










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiomédica
Print version ISSN 0120-4157On-line version ISSN 2590-7379
Biomédica vol.29 no.2 Bogotá Apr./June 2009
1 Equipe Microbiologie, Université de Rennes, Rennes, France.
2 Basic Oral Research Unit Institute, Universidad El Bosque, Bogotá, D.C., Colombia
3 Virology Institute, Universidad El Bosque, Bogotá, D.C., Colombia
Recibido: 24/11/08; aceptado:23/02/09Introduction. Porphyromonas gingivalis is considered as a major etiological agent in the onset and progression of chronic destructive periodontitis. Porphyromonus gingivalis fimA type has been correlated to the virulence potential of the strain; therefore this gene could be involved in the ability of P. gingivalis to reach blood stream.
Objective. The classifications of P. gingivalis fimA types will be compared in subgingival plaque and blood samples collected after scaling and root root planing of periodontitis patients.
Materials and methods. Fifteen periodontitis patients requiring scaling and root planing were enrolled. P. gingivalis isolates were classed to genotype with fimA type-specific PCR assay. fimA gene was sequenced if the isolate was listed as unclassifiable after PCR technique.
Results. Six patients showed positive P. gingivalis bacteremia. The most frequent fimA was fimA type II, followed by Ib, III and IV. In blood strains, type II was followed by IV, Ib and III.
Conclusion. Type II was the most frequent genotype in blood samples and in subgingival plaque samples. However, no correlation was found between the frequency of any fimA type with SRP induced bacteremia. P. gingivalis fimA type appears to be conserved within individual patients throughout the times of sample collection. fimA gene sequence results were not in agreement with results of fimA genotyping by PCR.
Key words: Porphyromonas gingivalis, bacteremia, periodontitis, polymerase chain reaction.
Introducción. Porphyromonas gingivalis es el principal agente etiológico de la periodontitis. El gen fimA ha sido relacionado con la virulencia del microorganismo, lo cual sugiere la participación de dicho gen en la capacidad del microorganismo para alcanzar el torrente sanguíneo.
Objetivo. Estudiar la distribución de los tipos de fimA de P. gingivalis en muestras de placa subgingival y de sangre obtenidas durante bacteriemias después de raspaje y alisado radicular.
Materiales y métodos. Se practicó un alisado radicular a 15 pacientes con periodontitis. Se obtuvieron aislamientos clínicos de P. gingivalis de la placa subgingival y durante la bacteriemia inducida por el procedimiento. Para la genotipificación se utilizó la técnica de reacción en cadena de la polimerasa (PCR) específica para fimA. En los aislamientos no clasificables por PCR se realizó secuenciación del gen fimA.
Resultados. Seis pacientes fueron positivos para bacteriemia por P. gingivalis. La distribución de fimA evaluada en 30 aislamientos de placa subgingival y de sangre mostró una mayor frecuencia del fimA tipo II de P. gingivalis. En los aislamientos de placa subgingival, la detección de fimA tipo II fue seguida por Ib, III y IV; sin embargo, en los aislamientos de sangre el tipo II fue seguido por los tipos IV, Ib y III.
Conclusión. En los aislamientos de sangre y de placa subgingival de pacientes con periodontitis el fimA más frecuente fue el tipo II; no fue posible correlacionar el tipo de fimA con la bacteriemia inducida por el alisado radicular. Los resultados de la secuenciación del gen fimA no concuerdan con los obtenidos por PCR.
Palabras clave: Porphyromonas gingivalis, bacteriemia, periodontitis, reacción en cadena de la polimerasa
Periodontal disease (gingivitis and periodontitis) represents a group of infections caused by microorganisms that colonize, modify, penetrate and invade periodontal tissue, destroying connective tissue attachment and alveolar bone, and ultimately leading to tooth loss. Periodontitis has been reported as a public health problem, with approximately 15% of the population suffering from the most severe form (1). Porphyromonas gingivalis is a Gram negative, anaerobic, black pigmented rod. It is an opportunistic periodontal pathogen, and, although it may be absent or decreased in healthy sites, it becomes increased in prevalence and proportion in actively degrading periodontal pockets. Consequently, it has been described as a major etiological agent in the onset and progression of chronic destructive periodontitis. Porphyromonas. gingivalis possesses several virulence factors: fimbriae, hemagglutinins, haemolysin, LPS, proteases, outer membrane vesicles, capsular antigens and ceramides. All these factors are important to fulfill nutritional and growth requirements of P. gingivalis and its virulence potential is likely to be an orchestration among all these components. Fimbriae are filamentous peritrichous structures located on the surface of the bacterium and described as one of the most important virulence factors of P. gingivalis because of their immunological and biological activities (2). Fimbriae are involved in most adherence properties exhibited by P. gingivalis, i.e., the interaction with salivary molecules, oral epithelial cells and other oral bacteria, as well as in the induction of cytokines secretion of infected cells (3-5). The major fimbriae subunit, fimbrillin (FimA), ranges from 41 to 49 kDa (6) and is encoded by the fimA gene. Six variants (I-V and Ib) of the fimA gene have been described based on nucleotide sequences from reference strains and dental plaque samples isolated from patients with or without periodontal disease. Indeed, fimA genotyping may prove useful to identify fimA types involved in adult periodontitis development and to assess risk and assist treatment planning of periodontal disease (7,8).
Several reports have suggested the relationship between periodontal disease and systemic complications (9), such as cardiovascular disease, stroke, lung inflammation and preterm low weight birth (10-13). Periodontal disease can be involved in systemic diseases if periodontopathic bacteria become present in the bloodstream, then leading to extraoral infections. Bacteria associated with transient oral bacteremia can reach the bloodstream after specific dental procedures, chewing, tooth brush or dental flossing (14-17). Once periodontopathic bacteria reach the bloodstream, their virulence can modify host cell functions, inducing the expression of several chronic inflammatory factors, and interfering with regulation of the local inflammatory reaction (18).
The aim of the current study was to determine the distribution of P. gingivalis fimA type in subgingival plaque samples and blood samples collected after scaling and root planing (SRP)-induced transient bacteremia.
Material and methods
Subjects and sample collection
Population. Fifteen unrelated, systemically healthy adults were included in this study. They were selected from subjects who attended the Graduate Clinic of Periodontics and Oral Medicine Service of Universidad El Bosque in Bogotá, Colombia. All subjects were diagnosed with either GCP (generalized chronic periodontitis) or GAP (generalized aggressive periodontitis). Patients must have had at least 10 pockets with probing depth ≥7 mm requiring periodontal surgery after scaling and root planing. All patients were positive for P. gingivalis in subgingival plaque sample before the SRP (scaling and root planning). In addition, laboratory exams, such as a glucose test and haemogram, were made in order to deselect patients with medical compromise. The Ethics Committee of Universidad El Bosque approved the current study, and all patients provided informed consent. All patients were diagnosed according to criteria established by the American Association of Periodontics (AAP) in 1999 (19). Clinical history was evaluated and radiographic examinations were performed for each patient. Periodontal examination included the assessment of pocket depth, loss of attachment and bleeding. The exclusion criteria for were applied as described previously (17).
Samples collection. Samples were collected from subgingival plaque and from blood (17). A subgingival study inclusion sample was collected from 6 pockets (>7 mm). On the surgical day, a second subgingival sample was collected prior to scaling and root planning procedure. The delay between the two samples was one to four weeks. Four blood samples were collected from the antecubital vein at the following time points: (H1) prior to the scaling and root planning procedure; (H2) immediately at the end of the procedure; (H3) 15 minutes and (H4) 30 minutes after the end of the procedure. SRP was assessed during 10 minutes (10 sites, 1 minute per site) (20).
Porphyromonas gingivalis growth conditions and identification. P. gingivalis was cultured on enriched Brucella agar (BBL Microbiology Systems, Cockeysville, Md, USA.) for 6 days. Colony morphology, CAAM (carbobenzoxy-L-arginin-7-amino-4-metylcou-marin, Sigma, St. Louis, Mo, USA) positivity, the commercially available rapid-ID 32A system (Biomerieux), and 16S rRNA based PCR (polymerase chain reaction) using primers described by Ashimoto et al. (21) (Table 1) were performed to identify and confirm P. gingivalis.

Identification of fimA genotypes by PCR. The six primer sets used were based on the primer design described by Amano et al. (22,23) and Nakagawa et al. (8,24) (Table 1). Porphyromonas gingivalis strains ATCC33277 (fimA type I), OMZ409 (provided by Pr Gmür, Zurich; fimA type II), TN (fimA type III), ML2G (fimA type Ib), and W83 (fimA type IV) were used as positive controls. For DNA samples, six-day-old bacterial colonies were added to sterile MiliQ water and boiled. PCR was performed in a final volume of 25 µl containing 10mM dNTPmix, 0.8 µM of each primer, 2mM of MgCl2, 1X of Taq Buffer and 1.25 U of Taq Polymerase Go taq Flexi DNA Polymerase (Promega); 5 µl of the bacterial DNA suspension was added to the PCR reaction, and the final volume was adjusted with sterile MiliQ water. Final concentration of MgCl2 for fimA type II PCR was decreased to 1mM. PCR was performed as follows: an initial denaturation cycle of 95°C for 5 min, 30 cycles of 94°C for 30 s, 58°C for 30 s, 72°C for 30s, and a final extension cycle at 72°C for 7 min, according to previously described methods by Amano et al. (22, 23). Samples showing positive amplification for fimA type I and fimA type II were processed for fimA type Ib. When fimA type Ib amplification was positive, RsaI restriction was performed. Samples showing positive restrictions were classified as fimA type Ib (25). PCR products were resolved by electrophoresis via a 2% agarose Seakem LE gel (Cambrex).
fimA sequencing. For unclassifiable strains, whole fimA gene amplification was undertaken using the universal primer set M1 and M2 (Table 1) and amplifying conditions previously described by Fujiwara et al. (26). The PCR products were purified using the Qiaquick PCR purification kit (Qiagen) following manufacturer instructions. The PCR sequencing of the fimA gene was performed with primers M1 and M2 with the BigDye® Terminator v3.1 Cycle Sequencing Kits following manufacturer instructions. The results were analyzed by an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems).
Nucleotide sequence analysis. Sequences obtained from each isolate were edited and assembled using the ContigExpress tool of the Vector NTI Advance 10 program (Invitrogen). The ClustalW2 multiple sequence alignment program (http://www.ebi.ac.uk/Tools/clustawl2/index.htm) was used to compare similarity between the nucleotide sequence of the fimA gene from clinical strain and the nucleotide sequences from reference strains for fimA type I (Genbank accession no. D17795 (strain 33277)), type II (Genbank accession no.D17799 (strain OMZ409)), type III (Genbank accession no. D17801 (strain BH6/26)), type IV (Genbank accession no. AE015924 (strainW83)), type V (Genbank accession no. AB027294 (strain HNA-99)), and type Ib (Genbank accession no. AB058848 (strain HG1691). In addition, phylogenetic trees were constructed based on the unweighted pair group method with arithmetic mean (UPGMA) analysis to represent relationship between DNA sequences by comparisons based on sequence homology. For the multiple alignment and phylogram construction, the start codon for all the strains was considered to be 5'-ATG-3'(98pb) and the stop codon 5'-TAA-3'(1247pb). The start codon was previously described by Xie et al. (27).
Results
Subjects and P. gingivalis strain classification
Fifteen patients were positively identified for P. gingivalis strains. Nine patients were diagnosed with GCP and 6 with GAP. Independent of the diagnosis, patients were divided in two groups. Group 1 (6 patients) were P. gingivalis-positive in samples from oral cavity (subgingival plaque) and bloodstream (positive bacteremia) (Table 2). The number of P. gingivalis collected in blood was higher 15 minutes after the end of the procedure (4/7) followed by those collected just after the end of the procedure (2/7) and 30 minutes after the end of the procedure (1/7). Group 2 (9 patients) were positive only in subgingival plaque samples (negative bacteremia). Several strains from to each group were lost due to inadequate cryopreservation and marked as "not able to process" (NA) (Table 2).
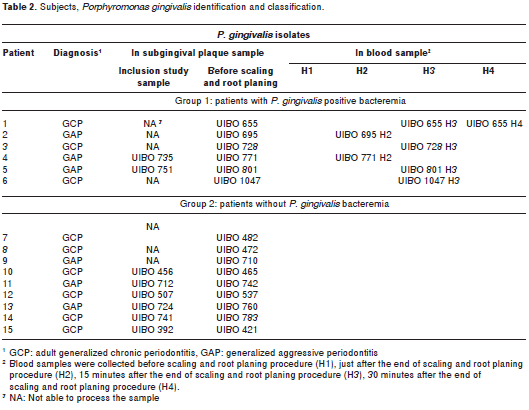
fimA genotypes by PCR
The results of fimA genotype identification from P. gingivalis clinical collection are summarized in Table 3. Genotype assignment was made as follows: strains were classified as a given fimA type when the expected PCR length was produced for one of the fimA types. From a total of 30 P. gingivalis clinical isolates, two were classified as being fimA type I, five as fimA type Ib, fourteen as fimA type II, and two as fimA type III. Strains were listed as unclassifiable when they were positive for more than one fimA type primer sets, as well as after being subjected to whole fimA gene sequencing. Seven strains were unclassifiable (Table 3).
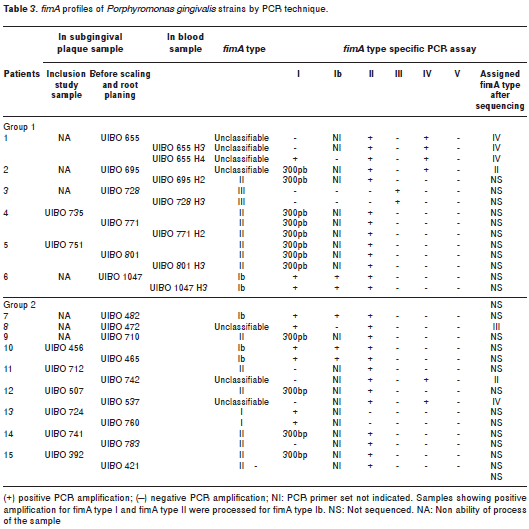
fimA sequencing analysis
Single nucleotide sequences of the entire fimA gene were obtained from 7 strains listed previously as unclassifiable (UIBO 655, UIBO 655 H3, UIBO 655 H4, UIBO 695, UIBO 472, UIBO 742 and UIBO 537). After multiple alignment and phylogenetic tree analysis between the nucleotide sequences of the clinical strains and the sequences of the reference strains for the six fimA types (Figure 1), one fimA type was assigned to each strain. Results are summarized in Table 3. As a whole, isolates UIBO 655, UIBO 655 H3 and UIBO 537 positive for specific primer sets fimA type II and IV PCR were assigned to fimA type IV cluster. Isolate UIBO 655 H4 was positive for specific primer sets fimA type I, II and IV PCR and was assigned to fimA type IV cluster. Strains UIBO 695 and UIBO 742 were positive for specific primer sets fimA type I, II and IV PCR and were assigned to fimA type II cluster. Strain UIBO 472 was positive for specific primer set fimA type I, and II PCR and was assigned to fimA type III cluster.
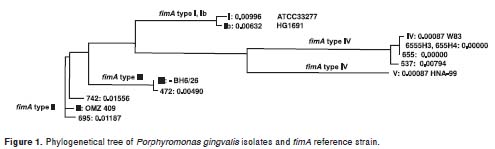
Frequency of fimA genotypes
The esults from fimA genotyping by PCR and nucleotide sequence analysis of fimA whole gene can be summarized as follows: three were classified as being fimA type II, two as fimA type IV, one as fimA type Ib, one as fimA type III. All P. gingivalis strains found in the bloodstream showed the same fimA type as the isolate from the same day SRP in patients 1-6 (Table 3, Table 4).
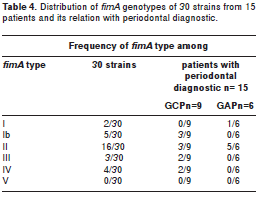
In group 2 (negative bacteremia) fimA type identifications of P. gingivalis were as follows: eight strains were fimA type II, three were fimA type Ib, two were fimA type I, one was fimA type III and one was fimA type IV. In samples collected from one patient, the fimA genotypes found among two strains (2/30) (12) were not the same.
Analysis summarizing fimA identifications in the study population and the relationship between fimA type and periodontal diagnostics are listed in Table 4. Results reveal the high frequency of fimA type II in the total of the strains (17/30), in the total of patients (9/15) and especially in those diagnosed with GAP (5/15).
Discussion
Classification of fimA genotypes identified in strains of Colombian periodontitis patients shows that type II is the most frequent type, as is consistent with Amano et al. (22,23), Nakagawa et al. (8), Missailidis et al. (25) and Eick et al. (28).Those results were obtained in large (73 patients, (23)) and small samples (15 patients, (28)) of periodontitis patients. The second most frequent type in the current study was type Ib. This again was in agreement with the fimA Ib distribution in periodontitis patients in the Brazilian population described by Missailidis et al. (25) and in an Asian population described by Nakagawa et al. (8) where type Ib was strongly associated with periodontitis disease. Type I, III and IV were detected less frequently, similar to the occurrence rate in periodontitis patients in the Brazilian population (25), where fimA type I, III and IV showed a similar frequency, although less frequently detected than fimA type II and Ib. Type V was not detected in the Colombian population. The absence of fimA type V was probably because type V is detected mainly in the healthy population (22). Moreover, type V was absent in gingivitis and periodontitis patients (25). Concerning the relation between fimA type and periodontitis aggression, fimA type II may exhibit an increased pathogenic potential, because it was the most frequent type among GAP patients. These findings are in agreement with previous studies that have shown that fimA type II, IV and Ib strains exhibit a more virulent potential, while fimA type I, III and V are referred as less virulent or avirulent (8,22,24,25,29).
fimA type identifications of P. gingivalis isolated from blood samples collected after orally induced bacteremia by SRP procedures showed a wide heterogeneity (type II 3/7, type IV 2/7, type Ib1/7, type III 1/7). This heterogeneity enabled us to associate a particular fimA type with the ability of the bacteria to reach bloodstream.
In both groups, genotyping data of isolates within individual patients showed the same fimA type. This finding indicated that the fimA genotype was conserved during the sample period of the study and also that one fimA type was unique to a single patient. However, in group 2 (negative bacteremia), fimA detection in two samples in the same patient shows that two allelic variants of P. gingivalis. This result is in agreement with those previously reported by Eick et al. in a German population (28) and Enersen et al. (30).
The presence of unclassifiable strains that has been reported in Brazilian (25), Caucasian (31) and Japanese population (22) suggested that other fimA types associated with periodontitis disease flora may be present in the group under investigation. In order to validate this hypothesis, the entire fimA gene of the seven strains was sequenced. However, the fimA nucleotide sequences obtained showed high homology with nucleotide sequence of the references strains. Therefore, no new fimA types were evidenced. Nevertheless, fimA gene sequences of isolates were not in accordance with the specific fimA type combinations amplifications, supporting previous findings by Ernersen et al. (30).
One unique datum from the current study was discovered in contrast to previous studies (8,22-24). When assessing fimA type I, some strains (12/30) showed a single amplicon (300 bp) not in accordance with the described 392 bp length product. A bioinformatic analysis of designated primers revealed that the fimA type IV reverse primer annealed with an intergenic region, as shown in the sequenced P. gingivalis genome of strain W83 [type IV fimA reference strain (32) GenBank accession number AE015924]. This information was confirmed with the recent whole genome sequence of P. gingivalis ATCC 33277 (33). Considering that reverse primer annealing was common for fimA type IV, III, II and I, these cases were assumed to be annealing with an intergenic region. Therefore, the presence of the 300 bp product suggested that the type I fimA primers set can hybridize elsewhere in the genome of clinical strains.
In sum, fimA type II was assumed to be the most frequently detected type in Colombian periodontitis patients, and specifically in patients exhibiting GAP. Although fimA type II is the most frequent type of P. gingivalis isolated from the bloodstream (3/7), due to the heterogeneity of those strains, the frequency of any fimA type cannot be correlated with the ability of P. gingivalis strains to reach the bloodstream. The number of P. gingivalis strains per person in the Colombian periodontitis group is mostly restricted to one type. The P. gingivalis strain seems to be conserved within a period of one to four weeks, as revealed by genotyping data obtained from inclusion sampling and treatment sampling. In addition, the P. gingivalis strain found in the subgingival plaque sample was often correlated to P. gingivalis found in bloodstream.
We are grateful to Rudolf Gmür, Institute of Oral Biology Section of Oral Microbiology and General Immunology, University of Zürich for kindly providing the OMZ strains; Céline Allaire (Université de Rennes 1); Philippe Gautier, Laboratoire de Bactériologie, Centre Hospitalier Universitaire, (Rennes); Diana Marcela Castillo, María Rosario Aya (Basic Oral Research Unit Institute, El Bosque University), and Germán E. Pérez (Universidad Nacional de Colombia).
This paper was translated by San Francisco Edit.
No conflict of interest.
This study was supported by the Langlois Foundation, Regional Council of Brittany, Investigation Division El Bosque University, COLCIENCIAS - Grant 1308-04-11854 and Ecos-Nord.
Corresponding: Martine Bonnaure-Mallet, Equipe de Microbiologie EA 1254, Université de Rennes 1, 2 avenue du Professeur Léon Bernard, 35043 Rennes, France. Tel: 332 23234358; fax: 332 23234913. martine.bonnaure@univ-rennes1.fr
1. Burt B. Position paper: epidemiology of periodontal diseases. J Periodontol. 2005;76:1406-19. [ Links ]
2. Hamada S, Amano A, Kimura S, Nakagawa I, Kawabata S, Morisaki I. The importance of fimbriae in the virulence and ecology of some oral bacteria. Oral Microbiol Immunol. 1998;13:129-38. [ Links ]
3. Eskan MA, Hajishengallis G, Kinane DF. Differential activation of human gingival epithelial cells and monocytes by Porphyromonas gingivalis fimbriae. Infect Immun. 2007;75:892-8. [ Links ]
4. Hajishengallis G, Sharma A, Russell MW, Genco RJ. Interactions of oral pathogens with toll-like receptors: possible role in atherosclerosis. Ann Periodontol. 2002;7:72-8. [ Links ]
5. Khlgatian M, Nassar H, Chou HH, Gibson FC 3rd, Genco CA. Fimbria-dependent activation of cell adhesion molecule expression in Porphyromonas gingivalis-infected endothelial cells. Infect Immun. 2002;70:257-67. [ Links ]
6. Lee JY, Sojar HT, Bedi GS, Genco RJ. Porphyromonas (Bacteroides) gingivalis fimbrillin: size, amino-terminal sequence, and antigenic heterogeneity. Infect Immun. 1991;59:383-9. [ Links ]
7. Koehler A, Karch H, Beikler T, Flemmig TF, Suerbaum S, Schmidt H. Multilocus sequence analysis of Porphyromonas gingivalis indicates frequent recombination. Microbiology. 2003;149:2407-15. [ Links ]
8. Nakagawa I, Amano A, Ohara-Nemoto Y, Endoh N, Morisaki I, Kimura S, et al. Identification of a new variant of fimA gene of Porphyromonas gingivalis and its distribution in adults and disabled populations with periodontitis. J Periodontal Res. 2002;37:425-32. [ Links ]
9. Lourbakos A, Potempa J, Travis J, D'Andrea MR, Andrade-Gordon P, Santulli R, et al. Arginine-specific protease from Porphyromonas gingivalis activates protease-activated receptors on human oral epithelial cells and induces interleukin-6 secretion. Infect Immun. 2001;69:5121-30. [ Links ]
10. Cueto A, Mesa F, Bravo M, Ocana-Riola R. Periodontitis as risk factor for acute myocardial infarction. A case control study of Spanish adults. J Periodontal Res. 2005;40:36-42. [ Links ]
11. Offenbacher S, Boggess KA, Murtha AP, Jared HL, Lieff S, McKaig RG, et al. Progressive periodontal disease and risk of very preterm delivery. Obstet Gynecol. 2006;107:29-36. [ Links ]
12. Renvert S, Pettersson T, Ohlsson O, Persson GR. Bacterial profile and burden of periodontal infection in subjects with a diagnosis of acute coronary syndrome. J Periodontol. 2006;77:1110-9. [ Links ]
13. Scannapieco FA, Stewart EM, Mylotte JM. Colonization of dental plaque by respiratory pathogens in medical intensive care patients. Crit Care Med. 1992;20:740-5. [ Links ]
14. Forner L, Larsen T, Kilian M, Holmstrup P. Incidence of bacteremia after chewing, tooth brushing and scaling in individuals with periodontal inflammation. J Clin Periodontol. 2006;33:401-7. [ Links ]
15. Heimdahl A, Hall G, Hedberg M, Sandberg H, Soder PO, Tuner K, et al. Detection and quantitation by lysis-filtration of bacteremia after different oral surgical procedures. J Clin Microbiol. 1990;28:2205-9. [ Links ]
16. Kinane DF, Riggio MP, Walker KF, MacKenzie D, Shearer B. Bacteraemia following periodontal procedures. J Clin Periodontol. 2005;32:708-13. [ Links ]
17. Lafaurie GI, Mayorga-Fayad I, Torres MF, Castillo D, Aya MR, Baron A, et al. Periodontopathic microorganisms in peripheric blood after scaling and root planing. J Clin Periodontol. 2007;34:873-9. [ Links ]
18. Bodet C, Chandad F, Grenier D. Modulation of cytokine production by Porphyromonas gingivalis in a macrophage and epithelial cell co-culture model. Microbes Infect. 2005;7:448-56. [ Links ]
19. Armitage GC. Development of a classification system for periodontal diseases and conditions. Ann Periodontol. 1999;4:1-6. [ Links ]
20. Perez-Chaparro PJ, Gracieux P, Lafaurie GI, Donnio PY, Bonnaure-Mallet M. Genotypic characterization of Porphyromonas gingivalis isolated from subgingival plaque and blood sample in positive bacteremia subjects with periodontitis. J Clin Periodontol. 2008;35:748-53. [ Links ]
21. Ashimoto A, Chen C, Bakker I, Slots J. Polymerase chain reaction detection of 8 putative periodontal pathogens in subgingival plaque of gingivitis and advanced periodontitis lesions. Oral Microbiol Immunol. 1996;11:266-73. [ Links ]
22. Amano A, Kuboniwa M, Nakagawa I, Akiyama S, Morisaki I, Hamada S. Prevalence of specific genotypes of Porphyromonas gingivalis fimA and periodontal health status. J Dent Res. 2000;79:1664-8. [ Links ]
23. Amano A, Nakagawa I, Kataoka K, Morisaki I, Hamada S. Distribution of Porphyromonas gingivalis strains with fimA genotypes in periodontitis patients. J Clin Microbiol. 1999;37:1426-30. [ Links ]
24. Nakagawa I, Amano A, Kimura RK, Nakamura T, Kawabata S, Hamada S. Distribution and molecular characterization of Porphyromonas gingivalis carrying a new type of fimA gene. J Clin Microbiol. 2000;38:1909-14. [ Links ]
25. Missailidis CG, Umeda JE, Ota-Tsuzuki C, Anzai D, Mayer MP. Distribution of fimA genotypes of Porphyromonas gingivalis in subjects with various periodontal conditions. Oral Microbiol Immunol. 2004;19:224-9. [ Links ]
26. Fujiwara T, Morishima S, Takahashi I, Hamada S. Molecular cloning and sequencing of the fimbrilin gene of Porphyromonas gingivalis strains and characterization of recombinant proteins. Biochem Biophys Res Commun. 1993;197:241-7. [ Links ]
27. Xie H, Lamont RJ. Promoter architecture of the Porphyromonas gingivalis fimbrillin gene. Infect Immun. 1999;67:3227-35. [ Links ]
28. Eick S, Rodel J, Einax JW, Pfister W. Interaction of Porphyromonas gingivalis with KB cells: comparison of different clinical isolates. Oral Microbiol Immunol. 2002;17:201-8. [ Links ]
29. Davila-Perez C, Amano A, Alpuche-Solis AG, Patino-Marin N, Pontigo-Loyola AP, Hamada S, et al. Distribution of genotypes of Porphyromonas gingivalis in type 2 diabetic patients with periodontitis in Mexico. J Clin Periodontol. 2007;34:25-30. [ Links ]
30. Enersen M, Olsen I, Kvalheim O, Caugant DA. fimA genotypes and multilocus sequence types of Porphyromonas gingivalis from patients with periodontitis. J Clin Microbiol. 2008;46:31-42. [ Links ]
31. Beikler T, Peters U, Prajaneh S, Prior K, Ehmke B, Flemmig TF. Prevalence of Porphyromonas gingivalis fimA genotypes in Caucasians. Eur J Oral Sci. 2003;111:390-4. [ Links ]
32. Nelson KE, Fleischmann RD, DeBoy RT, Paulsen IT, Fouts DE, Eisen JA, et al. Complete genome sequence of the oral pathogenic Bacterium Porphyromonas gingivalis strain W83. J Bacteriol. 2003;185:5591-601. [ Links ]
33. Naito M, Hirakawa H, Yamashita A, Ohara N, Shoji M, Yukitake H, et al. Determination of the genome sequence of Porphyromonas gingivalis strain ATCC 33277 and genomic comparison with strain W83 revealed extensive genome rearrangements in P. gingivalis. DNA Res. 2008;15:215-25. [ Links ]

