2000; Guretzky et al 2006; Leiva et al. 2015; Piepenbrink y Overton 2003); adicionalmente, como también se puede observar en la Tabla 4, los tratamientos no afectaron las concentraciones plasmáticas de TG. Solo se encontraron diferencias respecto a la concentración plasmática de BHB, donde el tratamiento control (T0) y el tratamiento que solo recibió suplementación con metionina (T1) presentaron valores significativamente más bajos. La explicación de estos resultados no radica en el balance nutricional; como se observa en la Tabla 4, los tratamientos no presentaron diferencias significativas respecto a los balances de ENL, PM y PDR.
Como se observa en la Tabla 5, los valores de AGNES alcanzaron su valor máximo el día del parto, esto debido al efecto li-político inducido por el cortisol (Weber et al. 2013), sin embargo, los niveles de AGNES decrecieron paulatinamente hasta el día 20 posparto a valores similares a los encontrados al día 10 preparto. Entre tanto, la variación en los TG plasmáticos presentó concordancia entre el balance energético preparto y posparto, con disminución significativa durante el posparto, lo que indica que el cambio en la disponibilidad de nutrientes tiene efectos importantes sobre la síntesis y exportación de TG desde el hígado.
Por su parte, la concentración hepática de TG no se afectó por la suplementación con metionina y colina (Tabla 6). Sin embargo, se observó una leve tendencia (p = 0,11) a su disminución en el tratamiento con dosis altas de colina (T3). En este sentido, la concentración de TG estuvo acorde con lo reportado por otros autores (Gross et al. 2013; Kalaitzakis et al. 2007; Starke et al. 2010) y en un rango de clasificación bajo según Kalaitzakis et al. (2007). Los valores reducidos de TG hallados corresponden con la demanda metabólica a la que estuvieron sometidos los animales, pues mientras que en la presente investigación se encontraron valores de AGNES entre 0,12 y 0,23 mM a lo largo de los periodos de muestreo (Tabla 5), otros autores reportaron que los valores de AGNES tuvieron una alta correlación (0,616; p < 0.05) con el grado de infiltración de triglicéridos hepáticos y solo observaron algún grado de lipidosis con valores de AGNES superiores a 0,25 mM (Kalaitzakis et al. 2007). Por su parte, Gross et al. (2013) reportaron valores de AGNES de 0,23 y 0,78 mM para las semanas -1 preparto y + 1 posparto, respectivamente, que se correspondieron con cambios significativos en la concentración de TG hepáticos entre la semana -1 preparto (25 mg/g) y la semana + 1 posparto (57 mg/g).
De otra parte, Roche et al. (2015) reportaron que en vacas en pastoreo el grado de condición corporal en el preparto y el nivel de alimentación (75, 100 y 125% de los requerimientos) hasta el primer mes posparto no tuvieron efecto sobre la concentración de TG hepáticos durante las 4 primeras semanas. Bajo las condiciones de pastoreo como las descritas por Roche et al. (2015), y como aquellas en las que se llevó a cabo la presente investigación, es probable que se presenten pocas condiciones de riesgo para la lipidosis hepática debido al reducido reto productivo al que están sometidos los animales; el nivel de producción reportado por estos autores (entre 22,9 y 24,6 kg/d) (Roche et al. 2015) es bastante similar al encontrado en la presente investigación.
La reducida respuesta en la concentración de TG hepáticos a las dosis utilizadas de metionina y colina pudo deberse a que con bajos niveles de engrasamiento hepático estos nutrientes no son limitantes para la formación de moléculas transportadoras y su exportación a la sangre, y por lo tanto, la suplementación no originó cambios en las concentraciones hepáticas y plasmáticas de TG. A su vez, la disminución en los triglicéridos plasmáticos después del parto no obedeció a deficiencias en las moléculas transportadoras, sino a una disminución significativa en su síntesis hepática debido al balance nutricional negativo durante este periodo, lo cual se evidenció porque no hubo cambios significativos en la concentración de TG hepáticos entre periodos de muestreo (Tabla 7). De este modo, durante la fase de balance nutricional positivo (preparto) aumentó significativamente la síntesis hepática de TG, los cuales no tuvieron limitantes para ser exportados a la sangre y por ende su nivel plasmático fue más alto durante este periodo, lo que permitió que no aumentaran significativamente las concentraciones hepáticas de TG. En el posparto, debido a que no hubo elevaciones dramáticas en las concentraciones de AGNES, su captura hepática no excedió su capacidad de oxidación, por el contrario, durante este periodo se observaron aumentos significativos en BHB, de tal modo que la disponibilidad de AGNES para reesterificación fue mínima, lo cual, sumado a la disminución significativa de la síntesis de novo de ácidos grasos, debido al balance nutricional negativo, posiblemente disminuyó significativamente la síntesis hepática de TG, lo que condujo a una disminución en su exportación a la sangre, que a su vez se notó en una disminución significativa en los TG plasmáticos, lo que permitió que no disminuyeran las concentraciones hepáticas de TG. Por su parte, otros autores reportaron que la suplementación con 15 g/d de colina protegida de la degradación ruminal (60g/d Reashure®) fue efectiva para prevenir la lipidosis inducida por restricción alimenticia (Cooke et al. 2007). A su vez, en el estudio de Zom et al. (2011) la suplementación con 14,4 g/d de colina protegida de la degradación ruminal (60g/d Reashure®) a vacas durante el periodo de transición a la lactancia solo consiguió disminuir la concentración de TG hepáticos en la semana 1 y 4 posparto, cuando las vacas no suplementadas presentaron concentraciones de 75 y 60 mg/g, respectivamente; por el contrario, en la semana -3 preparto y 6 posparto, cuando las vacas presentaron concentraciones de 27 y 40 mg/g, respectivamente, la suplementación fue ineficaz (Zom et al. 2011). Estos resultados ponen de manifiesto que la suplementación con colina muestra ser efectiva en condiciones de alto riesgo de lipidosis hepática, por el contario, a bajas concentraciones de TG hepáticos no existen limitantes para su exportación y por lo tanto, no se encuentra respuesta a la suplementación con colina.
Adicionalmente, se pudo evidenciar que los animales suplementados simultáneamente con metionina y colina presentaron valores más altos de BHB, lo que indica una mayor oxidación de ácidos grasos. La explicación de estos resultados podría ligarse a una mayor disponibilidad de carnitina para la incorporación de ácidos grasos a la P oxidación mitocondrial. En este sentido, es conocida la participación de metionina y colina como donantes principales de grupos metilos (Emmanuel y Kennelly 1984) y la importancia de estos en la síntesis de carnitina (Daily y Sachan 1995), por consiguiente, podría esperarse que el aumento en la disponibilidad de estos precursores aumentara su síntesis. Las concentraciones hepáticas de las diferentes formas de carnitina en los diferentes tratamientos (Tabla 6) estuvieron en el rango reportado por otros autores (Carlson et al. 2006; Carlson et al. 2007) en vacas que recibieron dosis bajas de L-carnitina (6 g/d); sin embargo, en la presente investigación no se presentaron diferencias significativas entre tratamientos. Ahora bien, la interacción entre colina y carnitina es poco conocida en bovinos, Daily y Sachan (1995) encontraron que en ratas y cobayos las concentraciones plasmáticas de BHB se incrementaron con la suplementación con colina, lo que fue interpretado como un indicador de una mayor capacidad de oxidación de ácidos grasos como consecuencia de una mayor disponibilidad de carnitina. Estos resultados sugieren que efectivamente en la presente investigación la suplementación con colina pudo haber aumentado la capacidad de oxidación de ácidos grasos, lo que se evidenció a través del aumento en las concentraciones de BHB, sin que necesariamente se elevaran las concentraciones hepáticas de carnitina, posiblemente debido a que el único tejido que realiza depósitos significativos de carnitina es el musculo, como lo demuestran las concentraciones significativamente más altas en éste tejido en bovinos, en los cuales, los reportes indican que las concentraciones de carnitina en músculo son entre 7 y 8 veces mayores a las encontradas en hígado (Carlson et al. 2007).
Finalmente, la suplementación con metionina y colina no consiguió aumentar las concentraciones hepáticas de colina; así, los valores encontrados (entre 1,30 y 1,66 mg/g, véase Tabla 6) son relativamente bajos comparados a los reportados (2,2 y 2,5 mg/g) por Sato et al. (2005). Las diferencias con estos autores pueden deberse a un mayor aporte de proteína metabolizable (PM) y colina derivadas de una ración totalmente mezclada. Por su parte, otros autores como Osorio et al. (2014) tampoco encontraron efecto de la suplementación con metionina sobre las concentraciones hepáticas de colina. En la presente investigación no se encontraron respuestas en las concentraciones hepáticas de colina, lo que pudo ser influenciado por el balance de proteína metabolizable (PM) y de energía neta de lactancia (ENL) del tratamiento control (T0) respecto al tratamiento que solo recibió metionina (T1) (Tabla 34). De esta forma, mientras que los tratamientos T2 y T3 presentaron balances de ENL y PM numéricamente más cercanos entre sí, el tratamiento control presentó un balance de ENL aproximadamente 2,5 veces superior con respecto a T2 y T3, y aún más, 4,7 veces superior respecto a T1. Respecto al balance de PM, T0 tuvo un balance positivo (78 g/d), mientras que en T1 se encontró el valor más bajo de todos los tratamientos (-132,24 g/d); bajo esta condición es probable que la utilización de metionina y colina como precursores de otras moléculas biológicamente importantes (carnitina, fosfatidilcolina) reste colina como para conseguir aumentar significativamente su concentración en hígado.
CONCLUSIÓN
Bajo concentraciones relativamente bajas de AGNES y TG hepáticos, la metionina y colina no son nutrientes limitantes para la exportación de TG desde el hígado a la sangre; por consiguiente, bajo esta condición no se observa lipidosis hepática y como consecuencia la suplementación con metionina y colina no ocasiona efectos significativos sobre las concentraciones hepáticas de TG.

