











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCTION
Hie tropical savannas of Northern South America located in the Orinoco River watershed cover 450,000 km2, between Eastern Colombia and Southern Venezuela. He well drained Eastern Plains of Colombia ("Llanos", 170,000 km2) extend to the right of the Meta River until it discharges in the Orinoco River. Approximately one-third of the area is high plain savannas and two thirds are rolling savannas. He Llanos are covered by tropical isohyperthermic savannas characterized by considerable plant and animal diversity. However, they provide low yields of low-quality forage growing on acid soils with deficient nutrient contents. In this context, Fisher et al. (1996) and CIAT-CIRAD (2001) made a detailed description of the region, while Rausch (2015) described the historical evolution of the area and noted that extensive cow calf systems constitute the traditional use of these savannas. Over the last 30 years an increasing proportion of introduced sown tropical grasses have contributed to the intensification of cattle production systems. Cattle fattening constitutes the primary use of these relatively expensive sown pastures. Nevertheless, the rolling savannas supported by superficial soils cannot generally be tilled. Beef breeding herds are mainly restricted to native savannas and occasional use of sown pastures receiving low external inputs (Rivas et al. 2013; Ramírez-Restrepo and Vera 2019; Ramírez-Restrepo and Vera-Infanzón 2019; Ramírez-Restrepo et al. 2019a).
Cow-calf extensive systems in the tropical savannas are notoriously variable in year-to-year reproductive and overall performance indices (Kleinheisterkmap and Habich 1985; O'Rourke 1986; O'Rourke et al. 1991; Cerón et al. 2017) due to nutritional and management limitations (Lascano 1991; Pereira et al. 2003; Ezenwa et al. 2006), and a negative correlation between pregnancy tests in consecutive years was reported by Kleinheisterkamp and Habich (1985). The above constraints make research particularly difficult due to variable results, and require a considerable number of animals over extended periods (O'Rourke 1986; O'Rourke and Howitt 1986; Ramírez-Restrepo and Vera-Infanzón 2019). Therefore, it is not surprising that the amount of research on extensive breeding systems is very restricted in the Llanos (Stonaker et al. 1976; Ramírez-Restrepo et al. 2019a).
The use of grasses with low nutrient requirements such as Brachiaria (syn. Urochloa) decumbens (Bd; signal grass;), B. brizantha, and B. humidicola (Bh; cv Llanero) has increased over the last decades in the region and they may improve female reproductive performance (Vera et al. 1993, 2002; Vera and Ramírez-Restrepo 2017), contribute to cattle fattening (Ramírez-Restrepo and Vera 2019); and/or maintain cattle that are of low priority for graziers such as young animals, particularly when Bh is used (Vera and Hoyos 2019). B. humidicola cv. Llanero (accession CIAT 679) is reputedly a low-quality forage when grazed by young stock (Hoyos and Lascano 1988), but cows and older steers can realize acceptable compensatory liveweight gains (LWGs; Ramírez-Restrepo and Vera 2019). Additionally, persistent Bh is associated with its biological-nitriication-inhibition (BNI) capacity, inhibition of soil-nitriier activity, and suppression of nitrous oxide gas emissions (Subbarao et al. 2017). These indings suggest that there is considerable latitude in managing sown pastures for beef production in integrated ranching approaches.
Nevertheless, knowledge of the mechanisms that promote heifers' body growth and link it to lifetime breeding efficiency in extensive tropical cow-calf systems is limited (Wathes et al. 2014), but it is essential to develop sustainable production systems. Poppi et al. (2018) recently remarked the continuing importance of research on the effects of growth rate on the performance of cattle grazing tropical pastures, and noted the inancial risks involved in different growth paths.
To the authors knowledge, there are no reports regarding lifetime LW yields of unsupplemented breeding herds composed of Brahman (Bos indicus) and crossbred [Brahman x San Martinero (native; B. taurus)] cattle grazed on these Brachiaria spp. In this context, we hypothesized that early growth patterns would affect lifetime production of beef commercial cow-calf systems raised on B. decumbens and B. humidicola pastures subject to extensive management in four complementary, long-term trials.
MATERIALS AND METHODS
Location
The experiments reported were conducted over a nine-years period at Carimagua Research Center (CRC; 4°36'44.6"N, 74°08'42.2"W) located in the well-drained tropical savannas (Vera, 1991; Vera et al. 1993, 2002) of the Colombian Meta Department. Their results apply to the savanna municipalities of Puerto López and Puerto Gaitán, 3° 55' to 4° 20' North latitude, and 72° 1' to 72° 55 West longitude, where other complementary studies were conducted over 20 years (Vera and Hoyos 2019).
Animals
In all cases, Brahman heifers used (n = 149) born on native savanna were reared and weaned at an average age of 270 days (140 kg LW), and allowed to reach 170 kg LW before randomly entering the experiments. Across years, LWG (n = 5000), reproductive (n = 550) and mortality (n = 8) events were recorded. They were used to construct the dataset presently used to assess LW performance during the "Growing" (i.e. nine-36 months of age) and "Breeding" (i.e. cows plus weaned calves; Holroyd and McGowan 2014) phases in terms of beef output per head, hectare (ha), and calendar year. Continuous mating was used in three experiments. Although calendar years do not necessarily coincide with reproductive events, they constitute the common unit of time used by ranchers for budgeting purposes.
Ethical considerations
Following national animal husbandry regulations, care of the experimental animals was supervised daily at CRC by registered Colombian Doctors of Veterinary Medicine (DVM) including the second author.
Cattle had always free access to fresh water and to a complete commercial mineral supplement containing as-fed 17.5%, 26.9%, 8.0%, 13.7%, 2.0%, 0.104%, 0.35%, 0.001% and 0.0076% of Na, Cl, P, Ca, S, Cu, Zn, Co and I, respectively (Ramirez-Restrepo and Vera 2019).
Measurements
Heifers were weighed at monthly intervals during the Growing phase. During the Breeding phase, they were subject to continuous mating using regularly tested purebred Brahman bulls, which were rotated every three months from a pool of ~ 20 bulls. The pregnancy status (rectal palpation) and LWs of cows were recorded four times per year. Birth weights of newborn and sequential identification were recorded in the field within 24 hours of calving. Suckling calves were weaned at approximately 270 days of age. At the end of experimentation, cows joined the main commercial CRC herd, and if empty, after two consecutive pregnancy diagnosis, all infertile-dry cows were subjected to a fattening period for achieving slaughter LWs (Ramirez-Restrepo and Vera 2019).
Experiment 1
The trial consisted of three growth treatments using individual animals as observation units. Stocking rates (SRs) of 2.72, 1.94 and 1.45 animals/ha were used to achieve low (L), medium (M) and high (H) rates of growth (Table 1). The aim was to mimic the range of weight gains observed in the commercial herd in much larger savanna paddocks at CRC as well as in a range of private ranches (Vera and Hoyos 2019). A total of 51 heifers [171 ± 3 kg initial LW (ILW); 441 ± 10 days of age] were used until reaching the target breeding weight of 270 kg while grazing three contemporary and adjacent Bh cv Llanero paddocks. Further details on pastures and their attributes are given below. The subsequent individual reproductive performance during the following six-year period was monitored while subject to common grazing on another contemporary Bh pasture stocked at 1.5 animals/ha. Individual reproductive performance and time and LWs to reach different reproductive events were analyzed by Vera (1991) and Vera et al. (1993, 2002), and are therefore not included here. The present paper reports yearly LWGs of the heifers-cows and that of the cow-calf pairs as well as their production per ha. These animals constituted temporal replicate one.
TABLE 1 Least squares means (LSM ± SEM) values of daily liveweight gains (DLWG, kg/head) during the Growing phase, in two temporal replicates at Carimagua Research Station, Colombia.
| Growth treatment | Replicate | DLWG, period 1 | Replicate | DLWG, period 2 |
|---|---|---|---|---|
| L | 1 | 0.061 ± 0.014 | 1 | 0.220 ± 0.018 |
| M | 1 | 0.197 ± 0.014 | 1 | 0.174 ± 0.015 |
| H | 1 | 0.251 ± 0.014 | 1 | 0.143 ± 0.018 |
| L1 | 2 | 0.108 ± 0.021 | 2 | 0.110 ± 0.021 |
| M1 | 2 | 0.192 ± 0.021 | 2 | 0.229 ± 0.021 |
| H1 | 2 | 0.203 ± 0.021 | 2 | 0.107 ± 0.021 |
| Overall LSM | 0.165 | 0.211 | ||
| RSME | 0.034 | 0.072 | ||
| Treatment p | < 0.0001 | < 0.0001 |
Experiments 2 and 3
Experiment 2 constituted a temporal replicate of Experiment 1 during the Growing period using the same treatments (L1, M1, H1; Table 1), the same SRs and the same Bh paddocks, with a total of 42 animals (165 ± 2 kg ILW, 410 ± 17 days of age). Upon reaching the target weight of 270 kg, the three groups were equally split into two subsequent grazing treatments during the Breeding phase (low, l, and high, h, respectively) that lasted six years. his provided a 3 x 2 factorial combination of before and after treatments (H1 h, H1 l; M1 h; M1 l; L1 h, and L1 l, respectively).
In the Breeding phase, the low treatment animals (i.e. H1 l, M1 l; L1 l) remained on a common Bh pasture, whereas the high (i.e. H1h, M1h; L1h) counterparts grazed a contemporary, well managed, Bd pasture stocked at 1.5 animals/ha and managed as described below, in order to achieve contrasting LWGs during that phase. he design therefore mimics studies on compensatory growth whose importance for tropical cattle was recently reviewed by Poppi et al. (2018).
The first period corresponds to the application of the initial low (L), medium (M) and high (H) rates of growth to maiden heifers on Brachiaria humidicola. The second period corresponds to the same heifers with less than 2.5 months of pregnancy during the mating season. RSME is the root mean square error.
Experiment 3 consisted of 33 heifers (271 ± 16 kg; 806 ± 138 of age) that were raised and maintained as a positive control (Vera et al. 2002) on a well-managed Bd pasture (see below), adjacent to the other experimental paddocks, and stocked at I. 5 animals/ha. Their Growing phase was very short, and their data is not included in the joint analysis of the growing phase. Overall, Experiments 1, 2 and 3 overlapped for the duration of the experimental period except for the initial year.
Experiment 4
Briefly, the research (Ramirez-Restrepo and Vera 2019) analyzed the LW of cull, non-pregnant-dry cows during three consecutive years (n = 9 per year; 309 ± 15 kg ILW, 8.7 ± 0.54 years; 296 ± 17 kg ILW, 10.9 ± 0.61 years and 314 ± 15 kg ILW, II. 4 ± 0.54 years ofage, respectively). They continuously grazed a Bh pasture stocked at 1.5 animals/ha and close to the rest of the remaining experimental paddocks, unencumbered by reproductive events during 160-180 days of the rainy season (April-November). The experiment overlapped the last three years of Experiments 1, 2 and 3, and it provides evidence on the LWGs of the last productive stage of female cattle as cull cows.
Pasture management and attributes
All four experiments were located in paddocks contiguous to one another at CRC. Pastures were established on the same soil type and topography, with minimal fertilization (20 kg/ha of P and K each) at least two years before the beginning of Experiment 1. Thereafter, paddocks were subject to the same strategy of minimal fertilization using 20 kg/ha of P every three years.
Average forage dry matter (DM) on offer in the B. humidicola cv. Llanero (CIAT 679) pastures were 3,584, 1,216 and 111 kg/ha for the low, medium and high SRs, respectively, with insignificant differences between the wet and dry seasons. Nevertheless, there was a large difference between seasons in the contents of green leaf, which amounted to 10.0-11.1 g/100 g of pasture during the dry period, and 20-30 g/100 g of forage over the length of the rainy season. These differences were associated with crude protein (CP) contents of70-80 g/100 g DM and 80-130 g/100 g DM during the dry and rainy seasons, respectively. The in vitro DM digestibility (IVDMD; g/100 g DM) varied from a high value of 58 to 65 at the highest SR in the dry season, to values between 56 and 62 at the lowest SR, in rainy season.
Mean forage DM and leaf CP content in the B. decumbens pasture offered in the dry season were 3,350 kg/ha and 9 g/100 DM, respectively, while in the rainy season corresponding values were 2,600 kg/ha and 14 g/100 DM. The IVDMD was 61 and 69 g/100 g DM for the dry and wet seasons respectively.
Statistical analyses
he data was rearranged on the basis of calendar years to allow computation of yearly LWGs. Graphical analyses of the data per year, per period and per phase (Growing and Breeding, respectively) were carried out to determine the distributional properties of the data, including calculation of boxplots, means, medians, and outliers using Proc Boxplot and Proc Univariate (SAS® 2009).
Each of the experiments was analyzed separately, followed by joint analyses as described below. Subsequently, the Growing phase of Experiments 1 and 2 was split into two periods (Table 1). Period 1 coincides with the duration of the initial treatments, lasting 100-417 days depending upon the initial growth treatment, whereas period 2 extends from the end of initial treatments until the heifers reached 2-3 months of pregnancy based on the assumption that they continued to grow and that stage of pregnancy did not significantly influence weight gains. The statistical model included the effects of the growth treatments imposed on heifers and cows, replicates or periods (fixed effects), and animals (random effect), plus all the corresponding interactions (as applicable). Non-significant effects where discarded in the final analyses. Effect sizes [partial omega-squared statistic (ω 2 ) were calculated (alpha set to 0.10 for confidence coefficients, with the F test set at 0.05)].
Consecutive LWs were fitted to generalizable linear mixed models with SAS (2009) Proc GLIMMIX using year, treatment and the animal x year interaction; given that animals were treated as random variable the interaction with year is also random. Production per ha during the breeding phase refers to the yearly change in cows' LW plus that of their weaned calves (Holroyd and McGowan 2014).
Scatter plots of lifetime weight gains per head and per ha were subjected to locally weighted smoothing using SAS® (2009) Proc LOESS with year as regressor variable to verify graphically the separation between treatments.
All values are presented as least squares means (LSM). Precise model p-values are shown when available, standard error of the means (SEM) were determined by bootstrapping for important parameters (Chernick 2008), and confidence intervals were calculated since the emphasis of all analyses is on confidence intervals rather than p values (Halsey et al. 2015; Wassersteina and Lazar 2016).
RESULTS
Growing phase
Growth treatments significantly (p < 0.0001) influenced average yearly LWG/ head in Period 1. Table 1 shows the re-peatable performance of the two replicates, and consistent differences between treatments. When one half of the animals of replicate 2 were allowed to graze the better quality Bd pasture (in Period 2), a large and significant compensatory growth occurred (Table 2) as soon as the pasture restriction was removed. The effect sizes yielded low to moderate values (0.15-.022) of the ω 2 statistic, and were interpreted according to Keppel and Wickens (2004).
TABLE 2 Treatment mean live weight gains (LWGs, kg/ha/year), averaged over 9 years, and mean annual LWGs during consecutive years averaged across experimental treatments. Data are least square means with their respective confidence intervals between parentheses.
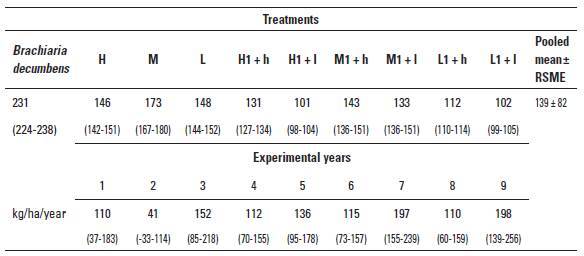
Low (L), medium (M) and high (H) rates of growth during the Growth phase on B. humidicola followed during the Breeding phase by grazing on B. humidicola (low, l) or B. decumbens (high, h) pastures. RSME: root mean square error.
Growth rates in the second temporal replicate for the H1, M1 and L1 treatments over the experimental period of 377, 497 and 484 days, showed a significant (p < 0.01, Table 1) and generally large advantage of animals' subject to the H1 and M1 treatment during period 1, and an even larger advantage when subject to the h treatment during period 2, relative to animals in either the L1 or l treatments, respectively (Tables 1 and 2).
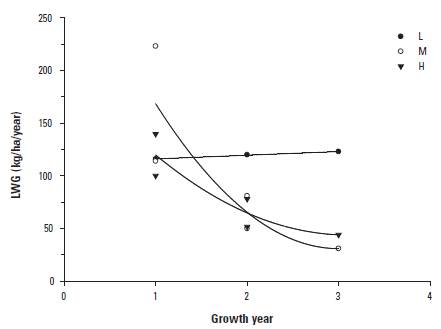
The previous results contrast with that of annual LWG/ha that showed different (p < 0.01) temporal behaviors depending upon the growth treatment (Figure 1). There were significant (p < 0.001) differences in yearly LWG/ha between treatments applied during the Growing phase. Pooled means and their confidence intervals for all H, M, and L treatments were 68 (66-70), 101 (98-103), and 117 (115-119) kg/ha/year, respectively. Figure 1 depicts the temporal trends of LW, and shows an expected spurt of compensatory growth for L in the second half of the Growing phase. As indicated previously, the Growing phase on Bd (Experiment 3) was very short, with individual gains of 0.419 ± 0.071 kg/day, equivalent to 229 kg/ha/year.
Breeding phase
Cow's LWGs/head and per year during the breeding phase were low, very variable, and did not differ between any of the treatments (p > 0.05), with an average gain of 12.9 ± 31.2 kg/head/year (range 4.6 to 15.0 kg). There was a significant year effect (p = 0.0043), with alternating low and high values of 49, 16, -10, 14, 1, -7, -2 kg/ha from year 1 to 7 of the breeding phase, respectively.
Liveweight gains per ha and year during the breeding phase were composed almost exclusively of the calves' weaning weights. he overall mean gain across all treatments was 147 ± 90 kg/ha/year. Years in the Breeding phase had a significant effect on performance per ha (p = 0.031), characterized by alternating high and low production in consecutive years. he cyclical behavior of animal performance anticipated the behavior of lifetime performance (Table 2). There were significant and practically important differences between treatments in the two replicates examined (Figure 2), including the 3 x 2 factorial combination of Experiment 2. Twofold important differences included that between animals on Bd versus the rest, and that between H1 h and M1 h and the low treatments. The ω2 statistic was 0.41, and the breeding treatment x year interaction was NS (p > 0.05).
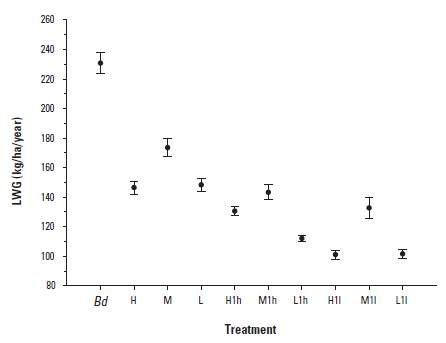
FIGURE 2 Mean lifetime liveweight gain (LWG) in all treatments. Data consists of jackknifed means and their standard errors. Brachiaria decumbens (Bd), H, M and L correspond to high, medium and low treatments, lower case letters identify high, medium and low treatments applied during the breeding season only, all on B. humidicola pastures.
Lifetime performance
The sum of yearly LWGs per ha over nine consecutive years of experimentation was taken as an indication of lifetime performance of the breeding system. Averaged across years, LWG was 139 ± 82 kg/ha/year, and the differences between treatments (Table 2) paralleled those shown in Figure 2. Similarly, to what was observed in the Growing and Breeding phases, there was a regular alternation of high and low performance in successive years (Table 2). Nevertheless, the LOESS analysis revealed a consistent, and less variable, superiority of the Bd treatment over all of the pooled H and L treatments (with means of232 ± 75, 135 ± 78 and 125 ± 85 kg/ha/year, respectively) with a clear discrimination between Bd and the rest.
The growth of non-gravid, dry, cull cows on Bh (Experiment 4) measured on three consecutive years and standardized to 180 days of the rainy season averaged 0.458 ± 0.094 kg/head/day. This is equivalent to 82 ± 4.8 kg/head and 123 ± 15 kg/ha/year over the experimental period.
DISCUSSION
The Colombian Llanos have a long and rich history of extensive beef cattle production which constitutes the main land use system of the region (Huertas-Ramírez and Huertas-Herrera 2015; Rausch 2015). However, in the context of climate variability (Ramírez-Restrepo and Charmley 2015; Tapasco etal. 2015), there is increasing pressure for the primary sector to achieve sustainable intensification, preserve ecosystem services, and meet national and global food demands (Boron et al. 2016; Norton 2017). In parallel, the gradual introduction of crops and plantations in the region increases the competition for, and the value of, tillable savanna lands. Brachiaria humidicola is a popular species throughout the lowland tropics, although its nutritive value is consistently low across a variety of environments and independently of SR (Ezenwa et al. 2006; Pereira et al. 2009). Ramírez-Restrepo and Vera (2019) confirmed the low to modest nutritive values of both grass species using fecal analyses across four classes of beef cattle in a contemporary, long-term experiment located next to the present ones.
A large (2,000 ha) five-year experiment reported by Rivera (1988) studied the performance of breeding herds and growth of heifers on two types of high-plains native savannas differing in soil texture. Rivera (1988) found heifers LWGs of 0.221 ± 0.024 kg/head/day (range 0.192-0.229) in five large (250 ha each) medium texture savanna paddocks next to our experiments, and 0.113 ± 0.020 kg/head/day (range 0.093-0.115) on three sandier savanna paddocks. hus, the treatments tested in Experiments 1 and 2 were successful in replicating savanna-based LWGs on smaller, better controlled paddocks of Bh since they stranded well the individual performance reported by Rivera (1988) and Vera and Hoyos (2019). he latter results were obtained in a variety of highly representative native savannas stocked at much lower rates than that possible on Bh (0.15-0.25 versus 1-1.5 head/ha, respectively), and contrast markedly with the performance observed on Bd (stocked at 1.5 head/ha; Experiment 3).
Growth treatments had a modest influence on overall weight gains per head and per ha during the Growing period, as revealed by the statistical analysis and the ω2 statistic, in part due to the expected high variability observed. Nevertheless, low to medium effect sizes (ω2) are generally or more interest than larger, evident, values (Keppel and Wickens 2004). Contrary to the initial hypothesis, differences in early growth rate of heifers were generally not carried over to lifetime LWGs per head with the exception of the two extreme treatments (Bd and L1 l, respectively). he relatively large value of ω2 and the loess analyses coincided in the superiority of the long-term Bd treatment. Within those limits, the data demonstrates a considerable range of alternative growth rates that resulted in similar, but variable, lifetime performance, but the year-to-year cyclical performance was large. his indicates the need for long-term evaluation in these extensive and highly variable production systems. As Roche et al. (2015) pointed out, the manager's challenge is to assess the economics of different paths in light of the existing feed resources, environmental constraints, and prices of inputs and outputs.
There was an overall superiority of growth and breeding performance in the Bd treatment that was higher than that of any other combination of low and high treatments, and that is consistent with its higher nutritive value (Lascano and Euclides 1996). Growth rates of replacement heifers in moderately intensive systems are considered to impact lifetime productivity (Patterson et al. 2003) and financial returns. Nevertheless, this is not so clear when prospective bio-economic experiments have been carried out in the context of extensive grazing systems (Roche et al. 2015). It would generally be expected that increasing the growth rate of replacement heifers would lead to improvements in systems performance and this may certainly be the case in more intensive grazing systems (Wathes et al. 2014). his expectation was not fulfilled in the present experiments unless breeding heifers and cows were allowed to graze full time on Bd. It is clear from Experiments 1 and 2 that the growth rates of heifers were low, even in the best before-and-after treatments. he review by Funston et al. (2012) suggested that it is unlikely that high heifer growth rates would result in significantly improved beef output of extensive systems. It is therefore hypothesized that this may be one of the reasons behind the low priority assigned by ranchers to growing replacement females on costly sown pastures requiring higher physical and management inputs. This claim would argue against the logic of raising heifers on pastures such as Bd (Experiment 3), usually assigned to fattening steers that provide higher short-term financial returns. This view coincides with that expressed by Poppi et al. (2018) to the effect that under some conditions, a minimalist approach to feeding grazing cattle may be economically optimal; this argument may explain the popularity of low input tropical pastures among ranchers in some regions.
As expected, growth rates during the growing phase generally decreased with time (Figure 1) as animals asymptotically approached the mature weights allowed for by the respective nutritional treatments (Vera et al. 1993), with the exception of the L growth rates. The late growth spurt of the L growth treatment during the Growing phase, allowed for by the necessary early removal of these animals from their treatment, resembles the strategy tested by Lynch et al. (1997) who maintained heifers with low LWGs during the initial two thirds of the development period, followed by a high growth rate in the last third. However, Lynch et al. (1997) did not find significant differences in reproductive performance compared to heifers that grew at a constant rate. Their findings are similar to those of Vera (1991) and Vera et al. (1993) that used much lower growth rates compared to Lynch et al. (1997). Nevertheless, contrary to Lynch et al. (1997), Vera et al. (1993) found also a negative carry-over effect of low early growth rate until the 4th calving, probably due to the limited opportunity afforded by Bh to realize compensatory LWGs in mature, breeding cows.
Long inter-calving intervals, together with continuous mating, result in large year-to-year variations in extensive beef production of tropical breeding herds (O'Rourke et al. 1991; Fordyce et al. 2013, 2014; Cerón et al. 2017). In parallel, beef output should also show year-to-year variations, as demonstrated here, but the magnitude of this phenomenon has not been previously reported. The repetitive two-year cycles of low production followed by higher yields in the following year (Table 2) is probably associated with the alternating low and high reproductive rates reported by Kleinheisterkamp and Habich (1985) for ranches in the region. Regardless of the magnitude of the year-to-year variation in breeding performance, our results reiterate the need to study extensive tropical cow-calf enterprises over the long term. Admittedly, given current trends in financing cattle, this would be difficult to achieve, and that can alternatively be approached via simulation studies (Pullet and Martin 2017; Ramírez-Restrepo and Vera 2019; Ramírez-Restrepo and Vera-Infanzón 2019; Ramirez-Restrepo et al 2019a, 2019b, 2020).
The well-established curvilinear relationships of SR with individual performance and with beef output per ha in steers (Mott 1961), that result in maximal output per ha with less than optimal individual gains, appear to extend to equivalent parameters in breeding herds as demonstrated here. Our M and M1 treatments of Experiments 1 and 2 out yielded the other growth rates. These findings do not appear to have been reported previously for breeding herds.
In comparison, the difference in lifetime production with animals raised and bred full time on Bd was large and practically important amounting to 70 kg/ha/ year relative to those animals grown on Bh and bred on Bd. The above differences would be larger if it is considered that heifers grown on Bh required between two and three years to reach first conception depending upon the severity of undernutrition, versus 18 months for those on Bd (Vera et al. 2002). Differences between these management strategies in deaths, lifetime production and voluntary culling have not been previously studied. Strategies that combine both pastures, particularly with heifers raised on Bh and then allowed to make compensatory gains on Bd offer a less demanding management alternative. On the contrary, strategies that keep females in marginal nutritional conditions are certain to result in low lifetime production as shown above. Depending upon economic conditions and farmers' preferences (Poppi et al. 2018), supplementation of cattle grazing these tropical pastures would be expected to increase their performance. Several studies conducted in the Brazilian Cerrado have unequivocally shown a positive response of fattening cattle grazing Brachiaria-based pastures to energy and protein supplements (da Silva-Marques et al. 2018a, 2018b; Neves et al. 2018). On the other hand, supplementation of tropical beef cows has been little studied, but da Silva et al. (2017) reported a small positive effect on weight losses in early lactation when Nellore (Bos indicus) cows, grazing Bd were supplemented with concentrates during the last 60 days of pregnancy; nevertheless, the effect of supplementation did not translate into reduced calving intervals.
The magnitude of the differences in cows' LW when exiting the breeding herd can be judged from the difference in the asymptotic LWs observed in the factorial experiment (range 332-387 kg) and that of 388-422 kg for cows grazing full time on Bd (Experiments 2 and 3; Vera et al. 2002). These exiting LWs would contribute to the final systems output of beef per ha and if the intent is to sell heavy cull cows, they impose different management strategies (Ramírez-Restrepo and Vera 2019). Hence, the present results clearly show that some flexibility is possible in managing replacement beef heifers and breeding cows in extensive systems, subject to low management inputs. Nevertheless, differences in outputs between some of the approaches tested here may pass unnoticed in extensive, low-management systems where records of animal outputs are infrequently kept (Wathes et al. 2014), and that are further compounded by between-years variability.
From a whole system perspective, the yields reported here should eventually include the final cows' LWs once they are culled from the herd as discussed previously, and in this case, animals bred and maintained on Bd would show an advantage compared to those coming from Bh. On the other hand, the results from Experiment 4 provide a baseline of what could be expected if thinner cull cows are partially fattened on a low input Bh pasture, as frequently observed on farm (Vera and Seré 1989).
Lastly, it should be noted that other important outputs of these systems such as potential carbon sequestration (Fisher et al. 1994; Mosquera et al. 2012; Ramírez-Restrepo et al. 2019a, 2020), methane emission dynamics (Ramírez-Restrepo and Vera 2019; Ramírez-Restrepo and Vera-Infanzón 2019), BNI inhibition (Subbarao et al. 2017), and other environmental impacts are not considered here, but they would need to be accounted for in the analyses of the whole enterprise. Similarly, much remains to be investigated in terms of the consequences of system intensification based on the combination of savanna lands with sown introduced pastures, on the conservation and biodiversity of the rangelands, and the overall management of increasingly complex systems (CIAT-CIRAD 2001; Belnap et al. 2012; Neilly et al. 2018). Finally, it can be inferred from the various alternatives discussed above that the dynamics of complex, extensive beef breeding systems with their numerous animal and pastures components, and management alternatives, could usefully be further analyzed with empirical simulation studies (Scanlan et al. 2013).
CONCLUSIONS
The performance of beef breeding herds on the low input pastures used in the present retrospective study was modest in keeping with the amount of external and management resources used, and showed large and significant variability between years and also between management strategies. With the exception of animals raised exclusively on Bd, beef output per year was markedly cyclical, such that relatively high outputs in one year were followed by lower production in the following year. This trend was most likely associated with alternating high and low reproductive rates previously reported from a large on-farm study (Kleinheisterkamp and Habich 1985). During the breeding years, significant and practically important differences were found between different LWG patterns associated with different growth and pasture strategies. All of the analyzes performed showed that the Bd system outyielded the rest. The final beef output of the various management strategies studied would be influenced by the cows' culling LW at the end of their useful breeding life, an issue briefly examined in the paper. A fuller, whole system analysis of these and other implications would require further retrospective studies using a dynamic system approach.
Implications
The present results offer an ample set of management alternatives to raise heifers and breeding cows in extensive tropical beef systems, and illustrate the tradeoffs involved in different management strategies that are influenced by the quality of the available forage resources. The difference between the Bd strategy and the rest reflect the differences between the two forage resources tested. Decision making regarding these systems should take a long-term view as physical and financial outcomes may vary widely over successive years, thereby demonstrating, the importance of long-term animal production records.

