










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiomédica
Print version ISSN 0120-4157On-line version ISSN 2590-7379
Biomédica vol.28 no.4 Bogotá Oct./Dec. 2008
1 Laboratorio de Biología Molecular y Patogénesis, Departamento de Ciencias Fisiológicas, Escuela de Ciencias Básicas, Facultad de Salud, Universidad del Valle, Cali, Colombia
2 Facultad de Medicina, Universidad Santiago de Cali, Cali, Colombia
3 Sección de Farmacología, Departamento de Ciencias Fisiológicas, Facultad de Salud, Universidad del Valle, Cali, Colombia
4 Facultad de Medicina, Universidad del Sinú, Montería, Córdoba, Colombia
Recibido: 14/12/07; aceptado:13/08/08
Introducción. Trabajos previos han aportado evidencias de que en la paraparesia espástica tropical/mielopatía asociada con el virus linfotrópico humano tipo I, existe un componente autoinmune asociado a su patogénesis.
Objetivo. Evaluar el estado autoinmune y la existencia de mimetismo molecular en pacientes con paraparesia espástica tropical del pacífico colombiano.
Materiales y métodos. A partir de muestras de plasma de 37 pacientes con paraparesia espástica tropical/mielopatía asociada al HTLV-I, 10 con leucemia de células T del adulto, 22 individuos portadores asintomáticos y 20 seronegativos para el HTLV-I, se determinaron niveles plasmáticos de anticuerpos antinucleares y anticardiolipina-2 y de interferón-g e interleucina-4. Se evaluó, por Western blot, la reactividad cruzada de plasmas contra proteínas obtenidas de varias fuentes celulares normales del sistema nervioso. Además, se estudió la reactividad cruzada de plasmas de seropositivos y del anticuerpo monoclonal LT4 anti-taxp40 en secciones de médula espinal de ratas Wistar no infectadas.
Resultados. El 70,2% y el 83,8% de los pacientes con paraparesia espástica tropical fueron reactivos para anticuerpos ANA y ACL-2, respectivamente, en contraste con los de leucemia de células T del adulto y los seropositivos asintomáticos (P<0,001). Además, el 70,3% y el 43,2% de los pacientes con paraparesia espástica tropical tuvieron niveles detectables de IFN-g e IL-4, respectivamente. El anticuerpo LT4 anti tax-p40 y los plasmas de paraparesia espástica tropical/mielopatía asociada al HTLV-I mostraron una reacción cruzada con una proteína de PMr 33-35 kDa, obtenida del núcleo de neuronas de la médula espinal de ratas Wistar no infectadas.
Conclusión. Se obtuvieron evidencias que apoyan la existencia de un síndrome autoinmune mediado por mimetismo molecular como parte de la etiopatogénesis de la degeneración axonal observada en la paraparesia espástica tropical en pacientes colombianos de la costa pacífica.
Palabras clave: paraparesia espástica, virus linfotrópico de células T humanas tipo 1, virus 1 T-linfotrópico de los primates, médula espinal, autoanticuerpos, autoinmunidad, imitación molecular.
Introduction. Previous reports have given evidence that in tropical spastic paraparesis (TSP)/ human T-lymphotrophic virus (HTLV-I)-associated myelopathy (HAM), an autoimmune process occurs as part of its pathogenesis.
Objective. The roles of autoimmunity and the molecular mimicry was evaluated in TSP/HAM patients.
Materials and methods. Plasma samples were characterized from patients in the Pacific coastal region of Colombia. Thirty-seven were identified as TSP/HAM, 10 were diagnosed with adult T-cell leukemia virus, 22 were asymptomatic carriers but seropositive for HTLV-I and 20 were seronegative and served as negative controls. Plasmatic levels of the following were determined: antinuclear antibody (ANA) levels, anticardiolipine-2 (ACL_2), interferon- (IFN-g) and interleukin-4 (IL-4). Using Western blot, the crossreactivity of the seropositive and seronegative samples was evaluated against proteins extracted from several central nervous system components of non infected Wistar rats. The HTLV-I seropositive plasmas were crossreacted with a monoclonal tax (LT4 anti-taxp40) from spinal cord neurons of non infected Wistar rats.
Results. Of the TSP/HAM patients, 70.2% were reactive against ANA and 83.8% against ACL-2, in contrast with those ATL and asymptomatic seropositives subjects that were not reactive (P<0.001). Moreover, 70.3% had detectable levels of IFN and 43.2% had detectable IL-4. LT4 anti-taxp40 and plasma of TSP/HAM exhibited cross reactivity with a MW 33-35 kDa protein from the rat spinal cord nuclei.
Conclusion. Support was provided for the existence of an autoimmune syndrome mediated by molecular mimicry; the syndrome was responsible for some of the axonal degeneration observed in TSP/HAM patients.
Key words: paraparesis, spastic; human T-lymphotropic virus 1, primateT-lymphotropic virus 1, spinal cord, autoantibodies, autoimmunity, molecular mimicry.
El virus linfotrópico humano tipo I (HTLV-I) está asociado con un amplio espectro de manifestaciones clínicas entre las que se incluyen la leucemia de células T del adulto y la paraparesia espástica tropical/mielopatía asociada al HTLV-I. Sin embargo, en la mayoría de los casos, los individuos infectados permanecen asintomáticos. A luz de varios resultados que muestran una significativa respuesta inmune en estos pacientes, se ha sugerido la participación del sistema inmunológico en la patogénesis de la paraparesia espástica tropical (1).
La paraparesia espástica tropical es una polirradiculoneuropatía crónica inflamatoria que cursa con dificultad para caminar, la cual es lentamente progresiva. Se observan ligeros cambios sensitivos, debilidad motora y espasticidad en los miembros inferiores, aumento de la frecuencia, urgencia o retención urinarias, estreñimiento e impotencia o frigidez sexual, además de dolor lumbar (2-4). Esta entidad clínica se presenta principalmente en personas mayores de 30 años y predomina en el sexo femenino (5).
Varios estudios efectuados en Colombia han demostrado que la paraparesia espástica tropical es un problema de salud pública importante en la región costera del Pacífico colombiano (6,7). Así, en Tumaco, un municipio del suroccidente del Pacífico, la tasa de casos de la enfermedad por año se ha calculado en aproximadamente 2,8 x 10-3, hecho que contrasta con su baja presencia en otras zonas que son endémicas para la infección por el HTLV-I (8).
A pesar de que epidemiológicamente las dos principales enfermedades asociadas con la infección por el HTLV-I son la leucemia de células T del adulto y la paraparesia espástica tropical, resultados previos también muestran la infección por el HTLV-I como factor importante en ciertas enfermedades de origen autoinmune, como el síndrome de Sjögren (9,10) y el lupus eritematoso sistémico (11), entre otras. Sin embargo, la mayoría de los individuos seropositivos para el virus permanecen asintomáticos y el riesgo a desarrollar enfermedades es muy bajo (12-14).
Con base en lo anterior, se ha abierto un nuevo camino hacia la elucidación de la etiología de la enfermedad; se ha postulado la hipótesis de que la paraparesia espástica tropical o mielopatía asociada al HTLV-I resulta, no de la acción directa del virus sobre las células afectadas, sino como consecuencia de un proceso autoinmune mediado por el mimetismo molecular existente entre anticuerpos específicos contra las proteínas del HTLV-I y componentes celulares del sistema nervioso central. Este proceso, que no es más que la reacción cruzada ocurrida entre los anticuerpos generados como parte de la respuesta inmune a un agente infeccioso y autoantígenos, ha sido confirmado en el complejo sida/demencia (15) y se ha sugerido recientemente para otras enfermedades degenerativas del sistema nervioso central como la esclerosis múltiple (16) y la misma paraparesia espástica tropical (17,18).
Con el objetivo de aportar evidencias que sustenten la hipótesis de la autoinmunidad y de un mimetismo molecular como parte de la etiopatogénesis de la paraparesia espástica tropical en la costa pacífica colombiana, se evaluaron los niveles de anticuerpos antinucleares (ANA) y anticardiolipina 2 (ACL-2), además de los de interleucina-4 (IL-4) y de interferón-g (IFN-g) en plasmas de pacientes con paraparesia espástica tropical y de portadores asintomáticos de la región del suroccidente colombiano. Por otra lado, se analizó la existencia de reacción inmunológica cruzada hacia componentes celulares del sistema nervioso central, y se probó la reactividad del anticuerpo monoclonal LT4 anti-taxp40 y de plasmas de pacientes con paraparesia espástica tropical, leucemia de células T del adulto, de individuos portadores asintomáticos y seronegativos para el HTLV-I contra blancos celulares normales del sistema nervioso central.
Materiales y métodos
Sujetos y muestras
Se incluyeron 37 pacientes con paraparesia espástica tropical, 22 individuos seropositivos asintomáticos, 20 individuos controles seronegativos para HTLV-I de la costa pacífica colombiana y 10 pacientes con leucemia de células T del adulto de diferentes regiones de Colombia. Los criterios para hacer el diagnóstico de paraparesia espástica tropical/mielopatía asociada al HTLV-I fueron aquellos descritos por Osame et al. (1987) (3). Los individuos con leucemia de células T del adulto fueron referenciados de la historia clínica de remisión como caso de leucemia con diagnóstico serológico positivo para anticuerpos HTLV-I. Los pacientes seropositivos asintomáticos se obtuvieron por el criterio de exclusión con base en lo propuesto por Osame et al. en 1987 (3); finalmente, los controles negativos se incluyeron por su ausencia de anticuerpos contra HTLV-I. A todos los individuos incluidos en este estudio se les informó los objetivos del mismo y aceptaron donar muestras de sangre total que fue obtenida por venopunción.
De todos los voluntarios, se obtuvieron 20 ml de sangre que fue colocada en medio anticoagulante y almacenada apropiadamente hasta su procesamiento. Cada una de las muestras de sangre se sometieron a un gradiente de Ficoll-Hipaque, recolectándose las células linfocitarias de sangre periférica; éstas fueron criopreservadas en medio conservante RPMI/DMSO (50% DMSO en medio RPMI 1640) a -156 °C; de otra parte, el plasma de cada muestra se almacenó en alícuotas a -20 °C para su posterior utilización.
Todas las muestras de plasma se examinaron para determinar la presencia de anticuerpos contra el HTLV-I mediante la aplicación de una prueba de micro-Elisa utilizando el estuche ELISA Murex HTLV I-II (Murex Biotech Limited, Dartford. UK). Las pruebas se hicieron siguiendo las recomendaciones del fabricante. Las positivas para HTLV-I se reconfirmaron mediante Western blot utilizando el estuche comercial Immunblot HTLV blot 2.4 (Genelabs® Diagnostics Pte Ltd., Singapore Science Park, Singapore).
Todos los plasmas de individuos participantes en el estudio se examinaron para la presencia de anticuerpos contra los virus de inmunodeficiencia humana 1 y 2, utilizando el estuche comercial micro-ELISA IgG-anti-HIV-1/2 (Enzygnost®, Behring Dade, Germany), y se reconfirmaron mediante Western blot.
Determinación de las diferentes clases de inmunoglobulinas
Los niveles de las inmunoglobulinas plasmáticas IgG, IgM e IgA1 reactivas para antígenos del HTLV-I, se midieron mediante ELISA. Para ello, se usó el estuche comercial HTLV-I 2.0 EIA (ABBOTT, Abbott Park, IL, USA). La determinación de anticuerpos anti-HTLV-I de las clases IgG, IgM e IgA, se efectuó utilizando los correspondientes conjugados obtenidos de Sigma Life Science (St. Louis, MO). Se realizaron pruebas de calibración empleando sueros de referencia. Las muestras de plasma se diluyeron a concentraciones apropiadas en 2% de albúmina sérica bovina fracción V en PBS 1X pH 7,4. La incubación de cada una de las muestras se efectuó en diluciones de 1:40 a 1:200 a 42 °C durante una hora. Los pasos posteriores se realizaron siguiendo las recomendaciones del fabricante. La densidad óptica de cada muestra se midió con ayuda de un equipo Quantum (Abbott, Abbott Park, IL, USA). La detección de anticuerpos para cada isotipo se registró en valores de absorbancia a 492 nm. Los valores de punto de corte se calcularon con base en lo recomendado por el fabricante.
Células y tejidos
En este estudio se incluyeron las siguientes fuentes celulares que hacen parte de la colección de células y tejidos de la Fundación in vitro de la Facultad de Salud, Universidad del Valle: cultivos primarios de cerebro humano que fueron mantenidos en DMEM con suplemento al 5% de suero fetal bovino y estimulado con factor de crecimiento neuronal. Los fibroblastos de pulmón humano, fibroblastos de piel humana, riñón embrionario humano, células pluripotenciales de cordón umbilical, además de las líneas celulares Hela y 3T3 se replicaron y mantuvieron en crecimiento por tiempos variables en MEM adicionado con suero fetal bovino al 5% y mezcla de antibióticos; para los cultivos primarios se incluyó factor de crecimiento de fibroblastos.
Por otra parte, se emplearon preparaciones de proteínas obtenidas de biopsias post mórtem de médula espinal de rata, cerebro fetal humano e hígado humano. Todas las muestras de tejido post mórtem humano se obtuvieron cumpliendo con los procedimientos legales requeridos para la obtención de este tipo de tejido (Resolución 8430 del Ministerio de la Protección Social de Colombia, 4 de octubre de 1993). Éstas se conservaron en medios apropiados y se criopreservaron a -156 °C hasta su utilización.
Anticuerpos monoclonales y policlonales
Se utilizó el anticuerpo monoclonal LT4 (anti-tax-p40) (18-20). Se emplearon los sueros policlonales para la proteína acídica fibrilar el IgG-anti-GFAP (N-18) (Santa Cruz Biotechnology. Santa Cruz, CA) y para neurofilamentos, el NCFL-NF68 (Novocastra, UK).
Inmunoblots
Aproximadamente, 107 células/ml de cada uno de los cultivos primarios y líneas celulares cultivadas incluidas en este estudio, se precipitaron por centrifugación; luego se resuspendieron y se incubaron durante 30 minutos a 0 °C con solución tampón preenfriada de lisis RIPA (NaCl 150 mM, NP-40 1%, DOC 0,5%, SDS 10%, Tris 50 mM pH 8,0). Se colocaron en nitrógeno líquido 2 g de médula espinal humana y de rata y corteza cerebral que se maceraron en solución tampón de lisis RIPA; posteriormente, los lisados se clarearon mediante centrifugación a baja velocidad (2.000g); el sobrenadante se centrifugó a 12.000g y el precipitado se resuspendió en PBS.
La concentración de proteínas totales de cada uno de los lisados se determinó usando el estuche Protein Assay (Bio-Rad, Richmond, CA) de acuerdo con las recomendaciones del fabricante. Concentraciones de proteínas totales de 100 a 300 µg/ml de los lisados obtenidos, se sometieron a electroforesis en geles de poliacrilamida al 12% con dodecil sulfato de sodio (SDS) en condiciones desnaturalizantes (SDS-PAGE). Las proteínas separadas electroforéticamente, fueron transferidas en forma pasiva a membranas de nitrocelulosa, de acuerdo con lo descrito por Burnette et al. (21) y Barin et al. (22). Después de la transferencia, se bloquearon con solución de 10% de albúmina bovina fracción V en solución salina concentrada de fosfatos pH 7,4 (PBS, pH 7,4) durante 1 hora a 37 °C con agitación continua. Posteriormente, se incubaron en una dilución 1:100 de suero humano normal (Sigma, Chemicals, St. Louis, USA) durante 16 horas a 4 °C y se almacenaron -20 °C hasta su utilización.
Las membranas de nitrocelulosa con las proteínas transferidas, se ensayaron en dilución 1:2.000 de los correspondientes plasmas. El LT4 anti-taxp40 se ensayó en diluciones de 1:5.000 y 1:10.000. Las preparaciones se incubaron en 1% de albúmina bovina fracción V, PBS, pH 7,4 y 0,2% de Tween 20 por 16 horas a 4 °C. Los anticuerpos ligados se detectaron mediante incubaciones secuenciales con anti-IgG-humana o de ratón, según el caso, marcada con biotina/avidina diluida 1:1.000 (Amersham, UK) y reveladas por incubación con 0,05% de diaminobenzidina y peroxido de hidrógeno (Sigma, St. Louis, MO, USA) en PBS, de acuerdo con lo descrito por Barin et al. (21). Entre cada incubación se hicieron tres lavados de 15 minutos cada uno en PBS, pH 7,4, y 0,2% de Tween 20, excepto en el último lavado, antes del revelado, en que se lavó sólo con PBS, pH 7,4.
La determinación del peso molecular relativo aproximado de las diferentes proteínas (PMr) se efectuó calculando la movilidad electroforética relativa (Mr) de proteínas de un patrón comercial de proteínas de amplio rango (6,5 a 200 kDa) (Bio-Rad, USA) en un gel de poliacrilamida-SDS del 10%. Con base en esta medición, se construyó una gráfica del logaritmo del peso molecular de las proteínas del patrón comercial versus su movilidad relativa. Sobre esta gráfica se extrapoló el valor de Mr de las proteínas obtenidas de las diferentes fuentes celulares y de tejidos sobre la ordenada logarítmica de peso molecular; de esta forma se les calculó su peso molecular relativo (PMr).
Marcadores de autoinmunidad
La presencia de anticuerpos antinucleares (ANA) y anticardiolipina 2 (ACL-2), se determinó mediante ELISA (Sigma Diagnostic, St. Louis, USA) siguiendo el procedimiento recomendado por el fabricante. Los niveles de IL-4 e IFN-g se determinaron mediante ELISA de doble captura (Bioscience Cytoscreen SA, USA), también siguiendo los protocolos recomendados por el fabricante; para el ensayo correspondiente se emplearon 100 µl de plasma. De la utilización de estos estuches se obtuvo una determinación semicuantitativa de las concentraciones de citocinas plasmáticas en pg/ml. Para IL-4, el rango de detección del estuche varió de 500 a 0,8 pg/ml, mientras que para IFN-g varió de 200 a 0,3 pg/ml. En estos rangos se observó un comportamiento linear de la recta construida con base en el patrón incluido en cada uno de los estuches.
Inmunohistoquímica
Las preparaciones histológicas de biopsias de médula espinal de ratas Wistar no infectadas, se incubaron con el anticuerpo monoclonal LT4 y plasmas de individuos HTLV-I positivos así como de un conjunto de plasmas seronegativos controles, utilizando el protocolo de Hsu et al. (1981) (23). La actividad de peroxidasa endógena de las fuentes biológicas se bloqueó mediante incubación de los especímenes en solución de peróxido de hidrógeno al 3% durante 35 minutos a temperatura ambiente; posteriormente, se realizaron dos lavados de cinco minutos en solución 0,01 de PBS, pH 7,5. El rescate del antígeno se realizó de acuerdo con Shi et al. (1995) (24). Las preparaciones se incubaron toda la noche en diluciones 1:500 del correspondiente anticuerpo monoclonal; para los plasmas se emplearon diluciones 1:200. Después de la incubación, las placas se lavaron como se describió previamente y se incubaron con el anticuerpo conjugado biotinilado IgG de chivo anti-ratón o anti-humano, según el caso, diluido 1:200 en PBS, pH 7,5, durante 30 minutos a temperatura ambiente.
Después de este procedimiento, se lavaron tres veces durante cinco minutos en PBS, pH 7,5. Posteriormente, se incubaron con el ABC Elite PK4000 (Vector Inc.) durante 30 minutos. Después de este periodo de incubación, se lavaron de nuevo en PBS y se revelaron con diaminobencidina (Sigma, MO) previamente disuelta en solución Tris-HCl pH 7,6 y 0,003% de peróxido de hidrógeno. Las preparaciones histológicas fueron teñidas con hematoxilina de Gills y se montaron de manera permanente en permount. Como control negativo de los experimentos se incluyó plasma de sujetos seronegativos para el HTLV-I y VIH1/2.
Cálculos estadísticos
El tamaño mínimo de muestra para un nivel de significancia del 95%, se calculó mediante la aplicación del programa EPI-Info 6,04d, a partir de la población estimada del municipio de Tumaco (169.000 habitantes), del cual provenían la mayoría de las muestras positivas para HTLV-I Con el nivel de significancia del 95%, el estimado del tamaño de la muestra fue de mínimo 69 casos.
Los niveles de significancia estadística de cada uno de los experimentos se calcularon mediante la aplicación de la prueba exacta de Fisher, además de pruebas estadísticas para la comparación de varianzas.
Para la realización de este trabajo, se solicitó y obtuvo aprobación del Comité de Ética institucional de la Universidad del Valle.
Resultados
Alteración inmune humoral en paraparesia espástica tropical
Cuando se compararon los títulos de IgG, IgM e IgA1 específicas para proteínas del HTLV-I, se determinó que los pacientes con paraparesia espástica tropical examinados, presentaron mayores títulos que los de leucemia de células T del adulto y los seropositivos asintomáticos (p<0,001). La reactividad de plasmas de pacientes con paraparesia espástica tropical a IgM fue estadísticamente mayor que los individuos asintomáticos y los pacientes con leucemia de células T del adulto (p<0,001) (cuadro 1). Las frecuencias IgA1 anti-HTLV-I de pacientes con paraparesia espástica tropical, mostraron diferencias estadísticamente significativas en relación con los portadores asintomáticos y en individuos con leucemia de células T del adulto. En el grupo de los controles negativos no se detectaron anticuerpos anti-HTLV-I en los plasmas de los controles negativos.

En todas las muestras de pacientes infectados con HTLV-I se registraron, mediante análisis de los patrones de Western blot, anticuerpos de la clase IgG contra las proteínas virales gag, p19, p24, env (gp46 y p21) y tax p40. Por el contrario, el porcentaje de bandas reactivas para anticuerpos de la clases IgM fue significativamente mayor en pacientes con paraparesia espástica tropical que en aquéllos con leucemia de células T del adulto (P<0,001) y seropositivos asintomáticos (P<0,001). Los análisis de los patrones de Western blot revelaron la presencia de anticuerpos IgM específicos para las proteínas virales p24, Tax p40 y env (gp46 y p21) solamente.
Marcadores de autoinmunidad en pacientes con paraparesia espástica tropical
De los resultados obtenidos de la aplicación de las pruebas correspondientes, se determinó que aquellas muestras de plasma de pacientes con paraparesia espástica tropical mostraron una mayor frecuencia de reactividad para ANA (70,2%) y ACL-2 (83,8%), en comparación con las de pacientes con leucemia de células T del adulto (P<0,001) y los seropositivos asintomáticos (P<0,001). El cuadro 2 presenta un resumen de los resultados obtenidos de los ensayos realizados para determinar la presencia de autoanticuerpos y marcadores de autoinmunidad en el plasma de los individuos incluidos en el estudio. De los controles negativos para HTLV-I, dos presentaron anticuerpos ANA plasmáticos.
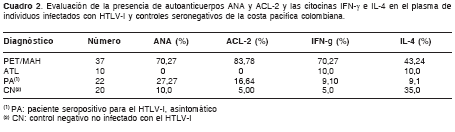
La determinación semicuantitativa de los niveles de IL-4 en pg/ml en plasma mostró los siguientes resultados: controles seronegativos, 2,116 ± 0,008; paraparesia espástica tropical, 3,170 ± 0,09; y leucemia de células T del adulto, 2,120 ± 0,230. De otra parte, los de IFN-g en pg/ml de plasma fueron: controles negativos, 0,045 ± 0,002; paraparesia espástica tropical, 0,360 ± 0,06; y leucemia de células T del adulto, 0,04 ± 0,01. Las diferencias en los niveles de cada una de las citocinas entre controles negativos y casos de paraparesia espástica tropical fueron estadísticamente significativas.
Autoanticuerpos contra células y tejidos del sistema nervioso central humano
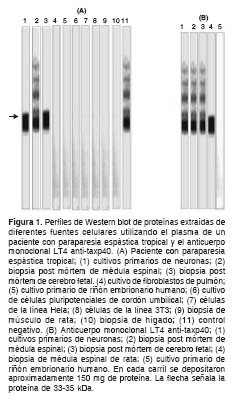
En el plasma de un paciente con paraparesia espástica tropical escogido al azar de la muestra, se observó reacción cruzada contra una proteína de PMr 33-35 kDa obtenida de cultivos primarios de neuronas, de biopsia post mórtem de médula espinal humana, de cerebro fetal y de médula espinal de ratas no infectadas (figura 1a). De otra parte, el anticuerpo monoclonal LT4 anti-taxp40 mostró reactividad cruzada similar a la encontrada en el plasma del pacientes con paraparesia espástica tropical escogido (figura 1b).
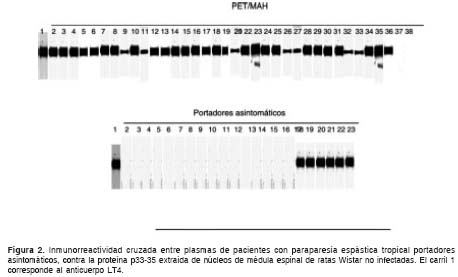
Cuando el experimento se repitió en las muestras incluidas dentro del estudio, se determinó que 91,9% (34/37) de los plasmas de los pacientes con paraparesia espástica tropical y el 27,3% (6/22) de los portadores asintomáticos mostraron reacción cruzada contra la proteína de 33-35 kDa obtenida de cultivos primarios de neuronas, de biopsia post mórtem de médula espinal y de cerebro fetal y de biopsia de médula espinal de rata (figura 2).
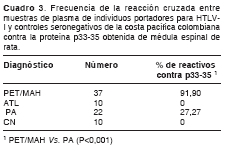
Los valores de las frecuencias de reactividad la proteína 33-35 kDa en los pacientes con paraparesia espástica tropical mostraron diferencias estadísticamente significativas con las de los portadores asintomáticos (P<0,001). Además, no se observó ninguna reacción cruzada en plasmas de los pacientes leucemia de células T del adulto y de los controles seronegativos (cuadro 3).
Localización celular de la reactividad de la p33-35 en médula espinal de rata
El anticuerpo monoclonal LT4 anti-taxp40 y el 91,9% de los plasmas de pacientes con paraparesia espástica tropical, mostraron una intensa reacción cruzada en el núcleo de las neuronas de preparaciones de la región lumbar de la médula espinal de ratas no infectadas (figura 3a). Se determinó una relación directa entre la presencia de la proteína p33-35 de médula espinal de rata y la reacción cruzada observada en el núcleo de neuronas motoras grandes de secciones de médula espinal identificadas por sus largas fibras axonales (figura 3b y figura 3c).
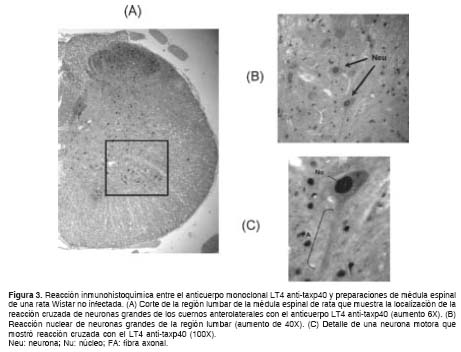
Discusión
En este trabajo se demostró la presencia de IgM e IgA1 específicas para proteínas del HTLV-I con mayor frecuencia en pacientes con paraparesia espástica tropical de la costa pacífica colombiana que en portadores asintomáticos. Los datos obtenidos demuestran que las diferencia de reactividades a las clases de inmunoglobulinas IgM e IgA1 en el plasma de pacientes con esta enfermedad en comparación con la de los portadores asintomáticos y aquéllos con leucemia de células T del adulto (disturbio inmune humoral), serían una característica inmunológica del plasma de los pacientes con paraparesia espástica tropical incluidos en este estudio; además, la mayor frecuencia de anticuerpos anti-HTLV-I de las clases IgM e IgA1, permite postular que el componente humoral juega un papel importante durante la progresión de la enfermedad. Se determinó una reactividad cruzada entre proteínas derivadas de neuronas humanas y de preparaciones de médula espinal de rata con plasmas de individuos colombianos infectados por el HTLV-I; ésta es una característica predominante en los pacientes con paraparesia espástica tropical de la costa pacífica colombiana.
En un estudio previo realizado por nuestro grupo de investigación, se registró una reacción cruzada entre el anticuerpo LT4 anti-Taxp40 y una banda proteica de Mr 33-35 kDa proveniente de porciones de cerebro fetal humano, la cual también se observó en el suero de pacientes con paraparesia espástica tropical provenientes de varias zonas endémicas de Colombia, tanto de la costa pacífica como de la región caribe (18). En el estudio histológico, la reacción cruzada del monoclonal LT4 anti-Taxp40 se localizó en el núcleo de neuronas grandes de las áreas motoras T6 a L2 de la médula espinal de ratas. Así pues, el trabajo inicial mostró la existencia una componente autoinmune en pacientes con paraparesia espástica tropical que coincide con un mimetismo molecular entre dominios de la proteína Tax y los de una proteína de Mr 33-35 proveniente de preparaciones celulares de cerebro fetal humano.
En el presente trabajo, además de confirmar los hallazgos previos, se mostró una reactividad similar entre la proteína nuclear Mr 33-35 kDa de médula espinal de rata y el 100% de los plasmas de pacientes con paraparesia espástica tropical y de algunos individuos seropositivos procedentes exclusivamente de regiones endémicas de la costa pacífica colombiana. La valoración inmuno-histoquímica de los plasmas de pacientes con paraparesia espástica tropical en preparaciones de médula espinal de rata, realizada en este trabajo, confirmó su localización en el núcleo de neuronas motoras grandes. Así pues, tomadas en su conjunto, las evidencias experimentales obtenidas expanden el fenómeno de reacción cruzada previamente descrito en preparaciones de cerebro fetal humano a otros componentes del sistema nervioso central, no sólo en humanos sino en ratas.
Un aporte importante del presente estudio es que reveló la presencia de autoanticuerpos ANA y ACL-2 en el plasma de los pacientes con paraparesia espástica tropical, cuya frecuencia fue significantemente diferente a la observada en los individuos seropositivos asintomáticos procedentes de la costa pacífica colombiana. Además, los niveles plasmáticos de IL-4 e IFN-g medidos en pacientes con paraparesia espástica tropical, mostraron valores significativamente mayores a los registrados para los individuos seropositivos asintomáticos incluidos en este trabajo. Estos resultados permiten postular la existencia de un síndrome autoinmune en aquellos pacientes con paraparesia espástica tropical procedentes de la costa pacífica incluidos en este trabajo.
Con base en nuestros análisis, es posible proponer que la circulación de autoanticuerpos séricos contra proteínas del sistema nervioso humano, sería una característica diferencial de la enfermedad en los pacientes colombianos. Es importante destacar que la circulación de autoanticuerpos que cruzan con proteínas celulares de tejidos normales constituye una evidencia muy fuerte de la existencia de un proceso autoinmune mediado por mimetismo molecular, el cual sería parte de la compleja etiopatogénesis de la paraparesia espástica tropical en estos pacientes.
Un aspecto interesante de este trabajo fue que algunos portadores asintomáticos presentaran la reacción con la proteína p33-35 de médula espinal; en cuanto a esto, una posible explicación es que estos sujetos estuviesen en una fase muy temprana de la enfermedad en la que no se pudo establecer con claridad la sintomatología de la enfermedad. Así, pues, en este sentido es posible proponer que la presencia de autoanticuerpos plasmáticos, precedería al establecimiento de los síntomas que caracterizan la paraparesia espástica tropical. Para validar esta hipótesis es necesario seguir en el tiempo estos sujetos para definir el momento en que desarrollan la sintomatología de paraparesia espástica tropical; sin embargo, esto es otro tipo de estudio.
Para definir de manera más precisa el proceso autoinmune observado en la paraparesia espástica tropical de la costa pacífica colombiana, es importante determinar el tipo de citocinas producidas diferencialmente por las poblaciones de sus linfocitos T ayudadores (Th). Así, la IL-4 es secretada por los linfocitos Th2 mientras que el IFN-g lo es por los Th1. Los resultados obtenidos en este trabajo, mostraron una tendencia a una respuesta preferencial de linfocitos Th1 en el 70,3% de los pacientes con paraparesia espástica tropical; sin embargo, en el 11,3% de ellos se obtuvieron niveles detectables de las dos citocinas. Estos resultados hacen difícil definir el tipo de población de linfocitos Th que sería importante para desencadenar la respuesta autoinmune en dichos pacientes de la costa pacífica. En este sentido, una serie de trabajos previos han favorecido la explicación de que el componente autoinmune es mediado por linfocitos Th1, especialmente en pacientes con paraparesia espástica tropical o mielopatía asociada al HTLV-I del Japón (25-27).
Fue evidente que las reacciones cruzadas observadas en los componentes del sistema nervioso central incluidos en este estudio, muestran la existencia de blancos celulares normales que presentan mimetismo molecular por parte de poblaciones de IgG anti-HTLV-I plasmáticas para ciertas proteínas del virus, específicamente contra la proteína tax-p40. Nuestros resultados confirman aquéllos obtenidos por Levin et al. (1998) (16) y Levin et al. (2002) (17), quienes identificaron una proteína de aproximadamente 35 kDa en neuronas de corteza cerebral normal humana, la cual reacciona en forma cruzada con un anticuerpo monoclonal para la proteína tax del HTLV-I. Sin embargo, un punto importante de nuestro trabajo es que expande el proceso de reacción cruzada de anticuerpos anti-taxp40 no sólo al cerebro sino a regiones de la médula espinal, como la lumbar y la sacra, que jugarían un papel importante en el fisiopatología de la enfermedad en estos pacientes. Por otro lado, el haber determinado esta misma reactividad cruzada en neuronas de la médula espinal de ratas no infectadas, demuestra que la proteína p33-35 estaría en neuronas no sólo de humanos sino en las de la médula espinal de otras especies de mamíferos, en nuestro estudio, la rata.
En aquellas células en las que se identificó la reactividad cruzada entre anticuerpos contra el HTLV-I y proteínas celulares normales, la colocalización de los componentes miméticos coincidió con la distribución intracelular de las proteínas virales que exhiben mimetismo molecular. Tal es el caso de la ubicación nuclear del monoclonal LT4 anti-taxp40. Se ha demostrado que la proteína tax es un activador transcripcional de genes virales y de grupos de genes celulares que participan en diversos eventos celulares (28,29); en este sentido, es importante definir cuál sería la acción de los anticuerpos anti-tax como efectores de la transcripción modulada de estos genes en las neuronas motoras del sistema nervioso central. Nuestros resultados postulan el importante papel que tendría la proteína tax durante la progresión de la enfermedad y el mimetismo molecular. Este hecho sugiere que la acción de las proteínas celulares en la progresión de la paraparesia espástica tropical involucraría mecanismos nucleares, muy posiblemente en la regulación de la expresión de genes celulares implicados en algunos de los eventos, aún no conocidos, de la patogénesis de la enfermedad (29).
Trabajos previos han presentado evidencias de que la hnRNP A1 tiene una reactividad cruzada con anticuerpos monoclonales para la proteína tax p40 del HTLV-I, y muestran que el reconocimiento de este autoantígeno involucra dominios definidos de esta proteína nuclear (30,31). Las ribonucleo-proteínas heterogéneas nucleares (hnRNPs) son las proteínas más abundantes del núcleo de células eucarióticas (32). Entre ellas, hnRNP A1 es una abundante proteína nuclear de células T, que participa en el procesamiento splicing alternativo y estabilización cromosómica, así como en el transporte del núcleo al citoplasma de ribonucleoproteínas mensajeras. Se han obtenido evidencias previas de que hnRNP A1 afecta la regulación de la transcripción de la expresión de genes del HTLV-I, interfiriendo con la unión de la proteína Rex al elemento XRE (33). Concomitantemente, la inhibición de la expresión de hnRNP A1 conlleva al incremento de la transcripción viral junto con un aumento de la producción viral; este hecho sugiere que la proteína hnRNP A1 es parte de los mecanismos reguladores del ciclo de vida de este retrovirus humano en células T (34).
Sin embargo, tal como se ha descrito previamente, hnRNP A1 es una nucleoproteína ubicua que está presente, no solamente en neuronas del sistema nervioso central, sino en otras células no nerviosas incluyendo los linfocitos T. Con base en este hecho, planteamos que debe existir un mecanismo específico del sistema nervioso central subyacente a hnRNP A1, el cual tendría relevancia en desencadenar procesos patológicos que conduzcan a la degeneración axonal observada durante la progresión de la enfermedad.
En su conjunto, los resultados aportaron evidencias de que la paraparesia espástica tropical en aquellos pacientes de la costa pacífica colombiana que se incluyeron en el estudio, se caracterizó como un síndrome autoinmune el cual tiene una respuesta preferiblemente mediada por anticuerpos circulantes mediante un potencial mimetismo molecular. En este sentido, la proteína p33-35, detectada exclusivamente en componentes del sistema nervioso central, sería el posible autoantígeno que participaría en algunos de los procesos que conllevarían a la degeneración axonal característica de esta enfermedad. Sin embargo, es necesario efectuar estudios adicionales de estructura y función, así como descartar otros posibles mecanismos celulares y moleculares de la presencia de reacción cruzada, para poder definir de manera más exacta los procesos complejos que ocurren en la progresión de la paraparesia espástica tropical.
Agradecimientos
A Julio Villalobos por sus sugerencias y críticas al manuscrito, a Yuetzu Tanaka por su donación del anticuerpo monoclonal LT4 anti-taxp40 y a la Vicerrectoría de Investigaciones de la Universidad del Valle.
Conflicto de intereses
Los autores declaramos que no existen conflictos de intereses que puedan influir en forma alguna en los resultados presentados y discutidos en este trabajo.
Financiación
Este trabajo es parte de los resultados obtenidos del proyecto financiado por el Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología «Francisco José de Caldas» (Colciencias), titulado Definición del componente autoinmune en la paraparesia tropical espástica/mielopatía asociada al HTLV-I (PET/HAM)(proyecto 1106-04-13063) y la Universidad del Valle.
Correspondencia:
Felipe García, Laboratorio de Biología Molecular y Patogénesis, Departamento de Ciencias Fisiológicas, Facultad de Salud, Universidad del Valle, Calle 4B Nº 36-00, edificio 116 Jesús María Borrero, oficina 508, sede San Fernando, apartado aéreo 25360, Cali, Colombia. Teléfono: (572) 518 5601; fax: (572) 518 5657. labiomol@gmail.comReferencias
1. Shuh M, Beilke M. The human T-cell leukemia virus type 1 (HTLV-1): new insights into the clinical aspects and molecular pathogenesis of adult T-cell leukemia/lymphoma (ATLL) and tropical spastic paraparesis/HTLV-associated myelopathy (TSP/HAM). Microsc Res Tech. 2005;68:176-96. [ Links ]
2. Vernant JC, Maurs L, Gessain A, Barin F, Gout O, Delaporte JM, et al. Endemic tropical spastic paraparesis associated with human T-lymphotropic virus type I: a clinical and seroepidemiological study of 25 cases. Ann Neurol. 1987;21:123-30. [ Links ]
3. Osame M, Matsumoto M, Usuku K, Izumo S, Ijichi N, Amitani H, et al. Chronic progressive myelophathy associated with elevated antibodies to human T-lymphotropic virus type I and adult T-cell leukemia like cells. Ann Neurol. 1987;21:117-22. [ Links ]
4. Zaninovic V, Galindo J, Blank A. Paraparesia espástica tropical en Colombia. En: Enfermedades asociadas con el virus HTLV-I. Cali: Fundación MAR; 1992. p. 77-86. [ Links ]
5. Bartholomew C, Jack N, Edwards J, Charles W, Corbin D, Cleghorn FR, et al. HTLV-I serostatus of mothers of patients with adult T-cell leukemia and HTLV-I-associated myelopathy/tropical spastic paraparesis. J Hum Virol. 1998;1:302-5. [ Links ]
6. Chávez M, Domínguez MC, Blank A, Quintana M, Eizuru Y, García-Vallejo F. Reconstrucción de la evolución molecular de la infección actual por el virus linfotrópico humano tipo I en Colombia. Biomédica. 2004;24:65-72. [ Links ]
7. Balcázar N, Sánchez G, García-Vallejo F. Sequence and phylogenetic analysis of Human T-Lymphotropic Virus type 1 from Tumaco, Colombia. Mem Inst Oswaldo Cruz. 2003 98:641-8 [ Links ]
8. Trujillo JM, Concha M, Muñoz A, Bergonzoli G, Mora C, Borrero I, et al. Seroprevalence and cofactors of HTLV-I infection in Tumaco, Colombia. AIDS Res Hum Retroviruses. 1992;8:651-7. [ Links ]
9. Nakamura H, Kawakami A, Tominaga M, Hida A, Yamasaki S, Migita K, et al. Relationship between Sjögrens syndrome and human T-lymphotropic virus type I infection: follow-up study of 83 patients. J Lab Clin Med. 2000;135:139-44. [ Links ]
10.Fox RI, Stern M, Michelson P. Update in Sjögren syndrome. Curr Opin Rheumatol. 2000;12:391-8. [ Links ]
11. Beger E, Deocharan E, Edelman M, Erblich B, Gu Y, Putterman C. A peptide DNA surrogate accelerates autoimmune manifestations and nephritis in lupus-prone mice. J Immunol. 2002;168:3617-26. [ Links ]
12.Bangham CR. HTLV-1 infections. J Clin Pathol. 2000;53:581-6. [ Links ]
13. Asquith B, Zhang Y, Mosley AJ, de Lara CM, Wallace DL, Worth A, et al. In vivo T lymphocyte dynamics in humans and the impact of human T-lymphotropic virus 1 infection. Proc Natl Acad Sci USA. 2007; 104:8035-40. [ Links ]
14.Verdonck K, González E, van Dooren S, Vandamme AM, Vanham G, Gotuzzo E. Human T-lymphotropic virus 1: recent knowledge about an ancient infection. Lancet Infect Dis. 2007;7:266-81. [ Links ]
15. Trujillo JR, Mclane MF, Lee TH, Essex M. Molecular mimicry between the human immunodeficiency virus type 1 gp120 V3 loop and human brain proteins. J Virol. 1993;67:7711-5. [ Links ]
16. Levin MC, Krikavsky M, Berck J, Foley S, Rosenfeld M, Dalmau J, et al. Neuronal molecular mimicry in immune-mediated neurologic disease. Ann Neurol. 1998;44:87-98. [ Links ]
17. Levin MC, Lee SM, Kalume F, Morcos Y, Dohan FC Jr, Hasty KA, et al. Autoimmunity due to molecular mimicry as a cause of neurological disease. Nat Med. 2002;8:509-13. [ Links ]
18. García-Vallejo F, Domínguez MC, Tamayo O. Autoimmunity and molecular mimicry in tropical spastic paraparesis/human T-lymphotropic virus-associated myelopathy. Braz J Med Biol Res. 2005;38:241-50. [ Links ]
19. Tanaka Y, Zeng L, Shiraki H, Shida H, Tozawa H. Identification of a neutralization epitope on the envelope gp46 antigen of human T cell leukemia virus type I and induction of neutralizing antibody by peptide immunization. J Immunol. 1991;147:354-60. [ Links ]
20. Lee B, Tanaka Y, Tozawa H. Monoclonal antibody defining tax protein of human T-cell leukemia virus type-I. Tohoku J Exp Med.1989;157:1-11. [ Links ]
21. Burnette WN. Western blotting: Electroforetic transfer of proteins from sodium dodecyl sulfate-polyacrilamide gels to unmodified nitrocellulose and radiographic detection with antibody and radiodinated protein. Anal Biochem. 1981;112:195-203. [ Links ]
22. Barin F, MBoup S, Denis F, Kanki P, Allan JS, Lee TH, et al. Serological evidence for virus related to simian T-lymphotropic retrovirus III in residents of West Africa. Lancet. 1985;2:1387-9. [ Links ]
23. Hsu S, Raine L, Fanger H. Use of avidine-biotine-peroxidase complex (ABC) in immnuperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedure. J. Histochem Cytochem. 1981;29:577. [ Links ]
24. Shi SR, Iman A, Young L, Cote R, Taylor CR. Antigen retrieval immunohistochemistry under the influenza of using monoclonal antibodies. J Histochem Cytochem. 1995; 43:193-201. [ Links ]
25. Ochi H, Wu XM, Osoegawa M, Horiuchi I, Minohara M, Murai H, et al. Tc1/Tc2 and Th1/Th2 balance in Asian and Western types of multiple sclerosis, HTLV-I- associated myelopathy/tropical spastic paraparesis and hyper-IgEaemic myelitis. J Neuroimmunol. 2001; 119: 297-305. [ Links ]
26. Wu X, Osoegawa M, Yamasaki K, Kawano Y, Ochi H, Horiuchi I, et al. Flow cytometric differentiation of Asian and Western types of multiple sclerosis, HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) and hyperIgEaemic myelitis by analyses of memory CD4 positive T cell subsets and NK cell subsets. J Neurol Sci. 2000;177:24-31. [ Links ]
27. Horiuchi I, Kawano Y, Yamasaki K, Minohara M, Furue M, Taniwaki T, et al. Th1 dominance in HAM/TSP and the optico-spinal form of multiple sclerosis versus Th2 dominance in mite antigen-specific IgE myelitis. J Neurol Sci. 2000;172:17-24. [ Links ]
28. Azran I, Schavinsky-Khrapunsky Y, Aboud M. Role of Tax protein in human T-cell leukemia virus type-I leukemogenicity. Retrovirology. 2004;1:20-43 [ Links ]
29. Barmak K, Harhaj E, Grant C, Alefantis T, Wigdahl B. Human T cell leukemia virus type I-induced disease: pathways to cancer and neurodegeneration. Virology. 2003;308:1-12. [ Links ]
30. Levin MC, Lee SM, Morcos Y, Brady J, Stuart J. Cross-reactivity between immunodominant human T lymphotropic virus type I tax and neurons: implications for molecular mimicry. J Infect Dis. 2002;186:1514-7 [ Links ]
31. Lee SM, Dunnavant FD, Jang H, Zunt J, Levin MC. Autoantibodies that recognize functional domains of hnRNPA1 implicate molecular mimicry in the pathogenesis of neurological disease. Neurosci Lett. 2006;401:188-93. [ Links ]
32. Caporali R, Bugatti S, Bruschi E, Cavagna L, Montecucco C. Autoantibodies to heterogeneous nuclear ribonucleoproteins. Autoimmunity. 2005; 38: 25-32. [ Links ]
33. Princler GL, Julias JG, Hughes SH, Derse D. Roles of viral and cellular proteins in the expression of alternatively spliced HTLV-1 pX mRNAs. Virology. 2003;317:136-45. [ Links ]
34.Kress E, Hachem BH, Bex F, Gazzolo L, Duc M. Critical role of hnRNP A1 in HTLV-1 replication in human transformed T lymphocytes. Retrovirology. 2005;2:1-8. [ Links ]

