










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiomédica
Print version ISSN 0120-4157On-line version ISSN 2590-7379
Biomédica vol.31 no.2 Bogotá Apr./June 2011
REVISIÓN DE TEMA
Grupo Inmunovirología, Facultad de Medicina, Universidad de Antioquia, Medellín, Colombia
The natural history of human immunodeficiency virus type-1 (HIV-1) infection is a complex and variable process that, similarly to other infections, has clearly demonstrated the existence of mechanisms of human natural resistance. The resistance either inhibits the establishment of infection or delays disease progression. When there is continuous exposure to infectious viral particles, several genetic and immunological mechanisms are essential to lead to resistance to HIV-1 infection/progression. The objective of this manuscript was to review the different mechanisms so far proposed to be responsible for HIV-1 resistance and to present the main results derived from 10 years of research in this area among Colombian subjects. In particular, this review focuses on determining the mechanisms involved in the protection of a group of individuals repeatedly exposed to the virus but who remained exempt of serological and clinical evidence of HIV-1 infection. Although the studies carried out in our research group corroborated the protective role of some of the previously proposed mechanisms of protection, ongoing research worldwide has made it clear that the phenomenon of human natural resistance depends on multiple factors with an important genetic influence, and only multicenter studies involving individuals with different genetic backgrounds may determine more universal mechanisms of resistance. Increasing our knowledge in this field will contribute to the development of novel preventive and therapeutic measures.
Key words: HIV, acquired immunodeficiency syndrome, infection control, HLA antigens; receptors, CCR5; killer cells, natural; apoptosis, defensins.
Mecanismos de resistencia natural al VIH en seres humanos: un resumen de 10 años de investigación en población colombiana
La historia natural de la infección por el virus de la inmunodeficiencia humana de tipo 1 (VIH-1) es un proceso variable y complejo que, en forma similar a otras infecciones, ha hecho evidente la existencia de mecanismos de resistencia natural que pueden inhibir el establecimiento de la infección o su progresión hacia estadios avanzados. Cuando hay una exposición continua a partículas virales infecciosas, varios mecanismos genéticos e inmunitarios son esenciales para que se establezca la resistencia.
El objetivo de este manuscrito fue revisar todos los mecanismos de resistencia al VIH-1, hasta el momento propuestos en seres humanos, y presentar los resultados más importantes que se han obtenido en diferentes estudios realizados en los últimos 10 años de investigación en esta área, en individuos colombianos, particularmente enfocados en determinar los mecanismos involucrados en la protección de un grupo de personas que se han expuesto repetidamente al virus, pero que han permanecido sin evidencia clínica ni serológica de infección por el VIH-1.
Aunque los estudios llevados a cabo por nuestro grupo de investigación han corroborado el papel protector de algunos de los mecanismos de protección previamente propuestos, la investigación actual en esta área, a nivel mundial, ha hecho evidente que el fenómeno de resistencia natural depende de múltiples factores con una gran influencia genética, y que sólo mediante estudios multicéntricos que involucren individuos con diferente componente genético, podrán establecerse los mecanismos universales de protección. Profundizar en el conocimiento en esta área permitirá el desarrollo de nuevas medidas preventivas y terapéuticas para la infección por el VIH-1.
Palabras clave: VIH, síndrome de inmunodeficiencia adquirida, control de infecciones, antígenos HLA, receptores CCR5, células asesinas naturales, apoptosis, defensinas.
Human immunodeficiency virus type-1 (HIV-1) infection continues to be an expanding epidemic worldwide. The risk of HIV-1 transmission varies and depends on multiple factors, in particular on the type of exposure: blood transfusions represent the greatest risk, with 90%, followed by vertical transmission with a 10-30% risk in the absence of any prophylactic intervention. In contrast, the risk of transmission by sharing infectious needles is low, 1% similar to the 0.3% risk during accidental exposure in health care personnel (1,2). The efficiency of sexual transmission is very low; the incidence of transmission has been estimated to be 0.0001-0.004 per sexual intercourse depending in the type of relation: the highest probability of infection occurs during male to male intercourses (1/10-1/1600), followed by male to women (1/200-1/2000) and women to male (1/200-1/10000) sexual intercourses (3). Oral sex might also lead to HIV-1 transmission with a very low frequency (3). Although sexual transmission is the least efficient, more than 80% of the infections worldwide have been acquired through sexual intercourse (4).
During sexual transmission, the virus can pass through the mucosa, come into contact mainly with macrophages and dendritic cells (DC) and infect them (through the CD4, CCR5 or CXCR4 molecules) (5-8), or else bind to C-type lectins (Langerin, DC-SIGN, DEC-205, mannose receptor and dendritic cell immunoreceptor-DCIR) expressed on their surface, both during infectious and noninfectious entry of HIV into these cells (9-11). It was recently also demonstrated that CD4+ T cells localized in the genital mucosa can be primary targets of HIV-1 (12). This occurs when the integrity of the epithelium is lost, a frequent phenomenon during sexual intercourse. Thereafter, the subsequent steps in HIV-1 infection are complex and variable, and the infected individuals exhibit different patterns of progression (13,14).
Moreover, despite the fact that they are repeatedly exposed to HIV-1, certain individuals exhibit no serological or clinical evidence of HIV-1 infection; they are known as exposed seronegative individuals (ESN) (15). This group of persons includes individuals who perform repeated unprotected sexual intercourses with HIV-1-seropositive individuals, neonates born to HIV-1 infected mothers, intravenous drug users, receptors of blood products from HIV-1-infected donors and health care workers with accidental exposures to HIV-1 (16,17). These individuals have clearly shown the existence of mechanisms of natural protection from acquired HIV-1 infection.
The mechanisms of human natural resistance to infectious diseases are essentially genetic or immunological. The main genetic mechanism so far reported to be responsible for resistance to HIV-1 infection is the Δ;32 mutation in the ccr5 gene (18,19). However, the Δ32/Δ32 genotype is only present in 2 to 4% of Caucasians individuals (7,20,21). Other variants in the chemokine genes have been more closely related to slow clinical progression than to resistance (22,23). The degree of concordance of the major histocompatibility complex between mother and neonate has been proposed to be a determining factor in the incidence of vertical transmission (24), similar to the presence of certain class I and class II MHC alleles (25).
Apoptosis or programmed cell death is an important mechanism of control during the ontogeny of the immune system that also plays a role during the immune response to several pathogens (26). Indeed, it seems to play a key role in natural resistance to Mycobacterium tuberculosis exhibited by some strains of mice, preventing the dissemination of M tuberculosis (27). During viral infections, apoptosis of target cells might also prevent viral dissemination (28,29).
The immune response against viruses is initiated by innate immune cells such as NK cells, granulocytes and γδ T cells that eliminate viruses through cytolytic and none-cytolytic mechanisms (30-32). DC and macrophages act as antigen presenting cells and produce several chemokines and cytokines which amplify the immune response (33). The humoral and cellular adaptive immune responses complement the antiviral action through the production of antibodies, cytokines and the cytotoxic activity of CD8+ T cells (CTLs) (34). More recently, several soluble factors, produced by different cells during the immune response have shown great antiviral activity as their primary function or as an accidental effect.
This article summarizes the different mechanisms so far associated with the phenomenon of human natural resistance to HIV-1, emphasizing the results of several studies carried out in different cohorts of sexually HIV-1 exposed-seronegative individuals in Medellin, Colombia during the last 10 years. It is important to stress that understanding the protective mechanisms involved in HIV-1 infection is essential for the development of new preventive and therapeutic measures.
Genetic mechanisms for HIV resistance
Polymorphisms and mutations in genes of the chemokine system
CCR5 is the port of entry for R5 viral strains, which are the primary transmission strains that predominate during the asymptomatic phase of HIV-1 infection (35). In 1996, a genetic factor, the deletion of 32 base pairs in the ccr5 gene known as the Δ32 mutation, was associated with a high degree of resistance to HIV-1 infection. This mutation generates a truncated protein that is not expressed on the cell membrane (18,19). Homozygous individuals for the Δ32 mutation (Δ32/Δ32) are highly resistant to infection by R5 HIV-1 viral strains while heterozygous persons (Δ32/ccr5) exhibit delayed progression to AIDS (18). Different reports indicate that the Δ32/Δ32 genotype is only present in 2-4% of Caucasian ESN individuals (7,20).
In addition, it is suggested that other polymorphisms in the ccr5 gene influence the risk of HIV-1 transmission and progression (36). The single nucleotide polymorphisms (SNP) so far reported in the regulatory region of ccr5 (A29G, G208T, G303A; T627C, C630T, A676G, C927T) is grouped in 7 different haplotypes (A-G); these haplotypes might dramatically affect disease progression depending on the genetic background of the infected individual (36).
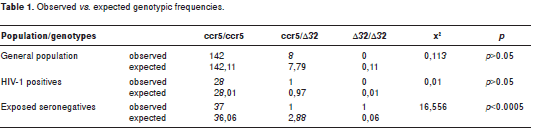
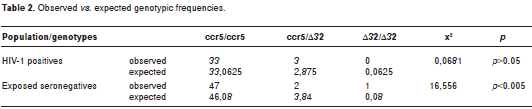
We evaluated in two studies the presence of mutations in the coding region of the ccr5 gene. These mutations could be associated with natural resistance of HIV-1 exposed-seronegative individuals from Medellin, Colombia. In the first study (37) we evaluated the frequency of the Δ32 mutation in the ccr5 gene by the polymerase chain reaction (PCR) in a population of 218 individuals: 29 HIV-1 positive (SP); 39 ESN and 150 individuals from the general population (GP). The frequency of the Δ32 mutated allele was 1.7% for SP, 3.8% for ESN and 2.7% for GP. The heterozygous genotype (Δ32/ccr5) was found in 3.4% of SP, 2.6% of ESN, and in 5.3% of GP. Only one ESN exhibited the homozygous genotype Δ32/Δ32. These differences were not statistically significant.
In the second study, we evaluated the complete codifying region of the ccr5 gene (21). Here, we screened 50 ESN and 36 SP individuals. A PCR was first performed for amplifying exon 4 to obtain an 1114 bp product; this product was digested with the Hin fI restriction enzyme yielding the followings fragments: 355, 215, 202, 159, 144 and 40 bp. These restriction fragments were analyzed by single strand conformational polymorphisms (SSCP). No significant differences in the allelic and genotypic frequencies were detected between ESN and SP. The frequency of the Δ32 mutated allele was 4.2% for SP and 4% for ESN. The heterozygous genotype (Δ32/ccr5) was observed in 8.3% of SP and 4% of ESN; only one ESN (2%) has the homozygous mutated genotype (Δ32/Δ32).
However, when we compared the observed vs. the expected genotypic frequencies in both studies, these frequencies were significantly higher in the ESN group (table 1 and table2), suggesting that the Δ32/Δ32 genotype could have a protective effect in the population of ESN individuals examined here.
Major histocompatibility complex (MHC)
Several studies in different cohorts of HIV-1-positive patients show the association between alleles expression of particular human leukocyte antigen (HLA) and resistance/susceptibility to HIV-1 infection; this association might be related to the influence that specific alleles have on the quality of the immune response that is established against certain infections. The heterozygosity of HLA class I alleles are associated with delayed progression of HIV-1 infection (38,39), similar to the association found between the presence of the B27 and B57 HLA alleles with delayed AIDS development (39-42). On the other hand, class I HLA allele discordance between mother and neonate is associated with protection against vertical transmission of HIV-1, an effect that is dose-dependent (43). Indeed, we postulated that during pregnancy alloantigen induce the production of soluble factors, including RNases that have anti-HIV-1 activity (44,45). Similarly, the presence of the super type HLA A2/6802 was associated with a lower risk of vertical and horizontal transmission of HIV-1 (45).
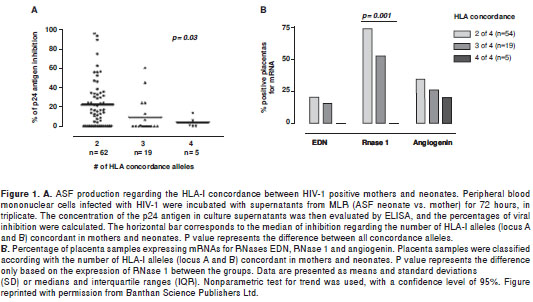
In this respect, we investigated whether the degree of mother to neonate discordance of class I HLA was associated with the ability to produce alloantigen-stimulated factor (ASF) with anti-HIV activity, and with the expression of RNases in placenta (46). The mother and neonate were HLA-typed and mixed lymphocyte reactions (MLRs) were carried out using cord-blood lymphocytes and the mother´s peripheral blood mononuclear cells. The antiviral activity of the supernatants from the MLRs was determined in an in vitro infection assay of PHA blasts, measured through the HIV-1 p24 antigen levels by ELISA. The expression of mRNAs for the RNase eosinophil-derived neurotoxin (EDN), RNase 1 and angiogenin was assessed by RT-PCR. An anti-HIV-1 activity was detected in the MLR supernatants (neonate versus mother), which was associated with the degree of class I HLA allele discordance (figure 1A). This class I HLA discordance was also associated with the expression of RNase 1 in placenta (figure 1B). These findings are consistent with the hypothesis that class I HLA allele discordance may induce the expression of this RNase in placenta and the production of soluble factors with anti-HIV-1 activity, contributing to the innate resistance to this pathogen in the context of vertical transmission.
Immunological mechanisms for HIV resistance
Apoptosis of target cells
Apoptosis is a regulated cell death program that can be used by different microorganisms as one of their pathogenic mechanisms, but at the same time can be used by the host to resist and control the establishment of different infections (47). Similarly, a variety of pathogens have developed surviving strategies blocking or inhibiting apoptosis (48). With respect to viral infections, both monocytes and macrophages use apoptosis to block early events of replication of virus Influenza A (49). In addition, BCS-1 cells infected by Theiler´s murine encephalomyelitis virus enter in apoptosis limiting viral dissemination (50).
During the early phases of HIV-1 infection, monocytes/macrophages are the main target cells that also act as viral reservoirs (51). Although the percentage of HIV-1-infected macrophages in tissues (brain, lymphoid node and lung) from patients can range from 1-50%, during natural infection only 0.001-1% of peripheral blood monocytes are infected (52,53). During an in vitro model of infection there is a low percentage of infected monocytes (54,55), indicating a lower susceptibility of monocytes to HIV infection when compared to macrophages (56,57).
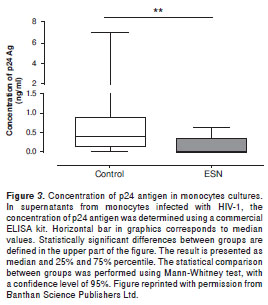
With this background we carried out a study to determine whether apoptosis is one of the mechanisms through which some of the ESN individuals prevent the establishment of HIV-1 infection (58). For this study we evaluated 15 SP, 20 ESN and 20 HIV-1 negative individuals. After monocytes and peripheral blood mononuclear cell isolation, Annexin V and DiOC6 assays allowed assessing spontaneous and HIV-1 induced apoptosis. Commercial ELISA determined the levels of p24 as indicator of viral replication. The results indicated that monocytes from ESN individuals suffer significant higher levels of spontaneous (figure 2A) and HIV-1 (figure 2B) induced apoptosis compared to the control group.
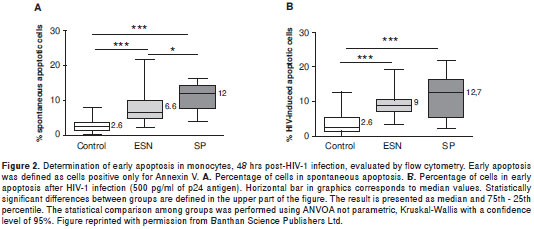
In addition, the levels of p24 were significantly lower in supernatants of HIV-1-infected monocytes from ESN, than in supernatants from the control group (figure 3). However, the correlation coefficient between percentage of cells in apoptosis and concentration of p24 antigen was not significant. These results suggest that after viral exposure, apoptosis of monocytes in some ESN individuals could play an important role in avoiding the establishment of HIV-1 infection, but other factors associated with natural resistance to this infection could co-participate in avoiding the process of invasion/infection in ESN subjects.
Innate immunity
Innate immunity is the first line of defense and is essential for the control of pathogens in skin and epithelial and mucous membranes. The innate immune response is activated through the engagement of toll-like receptors (TLRs) by pathogen-associated molecular patterns (PAMPs); this response is very heterogeneous and includes a cellular, intracellular and a soluble component (59). Macrophages, natural killer (NK) cells and NK T cells (NKT) very rapidly destroy infected cells, while plasmacytoid dendritic cells (pDC) produce high amounts of IFN-α, a soluble component with very potent antiviral activity. The myeloid dendritic cells (mDC), pDC and T cells with invariant TCR and restricted by the CD1d molecule, known as invariant NKT (iNKT) cells, establish the interaction between the innate and the adaptive branches during the anti-infectious immune responses (60).
Recent investigations established the very important role played by the innate immunity during the evolution of HIV-1 infection (61). Several cells from the innate response are target cells for this viral infection (62,63), and different alterations of this response in HIV-1 infected individuals have been described (62,64,65), suggesting that these abnormalities could increase the immunodeficiency caused by the massive loss of CD4+ T lymphocytes. However, the role of the innate response in HIV-1 resistance has not been established, despite the fact that all its components play an active role in mucous membranes the main port of viral entry, and in the induction of the adaptive immune responses.
Therefore, we investigated the association between quantitative and functional parameters of pDC, mDC, monocytes, NK and iNKT with human natural resistance to HIV-1 infection (66). In total, 38 SP, 30 ESNs and 35 HIV-1 negative controls were examined. The frequency and phenotype of the different innate immune cell subpopulations were determined by flow cytometry; the functional ability of these cells was also evaluated measuring the expression of CD86 and CD69, the production of the cytokines IL-6, IL-10 and TNF-α by flow cytometry and the concentration of IFN-α by ELISA in culture supernatants after stimulation of PBMC with TLR 9 agonists (class A CpG ODN 2216). In addition, the expression of intracellular IFN-γ was evaluated by flow cytometry after the stimulation of PBMC with PMA plus ionomycin.
The most striking finding in this study was the demonstration that NK and NKT cells from ESN individuals produced a significantly higher amount of IFN-γ in response to stimulation, compared to the control group (figure 4). Considering the strong antiviral activity of IFN-γ, this finding suggests that the production of this cytokine by innate immune cells could be associated with the phenomenon of natural resistance to HIV-1 exhibited by this group of ESN individuals.
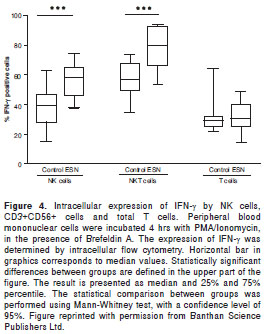
A similar frequency of all the evaluated innate immune cell subpopulations was found between ESN and the control group, while a higher frequency of DC, NK and iNKT cells was found in ESN compared to SP individuals. The CpG ODN stimulation induced similar expression of CD86 and CD69 on innate immune cells in all the groups under investigation. The secretion of IL-6, IL-10 and TNF-α cytokines was similar between ESN and controls, while the production of IFN-α was significantly decreased in the SP group. Although, we did not find differences between these parameters in this cohort of ESN, this does not rule out their relevance in human natural resistance to HIV-1.
Adaptive immunity
B cells, CD4+ and CD8+ T lymphocytes participate in the control of viral replication in HIV-1-infected individuals; in fact, certain reports indicate that these cell subpopulations might play an important role in preventing the establishment of HIV-1 infection (67).
Humoral immunity: since the majority of HIV-1 infections worldwide have been acquired through sexual intercourse, the virus enters more frequently through the genital mucosa. Therefore, mucosal immunity, particularly mediated by secretory IgA should play a role in preventing HIV-1 infection. In a cohort of sexually ESN individuals high levels of anti-HIV-1 IgA were detected in urine and vaginal washes, suggesting an association between humoral immunity and human natural resistance to HIV-1 (68). So far, this parameter has not been evaluated in our population of ESN.
Cellular immunity: a high production of Th1 cytokines in ESN individuals has been reported (67). In HIV-1 negative babies born to HIV-1 positive mothers, a significantly higher activity of anti-HIV-1 CD8+T lymphocytes was found, compared to babies who acquired HIV-1 infection (69,70). The presence of anti-HIV-1 specific CD8+ T cells has been also reported in sexually ESN and health care workers who have been accidentally exposed but remain seronegative to HIV-1 (71-73). In addition, HIV-1-gag specific T cells and an increased number of HIV-1 specific memory CD4+ and CD8+ T cells (CD45RA-/CCR7+) has been reported in ESN (74).
Immune soluble factors
Several soluble factors are postulated as essential for the anti-HIV-1 immune response. Their antiviral activity is executed through non-cytolytic mechanisms, underlining their therapeutic potential as a complement of the current antiretroviral therapy. These factors include β-chemokines (75), CAF (76), SDF-1 (23), IFN-α (77), LIF (78), lysozymes, RNases, ASF (79) and α y β defensins produced by epithelial and immune cells (80,81).
Defensins are microbicidal peptides of 30 to 48 cationic amino acids that are produced constitutively in response to different microorganisms or in response to proinflammatory cytokines (82), constituting one of the main mediators of innate responses in mucous membranes (8,40). α-defensins are found in neutrophils, paneth cells and epithelial cells in the urogenital female tract (83), (84). Recent investigations reported an increased production of α-defensins in CD8+ T lymphocytes and cervicovaginal mononuclear cells of ESN individuals compared to healthy controls, suggesting their role in human natural resistance to HIV-1 (85).
To date, 28 human β defensins (hBD) have been described. hBD 1 to 4 are the best characterized; hBD-1 is constitutively expressed while hBD-2 whose secretion is induced is present in the skin, saliva, plasma and, in higher amounts, in the urogenital tract. In addition, its mRNA has been located in lungs, the trachea, gut, uterus, liver and kidney. hBD-3 is induced in the heart, skeletal muscle, placenta, skin, esophagus, gingival keratinocytes, trachea and fetal thymus. hBD-4 is induced in testicles, the uterus, thyroid glands, lungs and kidneys (86,87). HIV-1 infection induces the expression of the mRNAs for hBD-2 and 3 in human oral epithelial cells, inhibiting replication of the virus; this inhibition involves direct binding to the viral particle and a negative regulation of the CXCR4 molecule, suggesting the hBD could play an important role in the protection of oral and other mucosal surfaces during exposure to HIV-1 (80). However, the expression of hBD in oral and genital mucosa of ESN individuals has not been explored.
On the other hand, it is postulated that some SNPs in the DEFB1 gene, which codes for hBD-1, could modulate the function of hBD during exposure to the pathogen and to explain their association with some biological events including resistance/susceptibility to infections. The SNP A692G could be associated with the transcription factor NF-κB and expression of molecules involved in host defense. The SNP A1836G is a probable polyadenylation site, and this polymorphism could alter transcription and/or translation of the gene. The SNP G1654A is adjacent to the first of six cysteine residues and due to its location it could be involved in the peptide function (88,89).
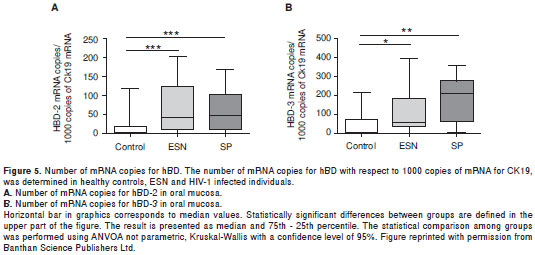
For this reason, we evaluated the association between the expression of hBD and the ESN condition in individuals who were sexually exposed to HIV-1 (90). Peripheral blood, oral, vaginal and endocervical mucosal samples were collected from 47 ESN, 44 SP and 39 healthy controls. In all ESNs the presence of the homozygous Δ32 mutation was ruled out. The number of mRNA copies for hBD-1, -2 and -3 was determined by real time RT-PCR. In addition, the polymorphisms A692G/G1654A/A1836G in the DEFB1 gene were determined by restriction fragment length polymorphisms and confirmed by sequencing. The ESN expressed significant higher numbers of copies of hBD-2 and -3 in the oral mucosa compared to healthy controls (figure 5A, figure 5B). The number of mRNA copies for hBD-1 and -2 in vaginal and endocervical mucosa was also higher in ESN compared to healthy controls, although the differences were not statistically significant. These results suggest that a higher expression of mRNA for hBD-2 and -3 could contribute to the natural resistance exhibited by this group of ESNs, and once again support the hypothesis that defensins, together with other soluble factors in this compartment, could be linked to a lower risk of HIV transmission via oral sexual practices.
Concluding remarks
Despite intensive research on HIV-1 infection, there is neither vaccine to prevent this disease nor treatment that allows complete eradication of this viral infection. Therefore, the identification of protective mechanisms to either infection or disease progression are required to develop new preventive or therapeutic protocols effective in controlling the pandemic caused by this virus.
In a setting in which there is sustained exposure to infectious virus, the key elements to resist HIV-1 infection, and ultimately slow-down disease progression, reside mainly on genetic and immune components of the host. Several gene products participate in recognizing the virus and responding to it. Both, the innate and the adaptive immune systems participate in a coordinate manner through soluble and cellular components in order to control viral replication. Considering that there are at least three different routes for HIV-1 transmission, different mechanisms are most likely associated with protection in each one of them, or at least the magnitude of their participation would be variable. Several studies carried out in different cohorts of ESNs have not led to the establishment of a unique mechanism responsible for protection from HIV-1 infection. On the contrary, these studies indicate that there is a great heterogeneity in the mechanisms so far associated with resistance, suggesting a multifactorial process that depends on the immunogenetic background of the host. However, it is important to point out that the criteria used to classify ESN individuals have varied between studies, and this might have influenced some of the results reported.
With the data so far at hand, it is clear that a preventive immunization protocol for HIV-1 should induce an immediate immune response that involves both humoral and cellular adaptive immune responses. Innate immunity and antiviral mechanisms active in mucosas, in particular soluble factors, are the ideal complement for an anti-HIV-1 response that might allow efficient viral control.
The central question raised is: could these factors, at least in combination, be enough to protect against HIV-1 infection? Unfortunately, this question is still unanswered and its response could only be established through collaborative studies including various cohorts of ESN individuals throughout the world; this would make it possible to determine mechanisms of human natural resistance beyond the influence of gene variants prevalent in particular racial backgrounds.
The magnitude of the current HIV-1 pandemic and the permanent difficulties encountered in developing therapeutic and preventive measures, underlines the fact that we are facing a pathogen that only through a natural evolutionary process, would humankind be able to face the challenges imposed by this infectious agent.
Acknowledgements
Authors thanks to Gene Shearer, Alan Landay, and Michael Lederman for their academic support, and Anne-Lise Haenni for the revision of the manuscript.
Conflict of interest
None of the authors has any potential financial conflict of interest related to this manuscript.
Financial support
The financial support for these investigations came from the following agencies: The Colombian Agency for the Development of Science and Technology - Colciencias; The Committee for the Development of Research from the University of Antioquia – CODI- Sostenibilidad 2008-2009; “Fundación para la Promoción de la Investigación y la Tecnología del Banco de la República”.
References
1. UNAIDS, WHO. Colombia: Epidemiological fact sheets on HIV/AIDS and sexually transmitted infections. Geneva: UNAIDS; 2004. [ Links ]
2. Summers T. Public policy for health care workers infected with the human immunodeficiency virus. JAMA. 2001;285:882. [ Links ]
3. Shattock RJ, Moore JP. Inhibiting sexual transmission of HIV-1 infection. Nat Rev Microbiol. 2003;1:25-34. [ Links ]
4. Mann JM, Tarantola DJ. HIV 1998: The global picture. Sci Am. 1998;279:82-3. [ Links ]
5. Kedzierska K, Crowe SM. The role of monocytes and macrophages in the pathogenesis of HIV-1 infection. Curr Med Chem. 2002;9:1893-903. [ Links ]
6. MacDougall TH, Shattock RJ, Madsen C, Chain BM, Katz DR. Regulation of primary HIV-1 isolate replication in dendritic cells. Clin Exp Immunol. 2002;127:66-71. [ Links ]
7. Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M, et al. Identification of a major co-receptor for primary isolates of HIV-1. Nature. 1996;381:661-6. [ Links ]
8. Dragic T, Litwin V, Allaway GP, Martin SR, Huang Y, Nagashima KA, et al. HIV-1 entry into CD4+ cells is mediated by the chemokine receptor CC-CKR-5. Nature. 1996;381:667-73. [ Links ]
9. Kwon DS, Gregorio G, Bitton N, Hendrickson WA, Littman DR. DC-SIGN-mediated internalization of HIV is required for trans-enhancement of T cell infection. Immunity. 2002;16:135-44. [ Links ]
10. Lambert AA, Gilbert C, Richard M, Beaulieu AD, Tremblay MJ. The C-type lectin surface receptor DCIR acts as a new attachment factor for HIV-1 in dendritic cells and contributes to trans- and cis-infection pathways. Blood. 2008;112:1299-307. [ Links ]
11. Trujillo JR, Rogers R, Molina RM, Dangond F, McLane MF, Essex M, et al. Noninfectious entry of HIV-1 into peripheral and brain macrophages mediated by the mannose receptor. Proc Natl Acad Sci USA. 2007;104:5097-102. [ Links ]
12. Li Q, Estes JD, Schlievert PM, Duan L, Brosnahan AJ, Southern PJ, et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature. 2009;458:1034-8. [ Links ]
13. Buchbinder SP, Katz MH, Hessol NA, O´Malley PM, Holmberg SD. Long-term HIV-1 infection without immunologic progression. AIDS. 1994;8:1123-8. [ Links ]
14. Cao Y, Qin L, Zhang L, Safrit J, Ho DD. Virologic and immunologic characterization of long-term survivors of human immunodeficiency virus type 1 infection. N Engl J Med. 1995;332:201-8. [ Links ]
15. Paxton WA, Martin SR, Tse D, O´Brien TR, Skurnick J, VanDevanter NL, et al. Relative resistance to HIV-1 infection of CD4 lymphocytes from persons who remain uninfected despite multiple high-risk sexual exposure. Nat Med. 1996;2:412-7. [ Links ]
16. Goh WC, Markee J, Akridge RE, Meldorf M, Musey L, Karchmer T, et al. Protection against human immunodeficiency virus type 1 infection in persons with repeated exposure: evidence for T cell immunity in the absence of inherited CCR5 coreceptor defects. J Infect Dis. 1999;179:548-57. [ Links ]
17. Stranford SA, Skurnick J, Louria D, Osmond D, Chang SY, Sninsky J, et al. Lack of infection in HIV-exposed individuals is associated with a strong CD8(+) cell noncytotoxic anti-HIV response. Proc Natl Acad Sci USA. 1999;96:1030-5. [ Links ]
18. Samson M, Libert F, Doranz BJ, Rucker J, Liesnard C, Farber CM, et al. Resistance to HIV-1 infection in caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature. 1996;382:722-5. [ Links ]
19. Liu R, Paxton WA, Choe S, Ceradini D, Martin SR, Horuk R, et al. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell. 1996;86:367-77. [ Links ]
20. Huang Y, Paxton WA, Wolinsky SM, Neumann AU, Zhang L, He T, et al. The role of a mutant CCR5 allele in HIV-1 transmission and disease progression. Nat Med. 1996;2:1240-3. [ Links ]
21. Rugeles MT, Solano F, Díaz FJ, Bedoya VI, Patino PJ. Molecular characterization of the CCR 5 gene in seronegative individuals exposed to human immunodeficiency virus (HIV). J Clin Virol. 2002;23:161-9. [ Links ]
22. Smith MW, Dean M, Carrington M, Winkler C, Huttley GA, Lomb DA, et al. Contrasting genetic influence of CCR2 and CCR5 variants on HIV-1 infection and disease progression. Hemophilia Growth and Development Study (HGDS), Multicenter AIDS Cohort Study (MACS), Multicenter Hemophilia Cohort Study (MHCS), San Francisco City Cohort (SFCC), ALIVE Study. Science. 1997;277:959-65. [ Links ]
23. Winkler C, Modi W, Smith MW, Nelson GW, Wu X, Carrington M, et al. Genetic restriction of AIDS pathogenesis by an SDF-1 chemokine gene variant. ALIVE Study, Hemophilia Growth and Development Study (HGDS), Multicenter AIDS Cohort Study (MACS), Multicenter Hemophilia Cohort Study (MHCS), San Francisco City Cohort (SFCC). Science. 1998;279:389-93. [ Links ]
24. MacDonald KS, Fowke KR, Kimani J, Dunand VA, Nagelkerke NJ, Ball TB, et al. Influence of HLA supertypes on susceptibility and resistance to human immunodeficiency virus type 1 infection. J Infect Dis. 2000;181:1581-9. [ Links ]
25. Mikhail M, Wang B, Saksena NK. Mechanisms involved in non-progressive HIV disease. AIDS Rev. 2003;5:230-44. [ Links ]
26. Hildeman D, Jorgensen T, Kappler J, Marrack P. Apoptosis and the homeostatic control of immune responses. Curr Opin Immunol. 2007;19:516-21. [ Links ]
27. Rojas M, Barrera LF, Puzo G, García LF. Differential induction of apoptosis by virulent Mycobacterium tuberculosis in resistant and susceptible murine macrophages: role of nitric oxide and mycobacterial products. J Immunol. 1997;159:1352-61. [ Links ]
28. Clem RJ, Fechheimer M, Miller LK. Prevention of apoptosis by a baculovirus gene during infection of insect cells. Science. 1991;254:1388-90. [ Links ]
29. Clouston WM, Kerr JF. Apoptosis, lymphocytotoxicity and the containment of viral infections. Med Hypotheses. 1985;18:399-404. [ Links ]
30. Wallace M, Malkovsky M, Carding SR. Gamma/delta T lymphocytes in viral infections. J Leukoc Biol. 1995;58:277-83. [ Links ]
31. Welsh RM, Lin MY, Lohman BL, Varga SM, Zarozinski CC, Selin LK. Alpha beta and gamma delta T-cell networks and their roles in natural resistance to viral infections. Immunol Rev. 1997;159:79-93. [ Links ]
32. Selin LK, Santolucito PA, Pinto AK, Szomolanyi-Tsuda E, Welsh RM. Innate immunity to viruses: Control of vaccinia virus infection by gamma delta T cells. J Immunol. 2001;166:6784-94. [ Links ]
33. DeVico AL, Gallo RC. Control of HIV-1 infection by soluble factors of the immune response. Nat Rev Microbiol. 2004;2:401-13. [ Links ]
34. Sierra S, Kupfer B, Kaiser R. Basics of the virology of HIV-1 and its replication. J Clin Virol. 2005;34:233-44. [ Links ]
35. Cheng-Mayer C, Liu R, Landau NR, Stamatatos L. Macrophage tropism of human immunodeficiency virus type 1 and utilization of the CC-CKR5 coreceptor. J Virol. 1997;71:1657-61. [ Links ]
36. Gonzalez E, Bamshad M, Sato N, Mummidi S, Dhanda R, Catano G, et al. Race-specific HIV-1 disease-modifying effects associated with CCR5 haplotypes. Proc Natl Acad Sci USA. 1999;96:12004-9. [ Links ]
37. Díaz FJ, Vega JA, Patino PJ, Bedoya G, Nagles J, Villegas C, et al. Frequency of CCR5 delta-32 mutation in human immunodeficiency virus (HIV)-seropositive and HIV-exposed seronegative individuals and in general population of Medellin, Colombia. Mem Inst Oswaldo Cruz. 2000;95:237-42. [ Links ]
38. O´Brien SJ, Gao X, Carrington M. HLA and AIDS: a cautionary tale. Trends Mol Med. 2001;7:379-81. [ Links ]
39. Hendel H, Caillat-Zucman S, Lebuanec H, Carrington M, O´Brien S, Andrieu JM, et al. New class I and II HLA alleles strongly associated with opposite patterns of progression to AIDS. J Immunol. 1999;162:6942-6. [ Links ]
40. Kaslow RA, Carrington M, Apple R, Park L, Munoz A, Saah AJ, et al. Influence of combinations of human major histocompatibility complex genes on the course of HIV-1 infection. Nat Med. 1996;2:405-11. [ Links ]
41. Gillespie GM, Kaul R, Dong T, Yang HB, Rostron T, Bwayo JJ, et al. Cross-reactive cytotoxic T lymphocytes against a HIV-1 p24 epitope in slow progressors with B*57. AIDS. 2002;16:961-72. [ Links ]
42. Migueles SA, Sabbaghian MS, Shupert WL, Bettinotti MP, Marincola FM, Martino L, et al. HLA B*5701 is highly associated with restriction of virus replication in a subgroup of HIV-infected long term nonprogressors. Proc Natl Acad Sci USA. 2000;97:2709-14. [ Links ]
43. MacDonald KS, Embree J, Njenga S, Nagelkerke NJ, Ngatia I, Mohammed Z, et al. Mother-child class I HLA concordance increases perinatal human immunodeficiency virus type 1 transmission. J Infect Dis. 1998;177:551-6. [ Links ]
44. Rugeles MT, Shearer GM. Alloantigen recognition in utero: Dual advantage for the fetus? Trends Immunol. 2004;25:348-52. [ Links ]
45. MacDonald KS, Embree JE, Nagelkerke NJ, Castillo J, Ramhadin S, Njenga S, et al. The HLA A2/6802 supertype is associated with reduced risk of perinatal human immunodeficiency virus type 1 transmission. J Infect Dis. 2001;183:503-6. [ Links ]
46. Bedoya VI, Jaimes FA, Delgado JC, Rugeles C, Usuga X, Zapata W, et al. Fetal-maternal HLA-A and -B discordance is associated with placental RNase expression and anti-HIV-1 activity. Curr HIV Res. 2008;6:380-7. [ Links ]
47. O´Brien V. Viruses and apoptosis. J Gen Virol. 1998;79:1833-45. [ Links ]
48. Benedict CA, Norris PS, Ware CF. To kill or be killed: Viral evasion of apoptosis. Nat Immunol. 2002;3:1013-8. [ Links ]
49. Fesq H, Bacher M, Nain M, Gemsa D. Programmed cell death (apoptosis) in human monocytes infected by influenza A virus. Immunobiology. 1994;190:175-82. [ Links ]
50. Jelachich ML, Lipton HL. Theiler´s murine encephalomyelitis virus kills restrictive but not permissive cells by apoptosis. J Virol. 1996;70:6856-61. [ Links ]
51. Crowe SM, Sonza S. HIV-1 can be recovered from a variety of cells including peripheral blood monocytes of patients receiving highly active antiretroviral therapy: A further obstacle to eradication. J Leukoc Biol. 2000;68:345-50. [ Links ]
52. Lewin SR, Kirihara J, Sonza S, Irving L, Mills J, Crowe SM. HIV-1 DNA and mRNA concentrations are similar in peripheral blood monocytes and alveolar macrophages in HIV-1-infected individuals. AIDS.1998;12:719-27. [ Links ]
53. Cassol E, Alfano M, Biswas P, Poli G. Monocyte-derived macrophages and myeloid cell lines as targets of HIV-1 replication and persistence. J Leukoc Biol. 2006;80: 1018-30. [ Links ]
54. Rich EA, Chen IS, Zack JA, Leonard ML, O´Brien WA. Increased susceptibility of differentiated mononuclear phagocytes to productive infection with human immuno- deficiency virus-1 (HIV-1). J Clin Invest. 1992;89:176-83. [ Links ]
55. Sonza S, Maerz A, Uren S, Violo A, Hunter S, Boyle W, et al. Susceptibility of human monocytes to HIV type 1 infection in vitro is not dependent on their level of CD4 expression. AIDS Res Hum Retroviruses. 1995;11:769-76. [ Links ]
56. Lewin SR, Lambert P, Deacon NJ, Mills J, Crowe SM. Constitutive expression of p50 homodimer in freshly isolated human monocytes decreases with in vitro and in vivo differentiation: A possible mechanism influencing human immunodeficiency virus replication in monocytes and mature macrophages. J Virol. 1997;71:2114-9. [ Links ]
57. Naif HM, Li S, Alali M, Sloane A, Wu L, Kelly M, et al. CCR5 expression correlates with susceptibility of maturing monocytes to human immunodeficiency virus type 1 infection. J Virol. 1998;72:830-6. [ Links ]
58. Velilla PA, Hoyos A, Rojas M, Patino PJ, Vélez LA, Rugeles MT. Apoptosis as a mechanism of natural resistance to HIV-1 infection in an exposed but uninfected population. J Clin Virol. 2005;32:329-35. [ Links ]
59. Lehner T, Wang Y, Pido-López J, Whittall T, Bergmeier LA, Babaahmady K. The emerging role of innate immunity in protection against HIV-1 infection. Vaccine. 2008;26:2997-3001. [ Links ]
60. Medzhitov R, Janeway C Jr. Innate immunity. N Engl J Med. 2000;343:338-44. [ Links ]
61. Levy JA, Scott I, Mackewicz C. Protection from HIV/AIDS: The importance of innate immunity. Clin Immunol. 2003;108:167-74. [ Links ]
62. Donaghy H, Pozniak A, Gazzard B, Qazi N, Gilmour J, Gotch F, et al. Loss of blood CD11c(+) myeloid and CD11c(-) plasmacytoid dendritic cells in patients with HIV-1 infection correlates with HIV-1 RNA virus load. Blood. 2001;98:2574-6. [ Links ]
63. Motsinger A, Haas DW, Stanic AK, van Kaer L, Joyce S, Unutmaz D. CD1d-restricted human natural killer T cells are highly susceptible to human immunodeficiency virus 1 infection. J Exp Med. 2002;195:869-79. [ Links ]
64. Soumelis V, Scott I, Gheyas F, Bouhour D, Cozon G, Cotte L, et al. Depletion of circulating natural type 1 interferon-producing cells in HIV-infected AIDS patients. Blood. 2001;98:906-12. [ Links ]
65. Sandberg JK, Fast NM, Palacios EH, Fennelly G, Dobroszycki J, Palumbo P, et al. Selective loss of innate CD4(+) V alpha 24 natural killer T cells in human immunodeficiency virus infection. J Virol. 2002;76:7528-34. [ Links ]
66. Montoya CJ, Velilla PA, Chougnet C, Landay AL, Rugeles MT. Increased IFN-gamma production by NK and CD3+/CD56+ cells in sexually HIV-1-exposed but uninfected individuals. Clin Immunol. 2006;120:138-46. [ Links ]
67. Clerici M. Cell-mediated immunity in HIV infection. AIDS. 1993;7 (Suppl.1):S135-40. [ Links ]
68. Mazzoli S, Trabattoni D, Lo Caputo S, Piconi S, Ble C, Meacci F, et al. HIV-specific mucosal and cellular immunity in HIV-seronegative partners of HIV-seropositive individuals. Nat Med. 1997;3:1250-7. [ Links ]
69. Rowland-Jones SL, Nixon DF, Aldhous MC, Gotch F, Ariyoshi K, Hallam N, et al. HIV-specific cytotoxic T-cell activity in an HIV-exposed but uninfected infant. Lancet. 1993;341:860-1. [ Links ]
70. Kuhn L, Meddows-Taylor S, Gray G, Tiemessen C. Human immunodeficiency virus (HIV)-specific cellular immune responses in newborns exposed to HIV in utero. Clin Infect Dis. 2002;34:267-76. [ Links ]
71. Rowland-Jones S, Sutton J, Ariyoshi K, Dong T, Gotch F, McAdam S, et al. HIV-specific cytotoxic T-cells in HIV-exposed but uninfected Gambian women. Nat Med. 1995;1:59-64. [ Links ]
72. Rowland-Jones SL, Dong T, Dorrell L, Ogg G, Hansasuta P, Krausa P, et al. Broadly cross-reactive HIV-specific cytotoxic T-lymphocytes in highly-exposed persistently seronegative donors. Immunol Lett. 1999;66:9-14. [ Links ]
73. Pinto LA, Sullivan J, Berzofsky JA, Clerici M, Kessler HA, Landay AL, et al. ENV-specific cytotoxic T lymphocyte responses in HIV seronegative health care workers occupationally exposed to HIV-contaminated body fluids. J Clin Invest. 1995;96:867-76. [ Links ]
74. Schenal M, Lo Caputo S, Fasano F, Vichi F, Saresella M, Pierotti P, et al. Distinct patterns of HIV-specific memory T lymphocytes in HIV-exposed uninfected individuals and in HIV-infected patients. AIDS. 2005;19:653-61. [ Links ]
75. Cocchi F, DeVico AL, Garzino-Demo A, Arya SK, Gallo RC, Lusso P. Identification of RANTES, MIP-1 alpha, and MIP-1 beta as the major HIV-suppressive factors produced by CD8+ T cells. Science. 1995;270:1811-5. [ Links ]
76. Walker CM, Moody DJ, Stites DP, Levy JA. CD8+ lymphocytes can control HIV infection in vitro by suppressing virus replication. Science. 1986;234:1563-6. [ Links ]
77. Alfano M, Poli G. Role of cytokines and chemokines in the regulation of innate immunity and HIV infection. Mol Immunol. 2005;42:161-82. [ Links ]
78. Patterson BK, Behbahani H, Kabat WJ, Sullivan Y, O´Gorman MR, Landay A, et al. Leukemia inhibitory factor inhibits HIV-1 replication and is upregulated in placentae from nontransmitting women. J Clin Invest. 2001;107:287-94. [ Links ]
79. Lee-Huang S, Huang PL, Sun Y, Kung HF, Blithe DL, Chen HC. Lysozyme and RNases as anti-HIV components in beta-core preparations of human chorionic gonadotropin. Proc Natl Acad Sci U S A. 1999;96:2678-81. [ Links ]
80. Quinones-Mateu ME, Lederman MM, Feng Z, Chakraborty B, Weber J, Rangel HR, et al. Human epithelial beta-defensins 2 and 3 inhibit HIV-1 replication. AIDS. 2003;17:F39-48. [ Links ]
81. Chang TL, Vargas J Jr, Del Portillo A, Klotman ME. Dual role of alpha-defensin-1 in anti-HIV-1 innate immunity. J Clin Invest. 2005;115:765-73. [ Links ]
82. Garcia JR, Krause A, Schulz S, Rodríguez-Jiménez FJ, Kluver E, Adermann K, et al. Human beta-defensin 4: A novel inducible peptide with a specific salt-sensitive spectrum of antimicrobial activity. FASEB J. 2001;15:1819-21. [ Links ]
83. Ayabe T, Satchell DP, Wilson CL, Parks WC, Selsted ME, Ouellette AJ. Secretion of microbicidal alpha-defensins by intestinal Paneth cells in response to bacteria. Nat Immunol. 2000;1:113-8. [ Links ]
84. Valore EV, Park CH, Quayle AJ, Wiles KR, McCray PB Jr., Ganz T. Human beta-defensin-1: an antimicrobial peptide of urogenital tissues. J Clin Invest. 1998;101:1633-42. [ Links ]
85. Trabattoni D, Caputo SL, Maffeis G, Vichi F, Biasin M, Pierotti P, et al. Human alpha defensin in HIV-exposed but uninfected individuals. J Acquir Immune Defic Syndr. 2004;35:455-63. [ Links ]
86. Ganz T. Defensins:A peptides of vertebrates. C R Biol. 2004;327:539-49. [ Links ]
87. Schutte BC, Mitros JP, Bartlett JA, Walters JD, Jia HP, Welsh MJ, et al. Discovery of five conserved beta -defensin gene clusters using a computational search strategy. Proc Natl Acad Sci USA. 2002;99:2129-33. [ Links ]
88. Jurevic RJ, Bai M, Chadwick RB, White TC, Dale BA. Single-nucleotide polymorphisms (SNPs) in human beta-defensin 1: High-throughput SNP assays and association with Candida carriage in type I diabetics and nondiabetic controls. J Clin Microbiol. 2003;41:90-6. [ Links ]
89. Jurevic RJ, Chrisman P, Mancl L, Livingston R, Dale BA. Single-nucleotide polymorphisms and haplotype analysis in beta-defensin genes in different ethnic populations. Genet Test. 2002;6:261-9. [ Links ]
90. Zapata W, Rodríguez B, Weber J, Estrada H, Quinones-Mateu ME, Zimermman PA, et al. Increased levels of human beta-defensins mRNA in sexually HIV-1 exposed but uninfected individuals. Curr HIV Res. 2008;6:531-8. [ Links ]

