










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiomédica
Print version ISSN 0120-4157On-line version ISSN 2590-7379
Biomédica vol.30 no.2 Bogotá Apr./June 2010
ARTÍCULO ORIGINAL
1 Grupo de Investigación en Agentes Infecciosos y Cáncer, Instituto Nacional de Cancerología
2 Grupo de Investigación en Biología del Cáncer, Instituto Nacional de Cancerología, Bogotá D.C. , Colombia.
Recibido: 30/07/09; aceptado:26/02/10
Introduction. Studies using Western Helicobacter pylori strains have shown that a risk factor for gastric cancer is the number of EPIYA-C motifs in the cytotoxin-associated A protein. CagA is delivered into epithelial cells, where it becomes tyrosine phosphorylated in their EPIYA repeats and induces cytoskeleton rearrangements.
Objectives. The objective of this study was to evaluate H. pylori cagA positive strains isolated from Colombian patients with gastroduodenal diseases for the number of EPIYA-C repeats in cagA and their ability to induce cytoskeleton rearrangements in epithelial cells.
Materials and methods. We analyzed the 3´ EPIYA repeats region of cagA by PCR in 93 H. pylori cagA positive strains from 49 patients with gastritis, 17 with gastric cancer, and 24 with duodenal ulcer. AGS cells exposed to the various H. pylori isolates were evaluated for rearrangements in their cytoskeleton.
Results. Strains with one EPIYA-C were the most frequent in gastritis and duodenal ulcer patients. Strains with three EPIYA-C were mainly found in gastric cancer. We found a significantly higher risk of gastric cancer for individuals infected with strains harboring three EPIYA-C motifs (OR=12.4, CI95%: 2.32-66.3). Strains from gastric cancer showed significantly higher percentages of induction of cytoskeleton rearrangements in comparison with those from gastritis (p Mann-Whitney<0.005).
Conclusions. H. pylori strains with three EPIYA-C repeats can confer an increased risk of cancer to infected individuals.
Key words: Helicobacter pylori; polymorphism, genetic; gastritis, stomach neoplasm, duodenal ulcer.
Variación en el número de repeticiones EPIYA-C en la proteína CagA de aislamientos colombianos de Helicobacter pylori y su capacidad para inducir fenotipo colibrí en células epiteliales gástricas
Introducción. En los aislamientos de Helicobacter pylori del hemisferio occidental, se ha observado que el número de repeticiones EPIYA-C en la proteína CagA es un factor de riesgo para cáncer gástrico. La proteína CagA es introducida en la célula epitelial y, posteriormente, es fosforilada en las tirosinas presentes en los motivos EPIYA e induce rearreglos en el citoesqueleto.
Objetivos. Nuestro propósito fue evaluar el número de repeticiones EPIYA-C y la habilidad para inducir rearreglos en el citoesqueleto en los aislamientos de H. pylori positivos para cagA, provenientes de pacientes colombianos con enfermedad gastroduodenal.
Materiales y métodos. Mediante PCR, se analizó la región 3´ que contiene las repeticiones EPIYA-C, en 93 aislamientos de H. pylori positivos para cagA provenientes de 49 pacientes con gastritis, 17 con cáncer gástrico y 24 con úlcera duodenal. Los rearreglos del citoesqueleto se evaluaron mediante cultivos simultáneos de células AGS con las cepas de H. pylori.
Resultados. En gastritis y úlcera duodenal se observó la mayor frecuencia de aislamientos con EPIYA C; los aislamientos con tres repeticiones EPIYA-C se encontraron con mayor frecuencia en cáncer gástrico. Encontramos un riesgo de cáncer gástrico significativamente mayor para individuos infectados con cepas con tres repeticiones EPIYA-C (OR=12,4; IC95% 2,32-66,3). Los aislamientos provenientes de cáncer gástrico mostraron mayores porcentajes de inducción de rearreglos en el citoesqueleto que los observados con aislamientos provenientes de gastritis (prueba de Mann-Whitney menor de 0,005).
Conclusiones. La infección con cepas de H. pylori con tres repeticiones EPIYA-C puede conferir un mayor riesgo de desarrollar cáncer gástrico.
Palabras clave: Helicobacter pylori, proteína CagA, polimorfismo, gastritis, neoplasias del estómago, úlcera duodenal.
Helicobacter pylori is a human gastric pathogen which chronically infects more than half the world´s population (1). It colonizes the human stomach and persists for decades, causing chronic gastritis. In a small fraction of individuals, the infection progresses to peptic ulcer disease, gastric adenocarcinoma, or mucosa-associated lymphoid tissue (MALT) lymphoma (2-7). The causes for the different outcomes of H. pylori infections may include bacterial virulence, host susceptibility, and environmental factors (8-11). Individuals infected with strains harbouring the cag pathogenicity island (cag-PAI) are at a higher risk of developing duodenal ulcer, atrophic gastritis, and gastric cancer (8,12-15).
The cag-pathogenicity island (PAI) is a 40 kb DNA segment which contains genes encoding for a type IV secretion system (16,17), which forms a syringe-like structure that penetrates epithelial cells and allows the translocation of CagA, the product of cagA gene (16,18). Once introduced into the cell, CagA is phosphorylated by src family kinases on specific EPIYA sequences repeats (18-20) and then binds and activates phosphatase SHP-2 to induce an elongated cell shape known as hummingbird phenotype (21).
CagA proteins show size variation due to the presence of different types and/or numbers of repeat sequences containing the EPIYA motifs within the C-terminal variable region (7). Four types of EPIYA segments have been described: A, B, C, and D (22). East Asian H. pylori strains commonly have an ABD pattern, while Western H. pylori strains almost always contain EPIYA-A and EPIYA-B segments, followed by one to three EPIYA-C segments. It has been shown that CagA with more EPIYA-C motifs induced more cytoskeleton rearrangements (hummingbird phenotype) (22) and Western H. pylori strains with more than one EPIYA-C motifs have been significantly associated with gastric cancer (6,23-25).
In this study we analyzed the number of CagA EPIYA-C segments and the ability of the CagA protein expressed by H. pylori strains isolated from Colombian patients with gastroduodenal diseases to induce a hummingbird phenotype.
Materials and methods
Bacterial strains
A total of 93 H. pylori cagA positive strains originally isolated from patients living in Bogotá, Colombia between 1998 and 2002 (12,15) were obtained from the H. pylori stock collection of the Instituto Nacional de Cancerología, in Bogotá, Colombia. These strains were isolated from antral gastric biopsy specimens from 52 patients with gastritis (29 male and 23 female; mean age 52 years; age range 31-78 years), 17 patients with gastric cancer (12 male, 5 female; mean age 56, age range 42-67 years), and 24 patients with duodenal ulcer (11 male, 13 female; mean age 37, range 19-57). Two reference strains were included: NCTC 11638 and NCTC 11637. These strains were grown on blood agar plates, supplemented with 7 % horse serum (Invitrogen, Grand Island, NY), 1% Vitox (Oxoid, Basingstoke, UK), and Campylobacter selective supplement (Oxoid, Basingstoke, UK), at 37 °C for 3 days under microaerophilic conditions. Chromosomal DNA was prepared by DNazol (Invitrogen, Carlsbad, CA) extraction method, according to the manufacturer´s instructions. Ethics committee of the Instituto Nacional de Cancerología approved the protocol for this study. All the strains were isolated from patients that provided informed consent.
Amplification of the 3´-region of cagA by PCR
The primers CAGTF 5´-ACCCTAGTCGGTAATGGG-3´ and CAGTR 5´- GCTTTAGCTTCTGAYACYGC, described by Yamaoka et al. (24), were used to amplify the entire 3´ repeat region of cagA. PCR was performed in a volume of 25 µL containing 10 mM Tris, 50 mM KCl, 2.5 mM MgCl2, 200 µM dNTPs, 25 pmol of the primers, 100 ng H. pylori genomic DNA, and 1U Taq polymerase (Promega, Madison, WI). The PCR conditions included an initial denaturation step at 94°C for 5 min, followed up by 35 cycles consisting of 1 min at 94°C, 1 min at 52°C, and 1 min at 72°C, and then an extension step at 72°C for 7 min. PCR products were analyzed by 1.5% agarose gel electrophoresis with ethidium bromide staining.
Sequence analysis
To sequence the 3´ repeat region of cagA, PCR amplified products were excised from a 1.5% agarose gel and purified by using a GFX PCR DNA purification kit (Amersham, Buckinghamshire, UK), and cloned into pCR 4 TOPO (Invitrogen, Paisley, UK). DNA sequencing was performed with a 7-Deaza-dGTP Cy5/5.5 Dye Primer Cycle DNA sequencing kit (Visible Genetics, Toronto, Canada) on a personal sequence system SEQ4x4 (Visible Genetics, Etobicoke, Canada), by using M13R universal primer labelled with the Cy5.5 fluorescent dye. M13 fluorescent primers targeting the margins of the cloning sites were used for DNA sequencing of both strands. Nucleotide sequences were aligned and analyzed with BLAST tools. The previous published cagA sequences of strains NCTC 11638 (gene bank access AF282853) and NCTC 11637 (gene bank access AF202973) were also included in the analysis. The deduced peptide sequences containing the EPIYA motifs were aligned by CLUSTALW (http://www.ebi.ac.uk/Tools/clustalw2).
AGS cell hummingbird phenotype formation
AGS cell hummingbird phenotype formation was evaluated according to Argent et al. with minor modifications (6). Briefly, AGS cells were grown in six-well plates at a density of 2x105 cells/well in RPMI 1640 (Sigma Chemical Co. , St. Louis, MO), supplemented with 10% heat-inactivated fetal bovine serum (Invitrogen/GIBCO BRL, Grand Island, NY) and incubated for 24 hours at 37°C in a 5% CO2-air humidified atmosphere to obtain monolayers with a 70% to 80% confluence. H. pylori strains were harvested from 48 to 72 hour growth plates, suspended in RPMI 1640 medium, adjusted to an optical density at 600 nm of 0.1 (1 OD600nm= 2.4 x 108 CFU/mL) and added to the AGS cells at a multiplicity of infection (MOI) of 100. H. pylori strains were cocultured in triplicate with AGS cells for 24 hours at 37 °C in a 5% CO2-air humidified atmosphere, and then the cells were examined for hummingbird phenotype formation by microscopy (Axiovert 135 microscope, Zeiss, Germany) reading three randomly chosen fields in each of three wells. The results were reported as mean percentages of cells exhibiting the cellular elongation effect in the three wells. A hummingbird cell was defined as a cell with a needle-like protrusion >2 µm (6).
Statistical analysis
The statistical analysis of data was made by using univariate logistical regression, X2 test, and non-parametric Mann-Whitney test in SPSS for Windows version 16.0 software. A p <0.05 value was considered statistically significant.
Results
Analysis of the number of cagA EPIYA-C repeats in H. pylori clinical isolates
The 3´-region of cagA was amplified from the 93 cagA carrying H. pylori isolates from Bogotá, Colombia. As expected, there was diversity in the fragment size among isolates, with PCR products ranging in size between 400 bp and 700 bp (Figure 1). We observed a single band product in 85 strains and a double band product in 8 isolates. Fragments of 500 bp, 600 bp, 700 bp, and 400 bp were amplified from 53, 36, 10 and 2 isolates, respectively.
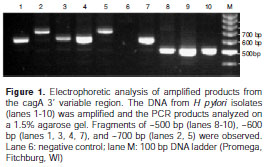
It has been shown that the size variation observed in the 3´-region of the cagA gene is due to variation in the number of repeat sequences containing the tyrosine phosphorylation motif EPIYA (6,26). In order to predict the number of EPIYA-C repeats in each strain, we performed nucleotide sequence analysis of six strains representing the different sizes in the 3´-region of the cagA gene and of two reference strains, 11637 and 11638. All the sequenced cagA variable regions were of the Western type. The deduced peptide sequences were aligned by CLUSTAL W (Figure 2). Isolates without EPIYA-C motifs (1071 and reference strain 11638), with one EPIYA-C motif (2033 and 3039), with two EPIYA-C motifs (1057), and with three EPIYA-C motifs (1034, 1039, and reference strain 11637) yielded 3´ cagA PCR products of 400 bp, 500 bp, 600 bp, and 700 bp, respectively. According to the sequencing data, we classified the cagA positive clinical isolates with regard to the predicted EPIYA-C number. The majority of our strains were found to harbor one EPIYA-C motif (49%). 31% of our strains contained two EPIYA-C motifs, and 10% of strains were found to have three EPIYA-C motifs. Only two strains were found to harbor no EPIYA-C motifs. With regard to the double bands, seven isolates were noted to harbor one and two EPIYA-C motifs and one isolate harbored one and three EPIYA-C motifs.

Association of the predicted number of EPIYA-C motifs with gastric cancer
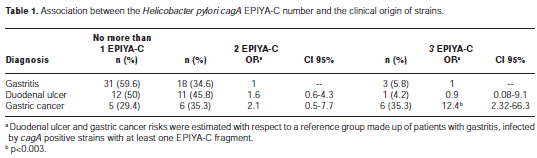
We analyzed the association between the number of EPIYA-C motifs at the 3´-end of cagA and the clinical origin of the strains (Table 1). Clinical isolates involving more than one strain were classified according to the highest observed number of EPIYA-C motifs. Strains with one EPIYA-C motif were the most frequent in gastritis and duodenal ulcer and were the least frequent in cancer patients. Strains with two EPIYA-C motifs showed frequencies between 34.6% and 45.8%, with gastritis showing the smallest frequency, and duodenal ulcer had the the highest. In gastric carcinoma patients, isolates harboring three EPIYA-C motifs represented 35.3% of cases, a higher percentage than the proportions observed in gastritis (5.8%) and duodenal ulcer (4.2%). We observed a significant higher risk of gastric cancer for individuals infected with strains harboring three EPIYA-C motifs in comparison with individuals infected with strains harboring one EPIYA-C motif (OR = 12.4, CI 95%: 2.32-66.3).
Higher induction of cytoskeleton rearrangements in AGS cells cocultured with H. pylori isolates from gastric cancer
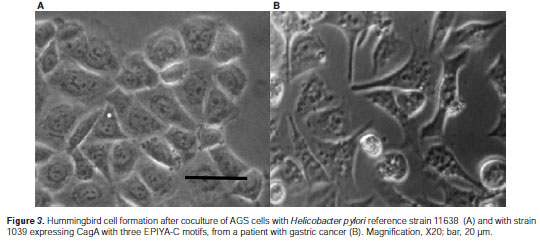
In order to assess, for clinical strains, whether the number of EPIYA-C motifs in the CagA protein affected the extent of induction of hummingbird phenotype, we cocultured AGS cells with 69 H. pylori strains (40 from gastritis, 15 from gastric cancer and 14 from duodenal ulcer). In spite of the high variability observed in the percentages of hummingbird cells in the cocultures, the mean percentages of hummingbird cells increased when the number of EPIYA-C motifs in the strains was high (one EPIYA-C: 38.2 %, two EPIYA-C: 41.3%, three EPIYA-C: 45.81%). These differences were not statistically significant (Figures 3 and figure 4A). When we analyzed the extent of humming bird cells induction in relation to the type of gastroduodenal disease, strains from gastric cancer showed significantly higher percentages of induction of cytoskeleton rearrangements, in comparison with gastritis (gastric cancer: 48.86% vs. gastritis: 36.4%, p< 0.005) (figure 4b).
<

Discussion
Infection with cagA positive H. pylori strains is correlated with a significantly higher risk of atrophic gastritis, gastric cancer, and duodenal ulcer (8,14). However, most persons colonized by these strains never develop severe gastroduodenal disease and remain asymptomatic. In Colombia, 72% to 90% of H. pylori strains possess the cagA gene (15,27), and the incidence of gastric cancer is high (28). It has been shown that the number of EPIYA-C segments on CagA influences the degree of virulence as well as the oncogenic potential of individual cagA positive H. pylori strains (25,29). Further, among Western strains, the most important factor for cancer risk is the number of CagA EPIYA-C motifs (25). In order to evaluate, in a high gastric cancer incidence population, the clinical relevance of the variations of the 3´-region of the cagA gene, we analyzed the number of EPIYA-C repeats and the ability to induce a hummingbird phenotype of H. pylori cagA positive strains isolated from Colombian patients with gastroduodenal diseases. Most of the strains possessed a cagA gene with one EPIYA-C motif in the 3´-region; its frequency was 49%, similar to the one reported by Yamaoka for Colombian isolates (55%) (30), but lower to the reported for isolates from South Africa (73%) (6), USA (80%) (30), France (68%) (30), and Greece (67.3%) (31). The frequency of strains harboring a cagA gene with two or three EPIYA-C motifs was 41%, similar to the frequency reported by Yamaoka for Colombian strains (44 %) (30) and higher than that reported for strains from USA (17 %) (30), France (32%)(30), South Africa (27%) (6), and Greece (21%) (31). Geographic variations in the frequency of H. pylori strains with one, two, or three EPIYA-C repeats may be explained by different acidic conditions in the gastric mucosa, which could be related to differences among populations in the frequencies of cytokine gene polymorphisms that attenuate gastric acid secretions (32). However, Argent et al. found no association between the number of EPIYA-C motifs and acid secretion (33).
We found 8.6% of isolates displaying a double band with sizes corresponding to different numbers of EPIYA-C repeats, all of them in benign diseases. This goes in accordance with the reports of Basso et al. (25) and Panayotopoulou et al. (31), who reported a similar frequency of isolates with this characteristic. Probably some of these strains reflect the adaptation of H. pylori to changes in the environment of the stomach by microevolution generating clones within an individual with different types of CagA proteins and also different degrees of virulence (33-35), and some could correspond to mixed infections.
As observed in previous studies (25,36), strains with cagA with higher number of EPIYA-C motifs were associated with a higher gastric cancer risk than strains with only one EPIYA-C motif. The risk of gastric cancer was 12 times higher in patients infected with cagA positive strains with three EPIYA-C motifs as compared with patients infected with cagA positive strains with one EPIYA-C motif. The high frequency of strains with more than one EPIYA-C motif could explain in part the high incidence of gastric cancer in Colombia.
Higashi et al (22) demonstrated, by generating EPIYA mutants from the strain NTCC 11637, that CagA with more EPIYA-C motifs induced more cytoskeleton rearrangements (hummingbird pheno-type). Argent et al. also reported an association between the number of EPIYA-C motifs and the extent of AGS cell elongation in strains from South Africa (6). In this study, we did not observe this association; instead, we found a high variability in the extent of hummingbird phenotype induction for strains with one and two EPIYA-C (3% to 60%), and for strains with three EPIYA-C (35% to 61%). Although the mean percentages of cytoskeleton rearrangement induction in AGS cells tended to increase in the direction one EPIYA-C ® two EPIYA-C ® three EPIYA-C, the differences were not significant. These results suggest that the induction of the hummingbird phenotype must be influenced by other mechanisms besides translocation and phosphorylation of CagA on EPIYA-C motifs. Interestingly, strains from gastric cancer showed a significantly higher ability to induce elongation than strains from gastritis. Probably, the ability to induce elongation is associated with the virulence of cagA positive strains.
In conclusion, our study documents a high frequency of H. pylori cagA positive strains with more than one EPIYA-C repeats in a high gastric cancer incidence country like Colombia. We do not confirm a correlation between the number of EPIYA-C repeats and the extent of the induction of morphological changes in epithelial cells. We confirmed that, among CagA positive strains, those with three EPIYA-C motifs are associated with a higher gastric cancer risk. The number of EPIYA-C motifs could be used as a risk marker for individuals with nonatrophic gastritis infected with cagA positive strains; this marker could be useful in a country like Colombia, with a high prevalence of infections with cagA positive H. pylori strains.
We did not identify any conflict of interest.
This research was supported by a grant from the Instituto Nacional de Cancerología, Bogotá D.C. , Colombia.
Corresponding author: María Mercedes Bravo, Grupo de Investigación en Agentes Infecciosos y Cáncer, Instituto Nacional de Cancerología, calle 1 Nº 9-85, Bogotá, D.C., Colombia. Teléfono: (571) 334 1111, extensión 4205; fax: (571) 334 1360 mbravo@cancer.gov.co
1. Suerbaum S, Michetti P. Helicobacter pylori infection. N Engl J Med. 2002;347:1175-86. [ Links ]
2. Blaser MJ, Berg DE. Helicobacter pylori genetic diversity and risk of human disease. J Clin Invest. 2001;107:767-73. [ Links ]
3. Israel DA, Peek RM. Pathogenesis of Helicobacter pylori-induced gastric inflammation. Aliment Pharmacol Ther. 2001;15:1271-90. [ Links ]
4. Parsonnet J. Helicobacter pylori: the size of the problem. Gut. 1998;43 (Suppl. 1):S6-9. [ Links ]
5. Peek RM, Jr. , Blaser MJ. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat Rev Cancer. 2002;2:28-37. [ Links ]
6. Argent RH, Kidd M, Owen RJ, Thomas RJ, Limb MC, Atherton JC. Determinants and consequences of different levels of CagA phosphorylation for clinical isolates of Helicobacter pylori. Gastroenterology. 2004;127:514-23. [ Links ]
7. Tummuru MK, Cover TL, Blaser MJ. Cloning and expression of a high-molecular-mass major antigen of Helicobacter pylori: evidence of linkage to cytotoxin production. Infect Immun. 1993;61:1799-809. [ Links ]
8. Blaser MJ, Perez-Perez GI, Kleanthous H, Cover TL, Peek RM, Chyou PH, et al. Infection with Helicobacter pylori strains possessing cagA is associated with an increased risk of developing adenocarcinoma of the stomach. Cancer Res. 1995;55:2111-5. [ Links ]
9. Camargo MC, Mera R, Correa P, Peek RM, Jr. , Fontham ET, Goodman KJ, et al. Interleukin-1beta and interleukin-1 receptor antagonist gene polymorphisms and gastric cancer: a meta-analysis. Cancer Epidemiol Biomarkers Prev. 2006;15:1674-87. [ Links ]
10. Gorouhi F, Islami F, Bahrami H, Kamangar F. Tumour-necrosis factor-A polymorphisms and gastric cancer risk: a meta-analysis. Br J Cancer. 2008;98:1443-51. [ Links ]
11. Bae JM, Lee EJ, Guyatt G. Citrus fruit intake and stomach cancer risk: a quantitative systematic review. Gastric Cancer. 2008;11:23-32. [ Links ]
12. Cittelly DM, Huertas MG, Martinez JD, Oliveros R, Posso H, Bravo MM, et al. Helicobacter pylori genotypes in non atrophic gastritis are different of the found in peptic ulcer, premalignant lesions and gastric cancer in Colombia. Rev Med Chil. 2002;130:143-51. [ Links ]
13. Kidd M, Lastovica AJ, Atherton JC, Louw JA. Conservation of the cag pathogenicity island is associated with vacA alleles and gastroduodenal disease in South African Helicobacter pylori isolates. Gut. 2001;49:11-7. [ Links ]
14. Nilsson C, Sillen A, Eriksson L, Strand ML, Enroth H, Normark S, et al. Correlation between cag pathogenicity island composition and Helicobacter pylori-associated gastroduodenal disease. Infect Immun. 2003;71:6573-81. [ Links ]
15. Quiroga AJ, Cittelly DM, Bravo MM. BabA2, oipA and cagE Helicobacter pylori genotypes in Colombian patients with gastroduodenal diseases. Biomédica. 2005;25:325-34. [ Links ]
16. Censini S, Lange C, Xiang Z, Crabtree JE, Ghiara P, Borodovsky M, et al. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc Natl Acad Sci USA. 1996;93:14648-53. [ Links ]
17. Akopyants NS, Clifton SW, Kersulyte D, Crabtree JE, Youree BE, Reece CA, et al. Analyses of the cag pathogenicity island of Helicobacter pylori. Mol Microbiol. 1998;28:37-53. [ Links ]
18. Odenbreit S, Puls J, Sedlmaier B, Gerland E, Fischer W, Haas R. Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science. 2000;287:1497-500. [ Links ]
19. Stein M, Rappuoli R, Covacci A. Tyrosine phosphorylation of the Helicobacter pylori CagA antigen after cag-driven host cell translocation. Proc Natl Acad Sci USA. 2000;97:1263-8. [ Links ]
20. Stein M, Bagnoli F, Halenbeck R, Rappuoli R, Fantl WJ, Covacci A. c-Src/Lyn kinases activate Helicobacter pylori CagA through tyrosine phosphorylation of the EPIYA motifs. Mol Microbiol. 2002;43:971-80. [ Links ]
21. Segal ED, Cha J, Lo J, Falkow S, Tompkins LS. Altered states: involvement of phosphorylated CagA in the induction of host cellular growth changes by Helicobacter pylori. Proc Natl Acad Sci USA. 1999;96:14559-64. [ Links ]
22. Higashi H, Tsutsumi R, Fujita A, Yamazaki S, Asaka M, Azuma T, et al. Biological activity of the Helicobacter pylori virulence factor CagA is determined by variation in the tyrosine phosphorylation sites. Proc Natl Acad Sci USA. 2002;99:14428-33. [ Links ]
23. Azuma T, Yamakawa A, Yamazaki S, Fukuta K, Ohtani M, Ito Y, et al. Correlation between variation of the 3´ region of the cagA gene in Helicobacter pylori and disease outcome in Japan. J Infect Dis. 2002;186:1621-30. [ Links ]
24. Yamaoka Y, el Zimaity HM, Gutierrez O, Figura N, Kim JG, Kodama T, et al. Relationship between the cagA 33´ repeat region of Helicobacter pylori, gastric histology, and susceptibility to low pH. Gastroenterology. 1999;117:342-9. [ Links ]
25. Basso D, Zambon CF, Letley DP, Stranges A, Marchet A, Rhead JL, et al. Clinical relevance of Helicobacter pylori cagA and vacA gene polymorphisms. Gastroenterology. 2008;135:91-9. [ Links ]
26. Reyes-Leon A, Atherton JC, Argent RH, Puente JL, Torres J. Heterogeneity in the activity of Mexican Helicobacter pylori strains on gastric epithelial cells and it´s association with diversity in the cagA gene. Infect Immun. 2007;75:3445-54. [ Links ]
27. Bravo LE, van Doom LJ, Realpe JL, Correa P. Virulence-associated genotypes of Helicobacter pylori: do they explain the African enigma? Am J Gastroenterol. 2002;97:2839-42. [ Links ]
28. Pineros M, Ferlay J, Murillo R. Cancer incidence estimates at the national and district levels in Colombia. Salud Publica Mex. 2006;48:455-65. [ Links ]
29. Naito M, Yamazaki T, Tsutsumi R, Higashi H, Onoe K, Yamazaki S, et al. Influence of EPIYA-repeat polymorphism on the phosphorylation-dependent biological activity of Helicobacter pylori CagA. Gastroenterology. 2006;130:1181-90. [ Links ]
30. Yamaoka Y, Osato MS, Sepulveda AR, Gutierrez O, Figura N, Kim JG, et al. Molecular epidemiology of Helicobacter pylori: separation of H. pylori from East Asian and non-Asian countries. Epidemiol Infect. 2000;124:91-6. [ Links ]
31. Panayotopoulou EG, Sgouras DN, Papadakos K, Kalliaropoulos A, Papatheodoridis G, Mentis AF, et al. Strategy to characterize the number and type of repeating EPIYA phosphorylation motifs in the carboxyl terminus of CagA protein in Helicobacter pylori clinical isolates. J Clin Microbiol. 2007;45:488-95. [ Links ]
32. Snaith A, El-Omar EM. Helicobacter pylori: host genetics and disease outcomes. Expert Rev Gastroenterol Hepatol. 2008;2:577-85. [ Links ]
33. Argent RH, Thomas RJ, Aviles-Jimenez F, Letley DP, Limb MC, El-Omar EM, et al. Toxigenic Helicobacter pylori infection precedes gastric hypochlorhydria in cancer relatives, and H. pylori virulence evolves in these families. Clin Cancer Res. 2008;14:2227-35. [ Links ]
34. Aras RA, Lee Y, Kim SK, Israel D, Peek RM Jr, Blaser MJ. Natural variation in populations of persistently colonizing bacteria affect human host cell phenotype. J Infect Dis. 2003;188:486-96. [ Links ]
35. Covacci A, Falkow S, Berg DE, Rappuoli R. Did the inheritance of a pathogenicity island modify the virulence of Helicobacter pylori? Trends Microbiol. 1997;5:205-8. [ Links ]
36. Yamaoka Y, Kodama T, Kashima K, Graham DY, Sepulveda AR. Variants of the 3´ region of the cagA gene in Helicobacter pylori isolates from patients with different H. pylori-associated diseases. J Clin Microbiol. 1998;36:2258-63. [ Links ]

