











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
El cuerpo lúteo (CL) del bovino es la glándula transitoria productora de P4 (Shrestha et al. 2019), encargada del mantenimiento de la gestación (Senger 2012), cuya vida útil no solo está controlada por la hormona luteinizante (LH), que actúa como una hormona luteotrófica, sino también por la prostaglandina F 2 alfa (PGF2α) que termina la función y la presencia de CL (Meidan et al. 2017).
Existe controversia en torno a si un CL de conformación grande estaría asociado o no con concentraciones elevadas de P4 plasmáticas, ya que algunos investigadores así lo han establecido, al menos en los primeros días de gestación (Mann 2009), y se ha asociado la intensidad de la expresión del estro con el tamaño folicular, el volumen del CL y las concentraciones plasmáticas de progesterona (Oosthuizen et al. 2020; Rodrigues et al. 2018).
Se encontró que la relación entre el peso del CL y la concentración de progesterona en el plasma dependería de la condición en el balance energético del animal (Yung et al. 1996) o de su condición fisiológica, pues se hallaron relaciones directas en novillas y vacas lactantes, pero no en vacas no lactantes en un mismo estudio (Sartori et al. 2002). También se reportó una fuerte relación entre el peso del CL y los niveles plasmáticos de P4 en el día 5 de la fase lútea (Green et al. 2005). Sin embargo, en otros estudios se estableció que esta relación se pierde en el día 8 (Robinson et al. 2005).
El incremento en el número de células de la granulosa (CG) del folículo preovulatorio podría resultar en un mayor número de células luteales grandes, con un subsecuente incremento en el tamaño del CL e incremento en la secreción de P4 (Sartori et al. 2002), puesto que el nuevo CL se desarrolla a partir de las células foliculares restantes después de la expulsión del ovocito (Shrestha et al. 2019).
Las CG son células somáticas y se consideran el tipo de células ováricas más importantes que rodean, apoyan y nutren al ovocito en desarrollo y proporcionan un microambiente adecuado para su maduración (Edson et al. 2009); se han resaltado diversos pasos transcripcionales que las CG deben completar antes de la ovulación y se han identificado transcriptos asociados con el estado de competencia del ovocito (Gilbert 2011); cualquier alteración de la calidad de las CG y su capacidad de proliferación también puede tener efectos sobre el desarrollo del folículo y puede interrumpir la maduración de los ovocitos y, en consecuencia, dar lugar a un desarrollo embrionario deteriorado con resultados insatisfactorios, revisado por Khan et al. 2020.
En consecuencia, una mayor comprensión de los eventos fisiológicos que suceden en cada uno de los compartimientos foliculares en las diferentes etapas de foliculogénesis permitirá una mejora no solo en la intervención de esos procesos con fines productivos, sino que facilitará avances en los resultados de los procedimientos biotecnológicos aplicados a la reproducción de los animales domésticos de interés zootécnico, particularmente en bovinos de las razas RS (raza criolla colombiana en peligro de extinción) y Br (raza de gran participación en la ganadería colombiana), en las cuales a la fecha no se han estudiado.
La realización de investigaciones que contribuyan al conocimiento de la fisiología de los animales en el trópico es importante, ya que si bien existe abundante información referente a los procesos fisiológicos acontecidos durante el ciclo estral de la hembra bovina (Galina y Arthur 1990) y a numerosos factores que se relacionan con el desarrollo folicular, esta ha sido generada en países con condiciones medioambientales diferentes o con animales de otra composición racial. Muchos estudios in vitro sobre el crecimiento folicular se n enfocado en la replicación de las CG, mientras que los estudios in vivo, mediante el uso de ultra-sonografía, se han centrado en la expansión del antro folicular y su fluido (Rodgers e Irving-Rodgers 2010). Sin embargo, en la literatura consultada no se encontraron reportes en los cuales se estudie en las razas RS y Br el efecto del tamaño de los componentes foliculares sobre el desarrollo y función esteroidogénica del CL en vacas mantenidas bajo condiciones propias del bosque seco tropical.
Con este trabajo se buscó comparar en folículos preovulatorios de hembras bovinas de las razas Br y RS el desarrollo de diferentes componentes foliculares, tamaño del antro folicular, el grosor y el volumen ocupado por la capa de células de la granulosa y de la teca interna, y su posible relación con el desarrollo máximo del CL y la producción de P4.
MATERIALES Y MÉTODOS
El trabajo contó con la aprobación del comité de bioética de la Facultad de Medicina Veterinaria y de Zootecnia de la Universidad Nacional de Colombia y se desarrolló en el Centro Universitario Regional del Norte (CURDN) de la Universidad del Tolima, en condiciones de temperatura media de 28,2°C, precipitación anual promedio de 1,791 mm, humedad relativa: 71% y altitud entre 275-550 m s. n. m. y está clasificado como bosque seco tropical.
Se utilizaron 5 hembras de la raza RS y 5 de la raza Br con edad entre 3 y 5 años, que habían registrado por lo menos un parto como indicador de su fertilidad, clínicamente sanas y cíclicas, las cuales permanecieron en potreros establecidos en pasto Ángleton (Dichanthium aristatum) y contaron con sal mineralizada y agua a voluntad.
Para verificar su normal ciclicidad ovárica, a las 10 hembras se les realizó seguimiento ecográfico durante 2 ciclos estrales, mediante el uso de un equipo Aquila Pro, con sonda microconvex con frecuencia de 7,5 MHz y guía de aspiración folicular de Pie Medical® (http://www.piemedicalimaging.com/). Se practicó la técnica de registro de información descrita por Pierson y Ginther (1988). Las imágenes se grabaron en una computadora portátil con la ayuda de una tarjeta capturadora de video Pinnacle Studio MovieBox Ultimate 12®, para posteriores análisis. Se elaboraron los correspondientes mapas ováricos para establecer el momento de la identificación del cuerpo lúteo como evidencia de ovulación, en el lugar donde se encontraba el folículo ovulatorio.
Para establecer las concentraciones séricas de P4 durante los 2 ciclos estrales, se realizó colecta de muestras sanguíneas por venopunción de la vena caudal media con tubos estériles al vacío. El suero sanguíneo se obtuvo por centrifugación a 1500 rpm durante 10 minutos y los sueros obtenidos se congelaron a -20°C hasta su análisis. La técnica utilizada fue la de Elisa, se manejó kit comercial DS-EIA-Steroid-Progesterone, que emplea calibradores para una curva referencial, con concentraciones de 0, 1,5, 10, 25, 50 y 100 nm/l (0, 0,47, 3,14, 7,85, 15,7 y 31,4 ng/ml), procedimiento previamente estandarizado en el laboratorio de hormonas de la Facultad de Medicina Veterinaria y de Zootecnia de la Universidad Nacional de Colombia. Se siguió el protocolo descrito por el fabricante. El coeficiente de variación intraensayo fue de 3,6 y el de interensayo de 4,2.
Para facilitar la detección de estros, los animales fueron identificados mediante numeración con pintura en los flancos y se realizó por observación directa en los potreros que habitualmente pastoreaban, entre las 05:30 y las 06:00 h, entre las 09:30 y las 10:00, entre las 13:30 y las 14:00, entre las 17:30 y las 18:00 y entre las 21:30 y las 22:00, con la ayuda de un macho presente en el hato, con desviación lateral del prepucio.
A partir de la observación de la finalización de los signos de estro de cada uno de los celos, momento determinado por la desaparición del reflejo de inmovilidad al intento de monta, se realizó ecografía con el mismo equipo y transductor transrectal con frecuencia de 8,0 MHz cada 4 h hasta que se presentara la ovulación (ausencia del folículo preovulatorio), con el propósito de precisar el promedio del lapso transcurrido entre el inicio de los signos de celo y la ovulación en cada una de las hembras objeto de estudio. El tiempo de ovulación se definió como el tiempo de desaparición del folículo, menos 2 h (Walker et al. 1996).
Durante el tercer estro, cuando por ultrasonografía se estableció la presencia de un folículo preovulatorio, a las hembras se les extirpó el ovario que contenía dicho folículo, mediante abordaje lateral derecho. Se practicó la técnica descrita por Peiró et al. (2009).
Para el procedimiento quirúrgico las vacas fueron sedadas con xilacina aplicada por vía intramuscular (IM) a una posología de 0,1 a 0,2 mg/kg y se les aplicó anestesia paravertebral con lidocaína al 2%.
Una vez obtenidos los ovarios, se realizó un corte diametral sobre el estigma del folículo preovulatorio para seccionarlo en 2 partes simétricas.
Una de las partes del folículo preo-vulatorio se conservó en solución acuosa de formalina al 10% de pH 7,4 hasta su procesamiento en el Laboratorio de histotécnica de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad del Tolima.
En el sitio de máximo diámetro folicular se realizaron 5 cortes consecutivos de 5 μm de grosor, los cuales se tiñeron con hematoxilina y eosina (H&E). Se siguieron las técnicas estandarizadas en el laboratorio. La morfometría folicular se determinó en el laboratorio de reproducción animal de la FMVZ de la Universidad del Tolima con microscopio de luz a 40x y la ayuda del software Image Pro Plus; la escala del software fue calibrada (figura 1).
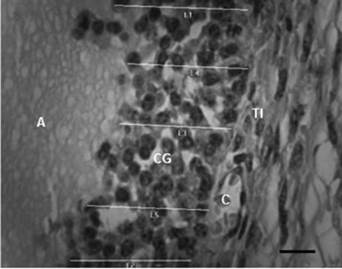
Fuente: elaboración propia.
FIGURA 1 Medidas de la capa de cg en folículo ovárico de hembra bovina de la raza Brahman. Barra de escala = 10 pm. Antro (A), células de la granulosa (CG), células de la teca interna (TI), capilar (C)
Para cada corte se estableció como radio del antro folicular el valor del diámetro, obtenido mediante la última ecografía previa a la extirpación del ovario, dividido en 2 (r = D / 2).
Para deducir el volumen ocupado por las CG y el de la TI, se calculó el radio total del folículo. Para esto, se adicionó al radio del antro folicular el promedio del grosor de las capas de CG y de la TI.
Una vez obtenido el radio total, asumiendo una forma perfectamente esférica de los folículos, se utilizó la fórmula de la esfera para el cálculo del volumen del antro: v = 4/3 • Π • r3 (Rodrigues et al. 2018).
El volumen ocupado por las capas de CG y de la TI del folículo se calculó por diferencias entre el volumen del folículo completo, el volumen ocupado por el antro folicular y el volumen ocupado por el antro folicular más la capa de CG.
El análisis estadístico de los resultados se realizó por análisis exploratorio de datos. Se aplicó la prueba de T de Student y se realizaron análisis de correlación de Pearson, mediante el software Info Stat, versión 2008 (Di Rienzo et al. 2008).
RESULTADOS
Parámetros del periodo interestros
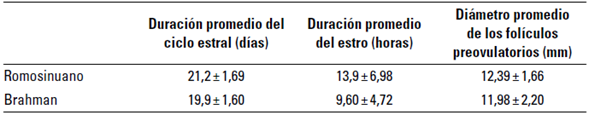
Una vez realizado el seguimiento ecográfico a las hembras de las 2 razas estudiadas, se determinaron los valores promedio del periodo interestro, duración del estro, tiempo de ovulación y tamaño del folículo preovulatorio. Estos valores se presentan en la tabla 1.
TABLA 1 Duración promedio del ciclo estral y estro, diámetro medio de vacas Romosinuano y Brahman bajo condiciones de bosque seco tropical (bs-T)
Fuente: elaboración propia.
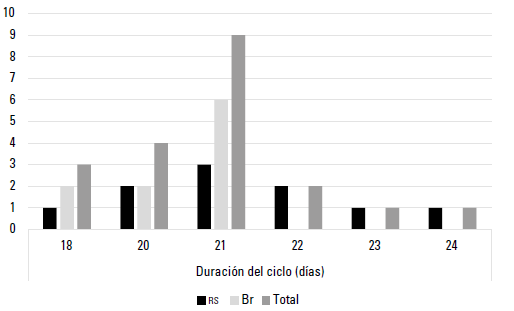
En la duración de ciclo estral no hubo diferencias entre razas (p > 0,05). Para la raza RS se observaron valores promedios de 21,2 ± 1,69 días, con variaciones entre 18 y 24 días y para Br 19,9 ± 1,60 días con variaciones entre 17 y 21 días (figura 2).
Fuente: elaboración propia.
FIGURA 2 Distribución promedio de la duración del ciclo estral en las razas RS y Br
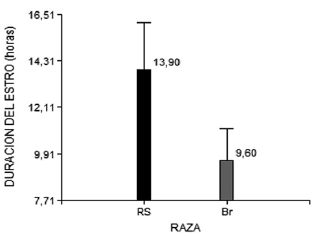
La duración del estro de 13,9 ± 6,98 y 9,60 ± 4,72 h para las razas RS y Br, respectivamente, fue igual en promedio (p > 0,05). En la raza RS se presentaron variaciones entre 4 y 24 h, mientras que en la raza Br se observaron estros de entre 4 h y 19 h (figura 3).
Fuente: elaboración propia.
FIGURA 3 Distribución promedio de la duración del estro en las razas RS y Br
Se observó, además, tendencia a una mayor aparición de estros durante las primeras horas de la mañana en la raza RS, 46,66% vs. 20% en la raza Br. No obstante, al totalizar los estros observados a primera hora en la mañana, es decir, con inicio en horas de la madrugada, y relacionarlos con los observados con inicio en horas de la noche para ambas razas, se encontró en ambos casos 7/15 celos; es decir, 46,66% de los estros iniciaron en horas de la noche. No se observaron estros anovulatorios en ninguna de las 2 razas estudiadas, ni ovulaciones sin evidencia de estro.
El tiempo transcurrido desde observación de los primeros signos del estro hasta la ovulación, fue de 21,20 ± 5,07 h para los animales de la raza Br, con variaciones desde 17 hasta 31 h y de 24,40 ± 6,43 h para los de la raza RS con un valor mínimo de 16 h y uno máximo de 35. No se observó diferencia estadísticamente significativa entre las 2 razas (p > 0,05).
La concentración media de progesterona sérica en este día para las hembras de los 2 grupos raciales fue de 0,51 ± 0,30 ng/ml para las de la raza RS y de 0,61 ± 0,27 para las de la raza Br. No se detectaron diferencias significativas entre las 2 razas (p > 0,05).
Medidas de los componentes foliculares
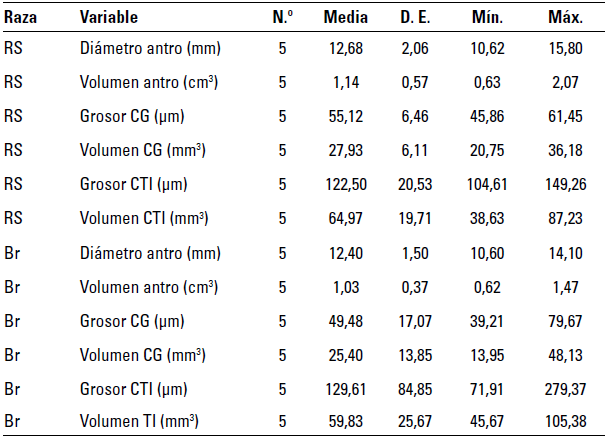
Los resultados de los promedios de las medidas de longitud de los componentes foliculares para las 2 razas en estudio se presentan en la tabla 2.
TABLA 2 Distribución de promedios de las medidas foliculares de longitud y volumen en folículos preovulatorios de las razas RS y Br
Fuente: elaboración propia.
En ella se puede apreciar que el diámetro de los folículos preovulatorios de las hembras de la raza RS presentó un promedio de 12,68 ± 2,06 mm en el momento previo a la ovulación, aunque se observaron variaciones en los datos individuales de las hembras entre 10,62 y 15,8 mm.
En las hembras de la raza Br el diámetro promedio de los folículos preovulatorios, establecido por ultrasonografía, presentó variaciones en los datos individuales entre 10,6 mm y 14,1mm, con promedio de 12,40 ± 1,50 mm. En las hembras estudiadas no hubo diferencias significativas en el diámetro de los folículos preovulatorios entre las razas RS y Br (p > 0,05). Se encontró que en los animales estudiados no se observaron diferencias significativas (p > 0,05) en lo relacionado con el volumen folicular, el cual fue de 1,14 ± 0,57 cm3 y de 1,03 ± 0,37 cm3 para las razas RS y Br, respectivamente.
El grosor de la capa de CG para los animales de la raza RS registró un promedio de 55,12 ± 6,46 μm, este parámetro mostró variaciones individuales en los animales estudiados entre 45,86 y 61,45 μm mientras que para la raza Br el promedio fue de 49,48 ± 17,07 μm, con valores mínimos y máximos de 39,21 y 79,67 μm, respectivamente, lo que establece que no hubo diferencias estadísticamente significativas entre las 2 razas (p > 0,05).
Al calcular el volumen ocupado por la capa de CG (tabla 2), parámetro tridimensional que asocia el grosor de la capa con el tamaño del folículo preovulatorio, no se encontraron diferencias estadísticamente significativas entre las 2 razas (p > 0,05), con valores promedio de 25,40 ± 13,85 mm3 para la raza Br y 27,93 ± 6,11 mm3 para la raza RS.
En cuanto a lo relacionado con la capa de células de la TI, el grosor promedio de esta para la raza RS fue de 122,50 ± 20,53 104,61 - 149,26 μm, entretanto para la raza Br fue de 129,61 ± 84,85 con variaciones desde 71,91 hasta 297,37 μm. Estadísticamente no se hallaron diferencias significativas entre las 2 razas (p > 0,05).
Al calcular el volumen ocupado por la capa de células de la TI (tabla 2), parámetro tridimensional que asocia el grosor de la capa con el tamaño del folículo preovulatorio, no se encontraron diferencias estadísticamente significativas entre las 2 razas (p > 0,05), con valores promedio de 64,97 ± 19,71 mm3 para la raza RS y 59,83 ± 25,67 mm3 para la raza Br.
Desarrollo del cuerpo lúteo y concentración sérica de P4 durante el ciclo estral
La observación diaria de los ovarios permitió identificar claramente el CL por ultrasonografía en promedio a los 5,0 ± 1,93 días posovulación, con variaciones entre 2,0 y 8,0 días. En las hembras de la raza RS se observó en promedio a los 5,38 ± 1,77 días, mientras que en las de la raza Br, a los 4,63 ± 2,13 (tabla 3).
TABLA 3 Distribución de promedios de desarrollo del cuerpo lúteo y concentración sérica de P4 durante el ciclo estral de las razas RS y Br

Día 0 = estro.
Fuente: elaboración propia.
El momento de máximo desarrollo estructural de los cuerpos lúteos para la totalidad de las hembras estudiadas, en promedio, ocurrió el día 12,6 ± 4,05 (día 0 = estro), con un diámetro promedio de 22,92 ± 3,60 mm. Para las de la raza RS el diámetro máximo se observó en promedio el día 14,88 ± 3,4, con 23,06 ± 3,9 mm, mientras que para las de la raza Br fue el día 10,00 ± 3,16 con un diámetro promedio de 22,75 ± 3,16 mm. No se encontraron diferencias significativas para el diámetro máximo del CL (p > 0,05) entre las razas estudiadas, mientras que sí se observaron diferencias para el día en que se presentó esta condición, entre las razas RS y Br (p > 0,05).
La concentración media de progesterona para el día en que se registró el máximo diámetro del CL fue de 5,37 ± 1,38 ng/ml para la raza RS y 5,74 ± 0,89 ng/ml para la raza Br. No se presentaron diferencias significativas entre razas (p > 0,05), con media general para los animales experimentales de 5,54 ± 1,15 ng/ml (tabla 3).
Al analizar el grado de relación entre el tamaño del CL y la concentración media de P en este día, se observó en los animales estudiados una débil correlación inversa entre el diámetro del CL y la concentración media de P4 en el suero sanguíneo (coeficiente de correlación de Spearman:-0,23, p > 0,05).
DISCUSIÓN
Duración del ciclo estral
La duración del ciclo estral en las hembras de la raza Br en este trabajo osciló entre 18 y 21 días, con promedio de 19,9 ± 1,6 días. Este parámetro se encuentra dentro de los rangos reportados como normales para la especie bovina en otras latitudes y concuerda con lo reportado por Enjoy et al. (2012) para hembras Br en Venezuela con promedio de 21,3 ± 0,7 días y variaciones entre 19-24 días.
Los valores obtenidos para la duración del ciclo estral en las hembras de la raza RS oscilaron entre 18 y 24 con promedio 21,2 ± 1,69 días, lo que coincide con lo revisado por Forde et al. (2011). Estos se distribuyeron de tal manera que el 10% de los ciclos estrales fueron de 18 días, el 20% de 20 días, el 30% de 21, el 20% de 22 días, el 10% de 23 días y el 10% de 24 días. El 80% de los ciclos observados fueron de 2 ondas de crecimiento folicular y el 20% de 3.
Duración del estro
Hubo una alta proporción de aparición de estros durante las primeras horas de la mañana (46,66%) en las 2 razas objeto de estudio, lo que coincide con lo reportado por Borges et al. (2004), quienes encontraron que la mayoría de los estros fueron observados en horas de la mañana (57,3%) en hembras de las razas Gir y Nelore Esta situación facilita la tarea de detección del estro y ayuda establecer tiempos para los procedimientos en los programas de inseminación artificial a celo detectado, lo que redunda en buenos indicadores de eficiencia reproductiva de las hembras en programas de reproducción asistida. Los hallazgos podrían deberse al estrés por calor (Góngora y Hernández, 2010), pues este altera el desarrollo y la dominancia folicular durante los primeros 8 días del ciclo estral. Si este efecto se mantiene en forma crónica, la actividad de la aromatasa y las concentraciones de estradiol (E2) en el líquido folicular disminuyen (Badinga et al. 1985).
Se encontró que la duración del estro en animales de razas Bos indicus fue de unas 10 h, más breve que en Bos taurus (Galina y Arthur, 1990). Así mismo, se reportó que hubo amplias variaciones en la duración, desde 1,3 hasta 20 h y en la mayoría de las vacas (53,8%) el estro empezó durante la noche (entre las 18:00 y las 06:00 h) y en el 34,6% de ellas el estro inició y finalizó durante la noche (Pinheiro et al. 1998).
Intervalo estro-ovulación
En los animales de las 2 razas objeto de estudio no se presentaron estros anovu-latorios a diferencia de lo observado en otras latitudes, pues se ha reportado que entre 3,0 y 8,4% de los animales de la raza Br presentan estros anovulatorios, al igual que un porcentaje entre 22 y 31% de ovulaciones sin evidencia de estro (Plasse et al. 1968). Se puede deducir que no hubo presentación de estros anovulatorios, pues en las hembras en las que se observó estro evidenciaron con el examen ultrasonográfico un folículo de tamaño preovulatorio, el cual desapareció con la consecuente aparición de un cuerpo lúteo en el lugar de aparente ocupación del mencionado folículo, CL que se hizo evidente en la ecografía 5 días más tarde. Es factible proponer que la escogencia de los animales utilizados en el presente estudio, además de su reconocida fertilidad, estuvieron bajo condiciones ambientales y de alimentación adecuadas y, por ello, se facilitó la expresión de los parámetros fisiológicos estimados.
El tiempo transcurrido desde la observación de los primeros signos de quietud al intento de monta por parte del macho hasta la ovulación fue de 21,20 ± 5,07 h para los animales de la raza Br, y de 24,40 ± 6,43 h para los de la raza RS, datos que no coinciden con lo reportado por Walker et al. (1996) en hembras Holstein (27,6 ± 5,4 h) y por Pinheiro et al. (1998) en hembras Nelore (26,6 ± 0,44 h), quienes además no encontraron diferencias en el tiempo transcurrido desde la detección del estro hasta la ovulación entre estros espontáneos o inducidos, pero sí coinciden con lo reportado en hembras Gir (23,9 ± 5,1 h) y Nelore (21,5 ± 6,2 h) (Borges et al. 2004).
La caracterización de este intervalo es esencial para la estimación del mejor tiempo para la inseminación en el ganado bovino (Pinheiro et al. 1998). En novillas Br, algunos reportes más recientes muestran intervalos un poco más largos, alrededor de 25-29 h (Cavalieri et al. 1997; Pinheiro et al. 1998), discrepancia que se ha sido atribuida al grado de precisión de los métodos utilizados para la detección del inicio y finalización del estro, como en el caso de la uso de dispositivos electrónicos instalados en los animales. La precisión en la detección puede también verse afectada por factores como el estrés crónico (Walker et al. 2008).
Si bien la evaluación morfológica de los folículos es indudablemente popular, su precisión como predictor de la calidad del oocito es todavía incierta (Perry et al. 2005), el tamaño de los folículos antra-les podría ser un valioso marcador para los procesos de fertilización in vitro, en el presente trabajo durante la evaluación ultrasonográfica no se encontró diferencia en el diámetro del folículo preovulatorio entre las razas RS y Br, 12,68 ± 1,85 vs. 12,40 ± 1,35 mm, respectivamente (> 0,05). Esto podría explicarse por el hecho de tratarse de folículos de más de 10 mm de diámetro, puesto que se encontró que el 80% de los animales con folículos de más de 8,5 mm de diámetro y el 90% de los de más de 10 mm ovulan en respuesta a una dosis de 25 mg de la LH (Gimenes et al. 2008).
Los valores encontrados para la raza Br difieren de los hallados por otros autores en latitudes similares. Enjoy et al.(2012), por ejemplo, encontraron en vacas Br que el diámetro de los folículos preovulatorios fue de 12,7 ± 0,4 mm, parámetro que estadísticamente no dista de lo encontrado en el presente trabajo y difiere de lo reportado para ganado Nelore (11 mm), otra de las razas Bos indicus de importancia en el trópico cálido (Figueiredo et al. 1997). No obstante estas diferencias, se cree que el tamaño del folículo preovulatorio aparentemente no tiene efecto sobre la fertilidad cuando la ovulación ocurre espontáneamente, ya que este ovula en un amplio rango de tamaños cuando está fisiológicamente maduro (Perry et al. 2005), lo que mostraría que el diámetro de los folículos no necesariamente indica si el folículo está saludable y en crecimiento o si está sufriendo atresia (Han et al. 2006).
Desarrollo de componentes foliculares
La tasa de crecimiento de los folículos ováricos es relativamente grande y el aumento en el tamaño de estos es el producto de la proliferación de las células que lo componen y, principalmente, del aumento en el contenido de líquido folicular. Se ha observado que la tasa de crecimiento folicular varía desde 0,78 ± 0,1 mm/d (Enjoy et al. 2012), lo que obliga a pensar que la expresión de la respuesta esteroidogénica tanto del folículo ovárico, como del CL estarían dependiendo del desarrollo de los mismos y de su tasa metabólica.
El incremento en el número de CG del folículo preovulatorio probablemente podría resultar en un mayor número de células luteales grandes, con un subsecuente incremento en el tamaño del CL e incremento en la secreción de P4 (Sartori et al. 2002). En el presente trabajo se pudo encontrar que no se registraron diferencias estadísticamente significativas en el grosor de la capa de CG entre los folículos preovulatorios de los animales de la raza RS (55,12 ± 15,51 μm) y los de la raza Br (49,48 ± 17,07 [m), ni en el volumen ocupado por estas (27,93 ± 6,11 vs. 25,40 ± 13,85 mm3, respectivamente), lo que haría suponer que el subsecuente CL debería haber alcanzado un volumen similar si se hubieran permitido las ovulaciones; sin embargo, la información previa obtenida de los folículos y CL de los ciclos anteriores permite suponer que no se hubieran presentado diferencias significativas en el tamaño. Aunque se ha observado que la inducción de la ovulación en novillas durante su primera onda de crecimiento folicular se refleja en la formación de cuerpos lúteos de menor tamaño y con menor capacidad para sintetizar esteroides comparados con aquellos cuerpos lúteos provenientes de una ovulación espontánea, al parecer por una menor expresión de receptores para LH (Gregson et al. 2016), la diferencia en tamaño podría ser compensada por la tasa de síntesis de esteroides, estimulada probablemente por la acción de otras moléculas no analizadas en el presente estudio, pues se ha reportado, por ejemplo, que la LH junto con el factor de crecimiento similar a la insulina tipo I (IGF-I) estimulan la esteroidogénesis en las células de la TI de los bovinos (Abreu et al. 2018).
CONCLUSIONES
Con base en los resultados obtenidos en el presente estudio, se podría establecer que no existen diferencias en los eventos fisiológicos estudiados entre las razas RS y Br; es importante seguir indagando en detalle en los procesos fisiológicos no solo durante el interestro, sino también durante otras etapas de la vida productiva de los animales, como durante el posparto y la gestación temprana, sin olvidar que las características reproductivas son de carácter multifactorial y que para efectos del presente trabajo solo se han abordado algunos elementos relacionados con la fisiología ovárica como aporte al conocimiento de la especie bovina que, sin duda, pueden servir de base para futuros análisis en los bovinos residentes en el trópico.

