










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiomédica
Print version ISSN 0120-4157On-line version ISSN 2590-7379
Biomédica vol.29 no.3 Bogotá July./Sept. 2009
1 Centro de Investigación en Enfermedades Tropicales (CINTROP), Facultad de Salud, Escuela de Medicina, Departamento de Ciencias Básicas, Universidad Industrial de Santander, Piedecuesta, Santander, Colombia.
2 Department of Infectious and Tropical Diseases, London School of Hygiene and Tropical Medicine, London, United Kingdom
Recibido: 28/11/08; aceptado:08/04/09
Introduction. The current chemotherapy for Chagas disease is unsatisfactory with only two drugs available for treatment. Research to discover new drugs for Chagas disease is urgent. Hexadecyl-phosphocholine (HPC, miltefosine) has been demonstrated to have in vitro activity against Trypanosoma cruzi parasites, but its activity on different Colombian T. cruzi strains is not known.
Objective. To evaluate the in vitro susceptibility of T. cruzi strains isolated from humans and vectors in Santander, Colombia. to miltefosine, nifurtimox and benznidazole.
Materials and methods. Eight T. cruzi Colombian strains and three reference strains (Esmeraldo, SilvioX10 and Y) were studied. Drug activities against extracellular epimastigotes and intracellular amastigotes were determined by microscopic counting. The results were expressed as the concentrations that inhibited 50% and 90% growth (IC50 and IC90).
Results. For miltefosine a similar range of drug activity was observed against all the Colombian strains, all parasites being more susceptible to miltefosine than to the reference drugs. The intracellular amastigotes were more susceptible to miltefosine (IC50 0.08 to 0.63 µM and IC90 0.21 to 2.21 µM) than extracellular forms (IC50 <0.92 to 2.29 µM and IC90 1.38 to 4.76 µM). For reference drugs, parasites were more susceptible to nifurtimox than to benznidazole and some differences in activity of benznidazole between T. cruzi strains was observed.
Conclusions. The results showed the significant in vitro activity of miltefosine against T. cruzi stages, and the expected results for the reference drugs. Further in vivo studies with miltefosine are planned.
Key words: Trypanosoma cruzi, Chagas disease, miltefosine, benznidazole, nifurtimox, drug therapy, Colombia.
Susceptibilidad in vitro a hexadecilfosfocolina (miltefosina), nifurtimox y benznidazole de cepas de Trypanosoma cruzi aisladas en Santander, Colombia
Introducción. Los tratamientos actuales para la enfermedad de Chagas son insatisfactorios y sólo existen dos medicamentos disponibles. La búsqueda de alternativas terapéuticas es prioritaria. La hexadecilfosfocolina (miltefosina) ha mostrado actividad in vitro contra Trypanosoma cruzi. Sin embargo, su actividad en aislamientos de T. cruzi obtenidos en Colombia aún no ha sido reportada.
Objetivo. Evaluar la susceptibilidad in vitro a miltefosina, nifurtimox y benznidazole de cepas de T. cruzi aisladas de humanos y vectores en Santander, Colombia.
Materiales y métodos. Se evaluó la susceptibilidad de los tres medicamentos en ocho cepas colombianas de T. cruzi y tres cepas de referencia: Esmeraldo, Silvio X10 y Y. La actividad de los compuestos fue determinada en epimastigotes extracelulares y amastigotes intracelulares, por conteo microscópico. Los resultados se expresaron en concentraciones inhibitorias 50 y 90 (CI50 y CI90).
Resultados. Para la miltefosina, se observaron rangos similares en la actividad del medicamento entre las cepas colombianas; todos los parásitos fueron más susceptibles a la miltefosina que a los medicamentos de referencia. Los amastigotes intracelulares fueron más sensibles a la miltefosina (CI50, 0,08 a 0,63 µM y CI90, 0,21 a 2,21 µM) que las formas extracelulares (CI50, <0,92 a 2,29 µM y CI90, 1,38 a 4,76 µM). En los medicamentos de referencia, los parásitos fueron más susceptibles al nifurtimox que al benznidazole. Se observaron algunas diferencias en la actividad del benznidazole en las cepas estudiadas de T. cruzi.
Conclusiones. Los resultados obtenidos de la actividad in vitro de miltefosina y de los medicamentos de referencia contra aislamientos de T. cruzi son satisfactorios y serán considerados en estudios posteriores in vivo.
Palabras clave: Trypanosoma cruzi, enfermedad de Chagas, miltefosina, benznidazol, nifurtimox, quimioterapia, Colombia.
Chagas disease, caused by the protozoan parasite Trypanosoma cruzi (Kinetoplastida: Trypanosomatidae), constitutes one of the largest parasitic disease burdens in Latin America where approximately 13-15 million of people are affected (1). Trypanosoma cruzi strains are divided into two main well-defined groups called T. cruzi I and T. cruzi II; five subgroups have been identified inside the T. cruzi II group (IIa, IIb, IIc, IId and IIe) (2). In Colombia, 5% of the population is infected in endemic areas and the predominance of T. cruzi I has been reported (3-7). The Santander Province is one of the territorial divisions where the prevalence of infection is highest (8,9). No vaccines are available, and the chemotherapy remains precarious with only two available drugs, the nitrofuran derivative, nifurtimox (Lampit, Bayer) and 2-nitroimidazole benznidazole (Radanil, Roche). Both are restricted mainly to the treatment of the acute phase or the congenital infection (10,11). Currently, these drugs have severe limitations, including high frequency of undesirable side effects, long protocols of treatment, and limited efficacy and availability (12,13). The research to discover new drugs for the treatment of Chagas disease is imperative.
Hexadecylphosphocholine (HPC; miltefosine), an alkylphosphocholine developed initially as an anticancer drug, constitutes the first oral treat-ment for visceral and cutaneous leishmaniasis (14,15). It shows a broad spectrum of drug activity on several types of parasites such as Leishmania spp., T. cruzi and Entamoeba histolytica (16). In T. cruzi models, it is active against free and intracellular live forms of parasites as well as strains with different levels of susceptibility to benznidazole (17,18). Its mechanism of action is not clear, but parasite membranes are probably the primary target, possibly by interference with lipid metabolism (18,19). Additionally, synergism of miltefosine or edelfosine (another alkyl-phospholipid drug) in combination with ketoconazole has also been demonstrated (18-20).
The purpose of the current study was to evaluate the in vitro activities of miltefosine, nifurtimox and benznidazole against Colombian T. cruzi strains, as well as against the Silvio X10, Esmeraldo and Y reference strains.
Materials and methods
Parasites
Eight strains of T. cruzi, isolated from various hosts by hemoculture and xenoculture, were used (Luna KP, Jaramillo CL, Gutierrez R, Esteban L, Angulo VM. Aislamiento de T. cruzi en pacientes en fase crónica de la enfermedad de Chagas por medio de hemocultivo y xenocultivo. Biomédica. 2003;23 (Supl.1):119). They have been previously characterized by amplification of the mini-exon gene nontranscribed spacer (Gutierrez R, Luna KP, Angulo VM, Fernandes O. Eficiencia de los métodos parasitológicos de diagnóstico y caracterización de poblaciones de Trypanosoma cruzi en un area endémica de Chagas. Biomédica. 2003;23 (Supl.1):119-20). Parasites originated from the domestic cycle in the biogeographical area of Santander, Colombia (Table 1). In addition, T. cruzi reference strains Silvio X10 (T. cruzi I) Esmeraldo and Y (T. cruzi II) were used. The parasites were maintained by serial passage in Liver Infusion Tryptose (LIT) medium supplemented with 10% heat inactivated fetal calf serum (hiFCS, Gibco, USA) and hemin (10 mg/L, Sigma) at 28 ºC. They were used within four passages of isolation.
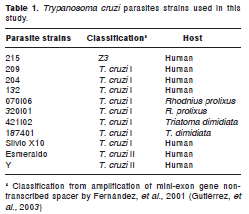
Drugs
Hexadecylphosphocholine (HPC, miltefosine) was obtained from A.G Scientific, INC, nifurtimox from Bayer (Germany) and benznidazole from Roche (Switzerland). Nifurtimox and benznida-zole were dissolved in dimethyl sulfoxide (DMSO). The final DMSO concentration did not exceed 1% (v/v). Miltefosine was dissolved directly in culture medium. Working solutions were prepared in LIT or RPMI 1640 culture media immediately before the assays.
Epimastigotes assay
Parasites were harvested during the exponential growth phase, diluted in LIT to 5x105 parasites/mL and incubated in a three-fold dilution series of miltefosine (0.92-25.0 µM), nifurtimox (1.1-261 µM) and benznidazole (1.2-288 µM) in 96 well multiwell plates (Becton Dickinson, New Jersey, USA), for 72 h at 28 ºC. Each drug concentration was evaluated in triplicate and control cultures were maintained without drug. Parasite inhibition was determined microscopic-ally by counting parasite numbers in a Neubauer haemocytometer.
Intracellular amastigotes assay
Vero cells, diluted to 3 x 105 cells/mL in RPMI 1640 medium plus 10% hiFCS, were plated in 16-well Lab-tekTM tissue culture chamber slides (Life Technologies, Paisley,UK) and allowed to adhere for 24 h at 37 ºC in a 5% CO2-95% air mixture. Adherent cells were infected with tissue derived trypomastigotes (TDT) at a ratio of parasites to cells of 10:1. The cultures were maintained in a 5% CO2-95% air mixture at 37 ºC. TDT were obtained by successive serial infections in Vero cells with released parasites from culture supernatants. Reference strains and the 215 strain were not tested in the amastigote assay due to a failure to generate sufficient TDT to infect cells. After 24 h, free parasites were removed by washing and infected cultures were incubated for 72 h with three fold dilution series of miltefosine (0.04 to 3.30 µM), nifurtimox (0.3-87.0 µM) and benznidazole (1.2 to 96.0 µM) for 72 h at 37 ºC, in a 5% CO2-95% air mixture. The cultures were then fixed with methanol and stained with Giemsa. Each drug concentration was evaluated in quadruplicate and control cultures were maintained without drug. Drug activity was determined by the percentage of infected cells in treated and untreated cultures.
Statistical analyses
The drug activities were expressed as the drug concentration needed to inhibit to 50% or 90% of parasite growth (IC50 and IC90). They were calculated by sigmoidal regression analysis (MsxlfitTM; ID Business Solution, Guildford, UK). Results were expressed as mean ± standard deviation (SD) and statistical significance was determined by Student t test. Values of p <0.05 were considered significant. Each experiment was repeated twice.
Results
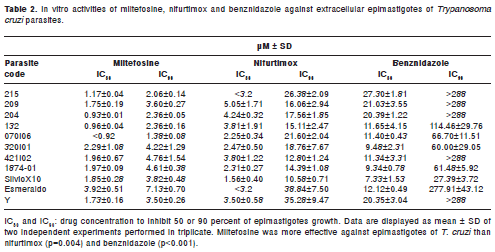
The activities of miltefosine, nifurtimox, and benznidazole against T. cruzi epimastigotes are shown in table 2. All Colombian T. cruzi strains were more susceptible to miltefosine (IC50 <0.92 to 2.29 ± 1.08 µM and IC90 1.38 ± 0.08 to 4.76 ± 1.54 µM) and nifurtimox (IC50 2.25 ± 0.34 to 5.05 ± 1.70 µM and IC90 12.80 ± 1.24 to 26.38 ± 2.09 µM) than to benznidazole (IC50 9.34 ± 0.78 to 27.30 ± 1.81 µM and IC90 60.00 ± 29.05 to >288 µM). Reference strains showed considerable variation in drug sensitivity: Esmeraldo was the least susceptible to miltefosine (IC50 3.92 ± 0.51 µM and IC90 7.13 ± 0.70 µM) and Y was the least susceptible to benznidazole (IC50 20.35 ± 3.04 µM and IC90 >288 µM). The results were reproducible in the two independent experiments.
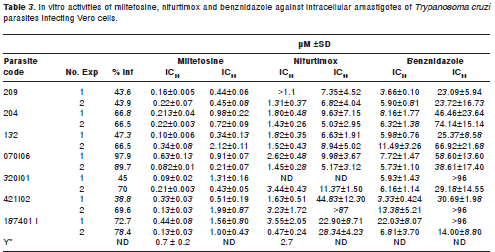
The activity range of miltefosine, nifurtimox and benznidazole against T. cruzi intracellular amastigotes is shown in table 3. All T. cruzi strains were highly susceptible to miltefosine (IC50 0.082 ± 0.01 to 0.63 ± 0.13 µM and IC90 0.21 ± 0.70 to 2.21 ± 0.07 µM). Nifurtimox and benznidazole were also active against intracellular amastigotes but with lower values than miltefosine.
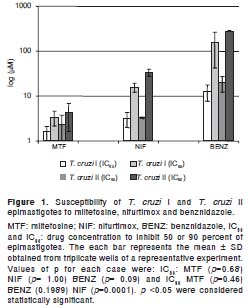
Variation in the activity of miltefosine against each life cycle stage was observed. Intracellular amastigotes were more sensitive than epimastigotes (p=0.0000) (tables 2 and 3). Additionally, comparisons of values of IC50 and IC90 between genetic groups (T. cruzi I and T. cruzi II) showed no significant difference, with one except for nifurtimox which showed significant differences in IC90 values (p=0.0001) (figure 1).
Discussion
Differences in the drug susceptibility of T. cruzi strains according to genetic group, host and geographical origin have been described (21-23). However, investigations performed to date have mainly evaluated susceptibilities to benznidazole and nifurtimox. Both of these standard drugs have limited efficacy in the chronic form of Chagas disease and they produce toxic side effects (12). Therefore, there is an urgent need for studies that evaluate the activity of new anti-trypanosomal drugs.
This is the first study that shows the in vitro susceptibility to miltefosine of T.cruzi strains isolated from humans and vectors in a Colombian endemic area. All the parasites strains and stages tested were highly susceptible to miltefosine. Similar results were obtained by other authors using T. cruzi reference strains confirming the broad activity of this drug (17-19,24). Miltefosine was significantly more active against intracellular amastigotes than epimastigotes (p<0.001) (table 3). This has been demonstrated in earlier studies as well (17,19). Initially, phospholipid-derived drugs were discovered as a new class of biological response modifiers, with therapeutic activity in cancer mediated by an increase in the cytotoxic properties of macrophages (25).
Saraiva et al. (2002) observed that miltefosine may activate macrophages in vitro and the antiparasitic activity of this compound on intra-cellular amastigotes was independent of nitric oxide production (17). The participation of some cytokines was suggested. However, the host cell mechanisms involved in the inhibition or parasite killing after miltefosine treatment is still unclear.
Against Leishmania parasites, a significant variation of the in vitro susceptibility to miltefosine has been described for several species of Leishmania (26). In patients with cutaneous leishmaniasis, it is more effective in lesions caused by L. panamensis as compared to those by L. braziliensis (27). In contrast with the results obtained in the present study, miltefosine was found to be more active against promastigotes of Leishmania than against intracellular amastigotes infecting bone marrow derived macrophages (26).This difference with T. cruzi may be due to intrinsic differences of both parasites. However, Croft (2003) proposed that in comparative studies between extracellular and intracellular stages one must consider that: (i) the effective drug concentration in host cells may be higher or lower than that outside the cells, (ii) the division rates of the stages differ, or (iii) the assay temperatures may differ (16).
In experimental in vivo models, miltefosine has also proved to be active against Leishmania and T. cruzi when it was administered orally (17,21,28,29). Unfortunately, only two studies with T. cruzi have been reported; both showed a suppressive effect against T. cruzi on BALB/c-infected mice, although they used different protocols of treatment and infection (17,21). Further research in a murine or another more appropriate in vivo model is necessary to determine aspects such as pharmacokinetic properties, dose-response relationship and protocols of treatment.
We also investigated the susceptibility of T. cruzi strains to two nitroheterocyclic compounds (nifurtimox and benznidazole). Although all strains were susceptible to both drugs, we noted that nifurtimox was more active than benznidazole both in epimastigotes (p<0.001) and intracellular amastigotes (P=0.0004). Benzni-dazole showed greater variation in its activity against different Colombian T. cruzi strains. This could be related to the different mechanisms of action of both drugs (30,31). Additionally, in contrast to the results obtained with miltefosine, the epimastigotes of strains isolated from vectors were more susceptible to nifurtimox (p=0.03) and benznidazole (p= 0.01) than those isolated from humans (table 2). Several studies have reported a wide variation in the susceptibility of a large number of T. cruzi strains isolated from different hosts and geographic areas to these nitroheterocyclic compounds (22,23,32-34). The existence of predominant strains with particular levels of susceptibility to these drugs in specific geographical areas has been also reported (22,23). In these cases, the parasite drug susceptibility before clinical treatment must be considered as a factor. Moreover, for a better understanding of the response to drugs, the local strains should be characterized genetically and biologically, as completed in this study (Luna KP, Jaramillo CL, Rueda CM, Hernández IP, Zorro MM, Angulo VM, et al. Biological and genetic characteristics of Trypanosoma cruzi isolates from an endemic area of Santander, Colombia. Bol Malariol Sal Amb. 2007;47 (Supl 1):139-40).
Many authors have explored possible links between the phylogenetic diversity of T. cruzi and biological properties (24,35-38). Correlations between the susceptibility to benznidazole and genetic groups of T. cruzi have been described (24,38). However, in the current study, no significant differences were observed in the drug susceptibility in vitro between T. cruzi I and T. cruzi II strains (figure 1). This is in agreement with Villareal et al. (2004) (39), although due to the low number of strains analyzed in the current work, the results obtained must be considered as preliminary data.
In conclusion, the current in vitro study described the susceptibility to miltefosine, nifurtimox and benznidazole of T. cruzi strains isolated from humans and vectors in the same transmission area of Chagas disease in Colombia. Miltefosine showed greater activity than the used reference drugs and no differences in the susceptibility according to host origin were observed. These were considered promising results. Further investigation with these strains in murine models of experimental Chagas disease will be necessary to establish the potential of miltefosine for the treatment of human Chagas disease before undertaking clinical studies.
The authors declare that there are no conflicts of interest on the results published in this paper.
This work was supported by the Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnologia "Francisco Jose de Caldas" Colciencias (Grant 1102-04-12928) and by the Universidad Industrial de Santander, Bucaramanga, Colombia.
Correspondence:
Patricia Escobar, CINTROP, Facultad de Salud, Escuela de Medicina, Departamento de Ciencias Básicas, Universidad Industrial de Santander, Bucaramanga, Colombia. Teléfono/fax: (0057) 656 3971 pescobar@uis.edu.co
1. World Health Organization. Tropical disease research, progress 2003-2004: 17th Programme Report of the UNICEF/UNDP/World Bank/WHO. Special Programme for Research and Training in Tropical Diseases. Report No. 17. Geneva: WHO; 2005. [ Links ]
2. Brisse S, Barnabe C, Tibayrenc M. Identification of six Trypanosoma cruzi phylogenetic lineages by random amplified polymorphic DNA and multilocus enzyme electrophoresis. Int J Parasitol. 2000;30:35-44. [ Links ]
3. Saravia NG, Holguin AF, Cibulskis RE, D'Alessandro A. Divergent isoenzyme profiles of sylvatic and domiciliary Trypanosoma cruzi in the eastern plains, piedmont, and highlands of Colombia. Am J Trop Med Hyg. 1987;36:59-69. [ Links ]
4. Ruiz-García M, Montilla M, Nicholls SO, Angarita L, Álvarez D. Genetic relationships and spatial genetic structure among clonal stocks of Trypanosoma cruzi in Colombia. Heredity. 2000;85:318-27. [ Links ]
5. Cuervo P, Cupolillo E, Segura I, Saravia N, Fernandes O. Genetic diversity of Colombian sylvatic Trypanosoma cruzi isolates revealed by the ribosomal DNA. Mem Inst Oswaldo Cruz. 2002;97:877-80. [ Links ]
6. Montilla MM, Guhl F, Jaramillo C, Nicholls S, Barnabe C, Bosseno MF, et al. Isoenzyme clustering of Trypanosomatidae Colombian populations. Am J Trop Med Hyg. 2002;66:394-400. [ Links ]
7. Salazar A, Schijman AG, Triana-Chávez O. High variability of Colombian Trypanosoma cruzi lineage I stocks as revealed by low-stringency single primer-PCR minicircle signatures. Acta Trop. 2006;100:110-8. [ Links ]
8. Moncayo A. Chagas disease: current epidemiological trends after the interruption of vectorial and transfusional transmission in the Southern Cone countries. Mem Inst Oswaldo Cruz. 2003;98:577-91. [ Links ]
9. Gutiérrez R, Angulo VM, Tarazona Z, Britto C, Fernandes O. Comparison of four serological tests for the diagnosis of Chagas disease in a Colombian endemic area. Parasitology. 2004;129:439-44. [ Links ]
10. Sosa S, Segura E. Tratamiento de la infección por Trypanosoma cruzi en fase indeterminada. Experiencia y normatización actual en la Argentina. Medicina. 1999;59(Suppl.2):166-70. [ Links ]
11. Lauria-Pires L, Braga MS, Vexenat AC, Nitz N, Simoes-Barbosa A, Tinoco DL, et al. Progressive chronic Chagas heart disease ten years after treatment with anti-Trypanosoma cruzi nitroderivatives. Am J Trop Med Hyg. 2000;63:111-8. [ Links ]
12. Urbina JA, Docampo R. Specific chemotherapy of Chagas disease: controversies and advances. Trends Parasitol. 2003;19:495-501. [ Links ]
13. Jannin J, Villa L. An overview of Chagas disease treat-ment. Mem Inst Oswaldo Cruz. 2007;102(Suppl.1):95-7. [ Links ]
14. Bhattacharya SK, Sinha PK, Sundar S, Thakur CP, Jha TK, Pandey K, et al. Phase 4 trial of miltefosine for the treatment of Indian visceral leishmaniasis. J Infect Dis. 2007;196:591-8. [ Links ]
15. Soto J, Soto P. Miltefosina oral para el tratamiento de la leishmaniasis. Biomédica. 2006;26(Suppl.1):207-17. [ Links ]
16. Croft SL, Seifert K, Duchene M. Antiprotozoal activities of phospholipid analogues. Mol Biochem Parasitol. 2003;126:165-72. [ Links ]
17. Saraiva VB, Gibaldi D, Previato JO, Mendonca-Previato L, Bozza MT, Freire-De-Lima CG, et al. Proinflammatory and cytotoxic effects of hexadecylphosphocholine (miltefosine) against drug-resistant strains of Trypanosoma cruzi. Antimicrob Agents Chemother. 2002;46:3472-7. [ Links ]
18. Lira R, Contreras LM, Rita RM, Urbina JA. Mechanism of action of anti-proliferative lysophospholipid analogues against the protozoan parasite Trypanosoma cruzi: potentiation of in vitro activity by the sterol biosynthesis inhibitor ketoconazole. J Antimicrob Chemother. 2001;47:537-46. [ Links ]
19. Santa-Rita RM, Santos H, Meirelles MN, de Castro SL. Effect of the alkyl-lysophospholipids on the proliferation and differentiation of Trypanosoma cruzi. Acta Trop. 2000;75:219-28. [ Links ]
20. Santa-Rita RM, Lira R, Barbosa HS, Urbina JA, de Castro SL. Anti-proliferative synergy of lysophospholipid analogues and ketoconazole against Trypanosoma cruzi (Kinetoplastida: Trypanosomatidae): cellular and ultrastructural analysis. J Antimicrob Chemother. 2005;55:780-4. [ Links ]
21. Andrade SG, Andrade V, Brodskyn C, Magalhaes JB, Netto MB. Immunological response of Swiss mice to infection with three different strains of Trypanosoma cruzi. Ann Trop Med Parasitol. 1985;79:397-407. [ Links ]
22. Filardi LS, Brener Z. Susceptibility and natural resistance of Trypanosoma cruzi strains to drugs used clinically in Chagas disease. Trans R Soc Trop Med Hyg. 1987;81:755-9. [ Links ]
23. Revollo S, Oury B, Laurent JP, Barnabe C, Quesney V, Carriere V, et al. Trypanosoma cruzi: impact of clonal evolution of the parasite on its biological and medical properties. Exp Parasitol. 1998;89:30-9. [ Links ]
24. Croft SL, Snowdon D, Yardley V. The activities of four anticancer alkyllysophospholipids against Leishmania donovani, Trypanosoma cruzi and Trypanosoma brucei. J Antimicrob Chemother. 1996;38:1041-7. [ Links ]
25. Klenner T, Beckers T, Nooter K, Holtmann H. Influence of hexadecylphosphocholine (miltefosine) on cytokine synthesis and biological responses. Adv Exp Med Biol. 1996;416:181-7. [ Links ]
26. Escobar P, Matu S, Marques C, Croft SL. Sensitivities of Leishmania species to hexadecylphosphocholine (miltefosine), ET-18-OCH(3) (edelfosine) and ampho-tericin B. Acta Trop. 2002;81:151-7. [ Links ]
27. Soto J, Arana BA, Toledo J, Rizzo N, Vega JC, Díaz A, et al. Miltefosine for new world cutaneous leishmaniasis. Clin Infect Dis. 2004;38:1266-72. [ Links ]
28. Escobar P, Yardley V, Croft SL. Activities of hexadecylphosphocholine (miltefosine), AmBisome, and sodium stibogluconate (Pentostam) against Leishmania donovani in immunodeficient scid mice. Antimicrob Agents Chemother. 2001;45:1872-5. [ Links ]
29. Murray HW, Delph-Etienne S. Visceral leishmanicidal activity of hexadecylphosphocholine (miltefosine) in mice deficient in T cells and activated macrophage microbicidal mechanisms. J Infect Dis. 2000;181:795-9. [ Links ]
30. Maya JD, Repetto Y, Agosin M, Ojeda JM, Téllez R, Gaule C, et al. Effects of nifurtimox and benznidazole upon glutathione and trypanothione content in epimastigote, trypomastigote and amastigote forms of Trypanosoma cruzi. Mol Biochem Parasitol. 1997;86:101-6. [ Links ]
31. de Castro SL. The challenge of Chagas' disease chemotherapy: an update of drugs assayed against Trypanosoma cruzi. Acta Trop. 1993;53:83-98. [ Links ]
32. Andrade SG, Rassi A, Magalhaes JB, Ferriolli Filho F, Luquetti AO. Specific chemotherapy of Chagas disease: a comparison between the response in patients and experimental animals inoculated with the same strains. Trans R Soc Trop Med Hyg. 1992;86:624-6. [ Links ]
33. Toledo MJ, Guilherme AL, da Silva JC, de Gasperi MV, Mendes AP, Gomes ML, et al. Trypanosoma cruzi: chemotherapy with benznidazole in mice inoculated with strains from Parana state and from different endemic areas of Brazil. Rev Inst Med Trop Sao Paulo. 1997;39:283-90. [ Links ]
34. Murta SM, Gazzinelli RT, Brener Z, Romanha AJ. Molecular characterization of susceptible and naturally resistant strains of Trypanosoma cruzi to benznidazole and nifurtimox. Mol Biochem Parasitol. 1998;93:203-14. [ Links ]
35. Laurent JP, Barnabe C, Quesney V, Noel S, Tibayrenc M. Impact of clonal evolution on the biological diversity of Trypanosoma cruzi. Parasitology. 1997;114:213-8. [ Links ]
36. de Lana M, da Silveira A, Barnabe C, Quesney V, Noel S, Tibayrenc M. Trypanosoma cruzi: compared vectorial transmissibility of three major clonal genotypes by Triatoma infestans. Exp Parasitol. 1998;90:20-5. [ Links ]
37. Toledo MJ, de Lana M, Carneiro CM, Bahia MT, Machado-Coelho GL, Veloso VM, et al. Impact of Trypanosoma cruzi clonal evolution on its biological properties in mice. Exp Parasitol. 2002;100:161-72. [ Links ]
38. Toledo MJ, Bahia MT, Carneiro CM, Martins-Filho OA, Tibayrenc M, Barnabe C, et al. Chemotherapy with benznidazole and itraconazole for mice infected with different Trypanosoma cruzi clonal genotypes. Antimicrob Agents Chemother. 2003;47:223-30. [ Links ]
39. Villarreal D, Barnabe C, Sereno D, Tibayrenc M. Lack of correlation between in vitro susceptibility to benznidazole and phylogenetic diversity of Trypanosoma cruzi, the agent of Chagas disease. Exp Parasitol. 2004;108:24-31. [ Links ]

