










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiomédica
Print version ISSN 0120-4157
Biomédica vol.33 no.2 Bogotá Apr./June 2013
https://doi.org/10.7705/biomedica.v33i2.790
doi: http://dx.doi.org/10.7705/biomedica.v33i2.790
ARTÍCULO ORIGINAL
Dulfary Sánchez 1,2,3 , Céline Lefebvre 4, Luis F. García 1,2, Luis F. Barrera 1,2
1 Grupo de Inmunología Celular e Inmunogenética (GICIG), Instituto de Investigaciones Médicas, Facultad de Medicina, Universidad de Antioquia, Medellín, Colombia
2 Centro Colombiano de Investigación en Tuberculosis (CCITB), Medellín, Colombia
3 Stanley S. Scott Cancer Center, Health Science Center, Louisiana State University, New Orleans, LA, United States
4ECOGENE-21 Clinical Trial Center, Centre de Médecine Génique Communautaire de l'Université de Montréal Centre Hospitalier Affilié Universitaire Régional de Chicoutimi, Pavillon Notre-Dame, Chicoutimi, Canada
Instituciones donde se llevó a cabo la investigación:
Universidad de Antioquia, Medellín, Colombia Laboratory for Genetics and Genomic Medicine of Inflammation, Montreal, Canada
Contribución de los autores:
Luis Fernando Barrera y Luis Fernando García fueron los responsables del diseño del proyecto y obtuvieron su financiación.
María Dulfary Sánchez y Céline Lefebvre procesaron y analizaron las muestras de ADN para el estudio de los polimorfismos mediante espectrometría de masas.
Todos los autores participaron en el análisis de datos, redacción y edición del artículo.
Recibido: 08/08/12; aceptado:19/12/12
Introduction: Interferon gamma (IFN ? ) is the most potent cytokine involved in the control of Mycobacterium tuberculosis ( Mtb ), the etiological agent of human tuberculosis (TB). Patients with active TB present reduced levels of IFN ? , which may explain the lack of effective immunity against Mtb in these patients. The diminished expression of or functional alterations in trans-acting factors that regulate IFN ? gene expression may explain the reduced levels of IFN ? in TB patients.
Objective: To investigate the relationships of genetic variants in the transcription factors TBET, STAT1, STAT4, and HLX to susceptibility/resistance to pulmonary TB.
Materials and methods: Eight candidate single-nucleotide polymorphisms (SNPs) were selected, and genotyped in 466 unrelated pulmonary TB patients and 300 healthy controls from Colombia, and the allelic and genetic associations with TB were analyzed.
Results: The results indicate that no SNP in the transcription factors studied is associated with TB.
However, polymorphism rs11650354 in the TBET gene may be associated with a decreased risk of TB; the TT genotype was significantly associated with TB protection in a recessive genetic model (OR=0.089, 95% CI: 0.01-0.73, p=0.0069), although this association was not maintained after multiple test correction (EMP2= 0.61).
Conclusion: In this study, the rs11650354 variant of TBET was suggested to promote resistance to TB in a Colombian population. A future replication case-control study using additional samples will be necessary to confirm this suggestive association.
Keywords: Mycobacterium tuberculosis , interferon gamma, transcription factor, STAT1, STAT4, casecontrol studies.
doi: http://dx.doi.org/10.7705/biomedica.v33i2.790
Variantes en los factores de transcripción para IFN ?, TBET, STAT1 , STAT4 y HLX , y el riesgo de desarrollar tuberculosis pulmonar en un estudio de casos y controles de una población colombiana
Introducción. El interferón gama (IFN ? ) es la citocina más potente para controlar la infección por Mycobacterium tuberculosis , el agente etiológico de la tuberculosis humana. Los pacientes con tuberculosis activa presentan reducción de los niveles de IFN ? , lo cual parece explicar la inmunidad poco efectiva contra el bacilo. La disminución de su expresión o alteraciones funcionales de los factores transactivadores del promotor del gen de IFN ? , podrían explicar la reducción de los niveles de IFN ? en los pacientes con tuberculosis.
Objetivo. Determinar la asociación de variantes genéticas en los factores de transcripción TBET STAT1, STAT4 y HLX con sensibilidad o resistencia a tuberculosis pulmonar.
Materiales y métodos. Se seleccionaron ocho polimorfismos de un solo nucleótido ( Single-Nucleotide Polymorphism , SNP) y se estableció su genotipo, en 466 pacientes con tuberculosis pulmonar y 300 controles sanos en Colombia; además, se hizo un análisis de asociación alélica y genética.
Resultados. Los resultados indican que los SNP de los factores de transcripción estudiados no están asociados con tuberculosis; sin embargo, el polimorfismo rs11650354 en TBET puede estar implicado en la disminución de riesgo de tuberculosis. El genotipo TT de TBET se asoció significativamente con protección contra tuberculosis usando un modelo genético recesivo (OR=0,089; CI 95% : 0,01-0,73; p=0,0069); sin embargo, la corrección mediante pruebas múltiples de ajuste abolió esta asociación ( Empirical P Value, EMP2=0,61).
Conclusión. En este estudio se sugiere un efecto de la variante rs11650354 de TBET sobre la resistencia a la tuberculosis en la población colombiana. Es necesario desarrollar un estudio de replicación usando muestras adicionales para confirmar esta asociación sugestiva.
Palabras clave: Mycobacterium tuberculosis , interferón-gamma, factor de transcripción STAT1, factor de transcripción STAT4, estudios de casos y controles.
doi: http://dx.doi.org/10.7705/biomedica.v33i2.790
It is estimated that one-third of the human population is infected with Mycobacterium tuberculosis ( Mtb ) the etiological agent of tuberculosis (TB). TB is a complex disease, and its occurrence is influenced by environmental, immunologic and genetic factors. It is estimated that 10% of Mtb -infected individuals may develop active TB during their lifetimes (1), suggesting that the majority of the infected individuals are endowed with a protective immune response.
IFN ? is a potent cytokine with pleiotropic activities in different cell types, and it is considered the major activator of macrophages, a cell type that is critical to the control of Mtb replication and dissemination. In addition, IFN ? acts on professional antigen presenting cells, increasing MHC class II expression and thus favoring the adaptive T cell response. IFN ? is produced by different cell populations, including Natural Killer (NK) cells (2), T lymphocytes (LT) a ß (CD4 + and CD8 + T cells), LT ? d cells (3), dendritic cells (DCs) (4), and monocytes/macrophages (5).
IFN ? activity is crucial for mycobacterial control in Mtb -infected cells. It stimulates the production of inducible nitric oxide synthase (NOS-2), which is known to be responsible for high levels of nitric oxide, and other reactive nitrogen intermediates that are bactericidal to Mtb in mice (6), although the role of NOS-2 in human TB remains controversial. IFN ? also induces the expression of LRG-47, a p47 GTPase that controls Mtb by allowing the autophagy of infected macrophages (7). The importance of IFN ? during Mtb infection has been demonstrated in both mice and humans. Mice that are deficient in IFN ? or the IFN ? receptor (IFN ? R) rapidly succumb to infection with Mtb or M. bovis BCG (8,9). Individuals with mutations in the IFNGR1 or IFNGR2 genes (encoding the IFN ? -R1 and R2 receptors, respectively) or IFN ? -intracellular pathway components such as the STAT1 protein may experience fatal disseminated BCG or non-tuberculous mycobacterial infection during childhood (10). Polymorphic variants in the IFNG and IFNG receptor genes have been associated with variable levels of IFN ? production (11,12).
Different reports indicate that peripheral blood mononuclear cells (PBMCs) from patients with severe active TB produce lower levels of IFN ? protein and mRNA in response to mycobacterial antigens than do healthy tuberculin reactor subjects (13,14) or with household healthy contacts of TB patients (15). This impaired IFN ? production could be associated with an inability to clear the bacterial load and the consequent progress of the disease and its pathology. This assumption is consistent with evidence suggesting that IFN ? expression correlates with protective immunity against TB (6,8,9); however, according to other studies, high levels of IFN ? protein post-infection represents a risk factor for developing active TB (16-20). These data suggest that the mechanisms that control the excessive inflammatory responses during Mtb infection may have a critical role in the immunopathology of TB.
The molecular regulation of IFN ? is a complex process because multiple signals converge to govern its expression (21,22). Different regulatory regions of the IFN ? locus have been identified, and multiple transcription factors have been implicated in the regulation of IFN ? gene expression (23). Transcription factors such as TBET (T-box 21) (24), GATA3 (25), STAT1 (Signal Transducers and Activators of Transcription 1) (26), STAT4 (27) HLX ( H2.0-like homeobox) (28) , CREB (29), ATF-2 c-Jun (30), and NF- ? B (22) have been implicated in the regulation of IFN ? gene expression. Few studies have tested whether the reduced IFN ? mRNA and protein production in human TB is due to functional mutations in any of these transcription factors. Reduced CREB protein expression or the absence of phosphorylated nuclear CREB in TB patients resulted in decreased IFN ? promoter activity and reduced IFN ? production (29). However, it is presently unknown whether genetic variants of this and other transcription factors may be associated with susceptibility/resistance to TB.
IFN ? plays a critical role during Mtb infection and there is evidence indicating the crucial role of regulation of IFN ? production. Therefore, we performed a case-control gene association study in a Colombian population of pulmonary TB cases and healthy controls, using transcription factors that positively or negatively regulate IFNG gene expression as the candidate associated genes. A trend for association with a decreased risk for TB (OR±95%CI=0.089±0.01-0.73; ? 2 =7.99; p=0.0069) was found for the TT genotype of TBET (rs11650354 SNP) assuming a recessive model of inheritance; however, this significance was lost after multiple test correction via permutation (EMP=0.61). The populations were also genotyped for additional single nucleotide polymorphisms in the STAT1 , STAT4 , and HLX genes; however none of these SNPs was found to be associated with susceptibility/resistance to TB.
Materials and methods
Subjects
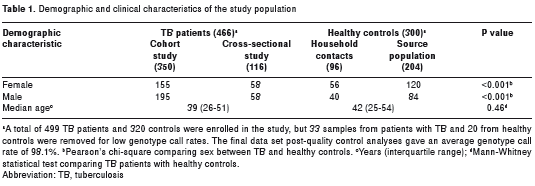
Pulmonary TB patients (n=499) and healthy controls (n=320) of Mestizo ethnicity were recruited from the metropolitan area of Medellín, the capital city of the Antioquia Department, Colombia. The incidence of tuberculosis in Colombia was last reported as 34 per 100,000 inhabitants in 2010, according to a World Bank report released in 2011 (31). In Medellin, the records of the incidence of TB show rates ranging from 40.4 to 60.7 cases per 100,000 inhabitants (32) . The cases included in this study were newly diagnosed TB patients with disease confirmed by sputum smear examinations for acidfast bacilli and/or culture for Mtb. The cases were enrolled from February 2002 to September 2004 (from a cross-sectional study; n=119) and from March 2005 to November 2006 (from a cohort study (20); n=380). The control subjects were healthy (as defined by Interferon Gamma Release Assays, IGRA), non-related household contacts enrolled (collected from March 2005 to December 2006; n=105), and non-contact individuals living in the same neighborhood as the TB patients (source population), randomly sampled and age-matched to the TB patients (n=215), enrolled from September 2007 to December 2008. The exclusion criteria for patients included positive serologic tests for Human Immunodeficiency Virus (HIV), a history of organ transplantation, primary immunodeficiency cancer, treatment with immunosuppressive drugs endocrine disorders such as diabetes, autoimmune and chronic renal disease, or the development of pleural, miliary or meningeal TB. The exclusion criteria for controls included persistent productive cough (for more than 2 weeks) and a previous history of TB. No household contact participating in this study showed clinical symptoms of TB during the 2 years of clinical follow-up. All subjects signed an informed consent document that was approved by the Ethics Committee of the Facultad de Medicina Universidad de Antioquia. table 1 shows the demographic characteristics of the 466 TB patients and 300 healthy controls that were included in the association analyses. Samples from 33 TB patients and 20 healthy controls were eliminated during the genotype quality control analyses, as described below.
Blood samples and DNA isolation
Venous blood samples (3-8 ml in EDTA; BD Vacutainers) were collected. Genomic DNA was extracted using the DNAzol reagent (ES Extra Strength MCR, Cincinnati, OH, USA) according to the manufacturer's instructions. The isolated DNA was suspended in TE 1X (pH 7.9) and stored at 4°C. DNA integrity was visualized by ethidium bromide staining of 0.7% agarose (Amresco; Solon OH, USA) gels. DNA samples were quantified using the Quant-iT PicoGreen dsDNA reagent (Invitrogen Molecular Probes, Eugene, OR, USA) as recommended by the manufacturer.
SNP selection and genotyping
Eight SNPs in the TBET , HLX, GATA3, STAT1, and STAT4 genes were evaluated. The SNPs were strategically selected according to their locations in areas of potential importance for gene regulation or protein function and/or regions that are in LD with other SNPs previously reported to be associated with different diseases. The SNPs rs11578466 and rs11588625 in the HLX gene were used as proxies for an HLX SNP that lies within the 3' untranslated region (HLX1-C/T, 3' UTR; NCBI dbSNP: rs2738756). SNP genotype information for the CEU population was downloaded from HapMap database (phase II release 21a from January 2007). Primer design software (MassARRAY Assay Design) was used to design PCR and hME primers for the high-plexed MassEXTEND (hME) assays to avoid overlapping mass signals in the available spectrum range for each target SNP. Samples were genotyped at the Laboratory for Genetics and Genomic Medicine of Inflammation (http://www.inflammgen.org), Université de Montreal, Montreal Canada, by MassARRAY (Sequenom, San Diego CA, USA); this technique uses chip-based Matrix Assisted Laser Desorption/Ionization Time-of-Flight (MALDI-TOF) mass spectrometry, as previously described (33).
Data processing
To optimize the quality of the data (quality control QC) while keeping as many SNPs and samples as possible, we selected different parameter thresholds using PLINK v1.04 (34). The PLINK QC options were set to the following thresholds: maximum missing genotypes per person (--mind option) =0.20, maximum missing genotypes per SNP (--geno option) =0.10 and Hardy–Weinberg Equilibrium (HWE; exact) P-value <0.05 in controls (--hwe option). These QC steps produced a final high-quality dataset with an average call rate of more than 80% per individual and per SNP. Of the initial DNA samples from 499 TB patients and 320 controls, 33 samples from TB patients and 20 from healthy controls were removed due to low genotype call rates. The final post-QC analysis dataset yielded an average genotype call rate of 98.1%.
Statistical analysis
Association testing was performed with PLINK software v1.04. A single marker basic allelic association test (--assoc --Fisher option) and genotypic association test (--model --Fisher option) were performed for each of the post-QC SNPs. To test for the mode of inheritance, 2×3 tables, Fisher's exact test and ORs with 95% confidence intervals (CI) were calculated. For SNPs that showed a significant genotypic association (p<0.05), a PLINK's max(T) permutation procedure (--mperm option) with 100,000 iterations was performed to limit type II error. PLINK generates both uncorrected (EMP1) and corrected (EMP2) empirical P-values. In both cases, p-values of <0.05 were considered significant.
The Mann-Whitney statistical test was used for age comparisons, and the ? 2 test was used to test for gender differences between cases and controls.
To adjust for potential confounding, gender was included in a forward stepwise logistic regression analysis for SNPs with significant genotypic association using the Statistical Package for the Social Sciences (SPSS v.11.5 for Windows; Chicago, IL). Statistically significant associations were defined by a p=0.05.
Results
The demographic and clinical characteristics of the population analyzed in our study are shown in table 1. Of the initial 499 TB patients and 320 controls, the DNA samples from 33 TB patients and 20 healthy controls were removed from the analysis due to low genotype call rates. There was no difference in age distribution between cases and controls (p=0.46) but men were more frequent in the TB patient group than in the control group (p<0.001). Two SNPs in GATA3 (rs4143094 and rs1058240) showed deviations from Hardy-Weinberg Equilibrium (HWE) in the control group and were removed for further analysis. The remaining 8 SNPs tested passed QC analysis (genotyping success <80%) and were included in the subsequent analyses.
The results of the allelic association analyses did not reveal any differences in the frequencies of TBET, HLX, STAT1, and STAT4 polymorphisms between TB cases and controls (table 2). Genotype frequency analysis was also performed (table 3) and a significant difference between TB patients and controls was found in TBET rs11650354 SNP, with genotypic frequencies significantly decreased in cases compared with controls ( ? 2 =8.06; P=0.019).


To analyze the genotypic effect, we performed a comparison of genotypic models in which the risk allele was dominant or recessive. The frequency of the TT genotype of TBET (recessive model of inheritance) was significantly lower in TB patients (OR±95%CI= 0.089±0.01-0.73; ? 2 =7.99; p=0.0069) than in controls. This difference persisted even after adjusting for the gender distribution between groups (p=0.02), as shown using a logistic regression analysis controlling for gender as a covariate. Genotypic significance was ascertained through a permutation of phenotypes (n=100,000 permutations) over all 81 SNPs (the total number of SNPs included in the complete genetic study using the same Colombian samples ((35) and unpublished data) in the analysis. After these stringently corrected permuted p-values were applied, the rs11650354 SNP did not remain significantly associated with TB protection (EMP2 p=0.61; table 3).
Discussion
Studies in the genetics of TB resistance/susceptibility have identified several important candidate genes (discussed in (36)) and genetic regions (37). IFN ? has been identified as a key cytokine in the control of mycobacterial infections, as clearly demonstrated in mice models of infection (38,39) as well as in human genetic studies evaluating polymorphisms in the IFNG gene (40-42) and other genes involved in the modulation of IFN ? production or signaling (43,44).
Under the hypothesis that IFN ? production is critical for the successful control of Mtb infection and multiplication, we studied SNPs from five specific genes that control the transcription of the IFN ? gene, namely, TBET (24), GATA3 (25), STAT1 (26) STAT4 (27), and HLX (28), and assessed whether their variants were associated with susceptibility/ resistance to TB. We found that the TT genotype of TBET is associated with a decreased risk of TB in the Colombian population studied, although this association was lost after multiple test correction. A study with larger numbers of samples will be necessary to confirm this association. TBET is a member of the T-box family of transcription factors that regulate the lineage commitment of T helper (Th)-0 lymphocytes toward a predominant Th1 phenotype. Different studies have highlighted the importance of TBET in the diverse subsets of cells that have been implicated in a Th1 immune response such as dendritic cells (45), NK cells (46), and CD8 + LT cells (47). The important regulatory role of TBET in the course of Mtb infection was established in a murine model. Mice that lack TBET due to a targeted mutation of this gene are more susceptible to Mtb infection than their wild-type (WT) counterparts. These mice exhibited a shorter mean survival time, less efficient control of bacterial replication and severe lung inflammation following both i.v. and respiratory infection (48). The lack of TBET expression resulted in a weakening of the systemic immune response to M. tuberculosis , evidenced by higher bacterial burdens in the lung, liver, and spleen. In addition, IFN ? production in response to a mycobacterial purified protein derivative (PPD) was significantly reduced in T cells isolated from TB lung lesions from TBET -/- mice (48).
However, it has been suggested that a disruption of the immune response profile balance during active TB facilitates an overregulation of the effector response, characterized by decreased IFN ? production (49), increased IL-4 production (50) and the development of regulatory T cells (Tregs) that inhibit protective Th1 responses (51). A lack of TBET expression generates conditions that strongly favor the development of the Th2 compartment (47). Heritable Th2-associated conditions might be caused by genetic variation in Th1 cytokine regulation through TBET (52). TBET polymorphisms have recently been associated with susceptibility to asthma (53,54), diabetes mellitus (55), autoimmune diseases (56,57), cancer (58,59) and viral infections (60,61). However, to date, no polymorphism in TBET has been associated with mycobacterial disease, including TB. The present report is the first to suggest an association between a TBET variant (rs11650354) and tuberculosis in a Colombian population.
The findings of recent genetic studies suggest an association between the rs11650354 SNP in TBET and systemic sclerosis (56). These authors showed that patients with the TT genotype had a more prominent Th2 cytokine profile, while patients in the wild-type group ( CC ) had a more prominent proinflammatory cytokine profile. They thus hypothesized that CD4 + T cells fail to differentiate into the Th1 lineage in the presence of the TBET TT polymorphism, leading to a Th2-predominant environment. This finding supports the potential importance of Th1/Th2 cytokine balance in autoimmune disease and asthma; this Th1/Th2 dysregulation could also be expected to affect the response to TB and potentially explains our finding of the TT genotype as a protective factor against TB development.
There is not enough evidence regarding whether specific variants of the TBET gene are associated with TBET expression/function. Interestingly the levels of TBET messenger RNA (mRNA) in peripheral blood cells have been reported to be lower in asthma patients than in controls (62), and two TBET variants (rs11650354 and rs17699436) have been associated with susceptibility to asthma (63,64), suggesting that TBET SNPs could affect TBET transcription. To better understand the immunologic significance of the TBET variant rs11650354 in TB, further studies need to be undertaken to demonstrate the effects of these polymorphisms on mRNA and protein levels and TBET function, which in turn could affect IFN ? transcription and disturb the immune response.
In summary, this study found evidence of a weak association between the TBET TT genotype (rs11650354) and resistance to pulmonary TB. However, the lack of evidence for this association after multiple test correction via permutation casts doubts on the robustness of this finding. A further replication case-control study will be needed to confirm this result.
Acknowledgements
This study would not be possible without the generous participation of the patients, their families and healthy controls.
Conflicts of interest
The authors have declared that no competing interest exists.
Funding
This work was funded by grants 11150412950 RC-431-2004, 1115408204820, and 11150416335- 2004 from Colciencias, Bogotá, Colombia.
Corresponding author: María Dulfary Sánchez, Stanley S. Scott Cancer Center, Health Science Center, Louisiana State University, 533 Bolivar Street Laboratory 452-455, New Orleans, LA 70112, USA Phone: (504) 875-9653 mdsanchezp@gmail.com
1. Bleed D, Dye C, Raviglione MC. Dynamics and control of the global tuberculosis epidemic. Curr Opin Pulm Med. 2000;6:174-9. [ Links ]
2. Garcia VE, Uyemura K, Sieling PA, Ochoa MT, Morita CT, Okamura H, et al. IL-18 promotes type 1 cytokine production from NK cells and T cells in human intracellular infection. J Immunol. 1999;162:6114-21. [ Links ]
3. Sharma S, Bose M. Role of cytokines in immune response to pulmonary tuberculosis. Asian Pac J Allergy Immunol. 2001;19:213-9. [ Links ]
4. Fricke I, Mitchell D, Mittelstadt J, Lehan N, Heine H, Goldmann T, et al. Mycobacteria induce IFN-gamma production in human dendritic cells via triggering of TLR2. J Immunol. 2006;176:5173-82. [ Links ]
5. Fenton MJ, Vermeulen MW, Kim S, Burdick M, Strieter RM, Kornfeld H. Induction of gamma interferon production in human alveolar macrophages by Mycobacterium tuberculosis. Infect Immun. 1997;65:5149-56. [ Links ]
6. Cooper AM, Adams LB, Dalton DK, Appelberg R, Ehlers S. IFN-gamma and NO in mycobacterial disease: New jobs for old hands. Trends Microbiol. 2002;10:221-6. http://dx.doi.org/10.1016/S0966-842X(02)02344-2 [ Links ]
7. Gutiérrez MG, Master SS, Singh SB, Taylor GA, Colombo MI, Deretic V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell. 2004;119:753-66. http://dx.doi.org/10.1016/j.cell.2004.11.038 [ Links ]
8. Dalton DK, Pitts-Meek S, Keshav S, Figari IS, Bradley A, Stewart TA. Multiple defects of immune cell function in mice with disrupted interferon-gamma genes. Science. 1993;259:1739-42. http://dx.doi.org/10.1126/science.8456300 [ Links ]
9. Kamijo R, Le J, Shapiro D, Havell EA, Huang S, Aguet M, et al. Mice that lack the interferon-gamma receptor have profoundly altered responses to infection with Bacillus Calmette-Guerin and subsequent challenge with lipopolysaccharide. J Exp Med. 1993;178:1435-40. [ Links ]
10. Ottenhoff TH, Verreck FA, Hoeve MA, van de Vosse E. Control of human host immunity to mycobacteria. Tuberculosis. 2005;85:53-64. http://dx.doi.org/10.1016/j.tube.2004.09.011 [ Links ]
11. Pravica V, Asderakis A, Perrey C, Hajeer A, Sinnott PJ, Hutchinson IV. In vitro production of IFN-gamma correlates with CA repeat polymorphism in the human IFN-gamma gene. Eur J Immunogenet. 1999;26:1-3. http://dx.doi.org/10.1046/j.1365-2370.1999.00122.x [ Links ]
12. Pravica V, Perrey C, Stevens A, Lee JH, Hutchinson IV. A single nucleotide polymorphism in the first intron of the human IFN-gamma gene: Absolute correlation with a polymorphic CA microsatellite marker of high IFN-gamma production. Hum Immunol. 2000;61:863-6. http://dx.doi.org/10.1016/S0198-8859(00)00167-1 [ Links ]
13. Sánchez FO, Rodríguez JI, Agudelo G, García LF. Immune responsiveness and lymphokine production in patients with tuberculosis and healthy controls. Infect Immun. 1994;62:5673-8. [ Links ]
14. Hirsch CS, Toossi Z, Othieno C, Johnson JL, Schwander SK, Robertson S, et al. Depressed T-cell interferon-gamma responses in pulmonary tuberculosis: Analysis of underlying mechanisms and modulation with therapy. J Infect Dis. 1999;180:2069-73. http://dx.doi.org/10.1086/315114 [ Links ]
15. Rueda CM, Marín ND, García LF, Rojas M. Characterization of CD4 and CD8 T cells producing IFN-gamma in human latent and active tuberculosis. Tuberculosis (Edinb). 2010;90:346-53. http://dx.doi.org/10.1016/j.tube.2010.09.003 [ Links ]
16. Fonseca DM, Silva CL, Paula MO, Soares EG, Marchal G, Horn C, et al. Increased levels of interferon-gamma primed by culture filtrate proteins antigen and CpG-ODN immunization do not confer significant protection against Mycobacterium tuberculosis infection. Immunology. 2007;121:508-17. http://dx.doi.org/10.1111/j.1365-2567.2007.02597.x [ Links ]
17. Lin PL, Rodgers M, Smith L, Bigbee M, Myers A, Bigbee C, et al. Quantitative comparison of active and latent tuberculosis in the cynomolgus macaque model. Infect Immun. 2009;77:4631-42. [ Links ]
18. Doherty TM, Demissie A, Olobo J, Wolday D, Britton S, Eguale T, et al. Immune responses to the Mycobacterium tuberculosis -specific antigen ESAT-6 signal subclinical infection among contacts of tuberculosis patients. J Clin Microbiol. 2002;40:704-6. http://dx.doi.org/10.1128/JCM.40.2.704-706.2002 [ Links ]
19. Winek J, Rowinska-Zakrzewska E, Demkow U, Szopinski J, Szolkowska M, Filewska M, et al. Interferon gamma production in the course of Mycobacterium tuberculosis infection. J Physiol Pharmacol. 2008;59(Suppl.6):751-9. [ Links ]
20. del Corral H, París SC, Marín ND, Marín DM, López L, Henao HM, et al. IFNgamma response to Mycobacterium tuberculosis , risk of infection and disease in household contacts of tuberculosis patients in Colombia. PloS One. 2009;4:e8257. [ Links ]
21. Schoenborn JR, Wilson CB. Regulation of interferongamma during innate and adaptive immune responses. Adv Immunol. 2007;96:41-101. http://dx.doi.org/10.1016/S0065-2776(07)96002-2 [ Links ]
22. Balasubramani A, Shibata Y, Crawford GE, Baldwin AS, Hatton RD, Weaver CT. Modular utilization of distal cisregulatory elements controls Ifng gene expression in T cells activated by distinct stimuli. Immunity. 2010;33:35-47. http://dx.doi.org/10.1016/j.immuni.2010.07.004 [ Links ]
23. Young HA. Unraveling the pros and cons of interferongamma gene regulation. Immunity. 2006;24:506-7. http://dx.doi.org/10.1016/j.immuni.2006.05.004 [ Links ]
24. Szabo SJ, Kim ST, Costa GL, Zhang X, Fathman CG, Glimcher LH. A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell. 2000;100:655-69. http://dx.doi.org/10.1016/S0092-8674(00)80702-3 [ Links ]
25. Usui T, Nishikomori R, Kitani A, Strober W. GATA-3 suppresses Th1 development by downregulation of Stat4 and not through effects on IL-12Rbeta2 chain or T-bet. Immunity. 2003;18:415-28. http://dx.doi.org/10.1016/S1074-7613(03)00057-8 [ Links ]
26. Agnello D, Lankford CS, Bream J, Morinobu A, Gadina M, O'Shea JJ, et al. Cytokines and transcription factors that regulate T helper cell differentiation: new players and new insights. J Clin Immunol. 2003;23:147-61. [ Links ]
27. Nakahira M, Ahn HJ, Park WR, Gao P, Tomura M, Park CS, et al. Synergy of IL-12 and IL-18 for IFN-gamma gene expression: IL-12-induced STAT4 contributes to IFNgamma promoter activation by up-regulating the binding activity of IL-18-induced activator protein 1. J Immunol. 2002;168:1146-53. [ Links ]
28. Becknell B, Hughes TL, Freud AG, Blaser BW, Yu J, Trotta R, et al. Hlx homeobox transcription factor negatively regulates interferon-gamma production in monokineactivated natural killer cells. Blood. 2007;109:2481-7. [ Links ]
29. Samten B, Ghosh P, Yi AK, Weis SE, Lakey DL, Gonsky R, et al. Reduced expression of nuclear cyclic adenosine 5'-monophosphate response element-binding proteins and IFN-gamma promoter function in disease due to an intracellular pathogen. J Immunol. 2002;168:3520-6. [ Links ]
30. Samten B, Townsend JC, Weis SE, Bhoumik A, Klucar P, Shams H, et al. CREB, ATF, and AP-1 transcription factors regulate IFN-gamma secretion by human T cells in response to mycobacterial antigen. J Immunol. 2008;181:2056-64. [ Links ]
31. Word Bank Report . Incidence of tuberculosis (per 100,000 people). Fecha de consulta: 9 de diciembre de 2012. Disponible en: http://data.worldbank.org/indicator/SH.TBS.INCD/countries. [ Links ]
32. Secretaría de Salud de Medellín . Boletín Número 5 de 2011. Fecha de consulta: 9 de diciembre de 2012. Disponible en: http://www.medellin.gov.co/irj/go/km/docs/wpccontent/Sites/Subportal%20del%20Ciudadano/Salud/Secciones/Indicadores%20y%20Estad%C3%ADsticas/D o c u m e n t o s/2 0 11 /B o l e t % C 3 % A D n % 2 0 Epidemiol%C3%B3gico%20Sivigila/Boletines%202011/Bolet%C3%ADn%20Epidemiol%C3%B3gico%205%20-%202011.pdf . [ Links ]
33. Storm N, Darnhofer-Patel B, van den Boom D, Rodi CP. MALDI-TOF mass spectrometry-based SNP genotyping. Methods Mol Biol. 2003;212:241-62. [ Links ]
34. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81:559-75. http://dx.doi.org/10.1086/519795 [ Links ]
35. Sánchez D, Lefebvre C, Rioux J, García LF, Barrera LF. Evaluation of Toll-like receptor and adaptor molecule polymorphisms for susceptibility to tuberculosis in a Colombian population. Int J Immunogenet. 2012;39:216-23. http://dx.doi.org/10.1111/j.1744-313X.2011.01077.x. [ Links ]
36. Cooke GS, Hill AV. Genetics of susceptibility to human infectious disease. Nat Rev Genet. 2001;2:967-77. http://dx.doi.org/10.1038/35103577 [ Links ]
37. Bellamy R. Genome-wide approaches to identifying genetic factors in host susceptibility to tuberculosis. Microbes Infect. 2006;8:1119-23. http://dx.doi.org/10.1016/j.micinf.2005.10.025 [ Links ]
38. Flynn JL, Chan J, Triebold KJ, Dalton DK, Stewart TA, Bloom BR. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J Exp Med. 1993;178:2249-54. [ Links ]
39. Suzue K, Asai T, Takeuchi T, Koyasu S. In vivo role of IFNgamma produced by antigen-presenting cells in early host defense against intracellular pathogens. Eur J Immunol. 2003;33:2666-75. http://dx.doi.org/10.1002/eji.200323292 [ Links ]
40. Amim LH, Pacheco AG, Fonseca-Costa J, Loredo CS, Rabahi MF, Melo MH, et al. Role of IFN-gamma +874 T/A single nucleotide polymorphism in the tuberculosis outcome among Brazilians subjects. Mol Biol Rep. 2008;35:563-6. [ Links ]
41. Pacheco AG, Cardoso CC, Moraes MO. IFNG +874T/A IL10 -1082G/A and TNF -308G/A polymorphisms in association with tuberculosis susceptibility: A meta-analysis study. Hum Genet. 2008;123:477-84 [ Links ]
42. Henao MI, Montes C, París SC, García LF. Cytokine gene polymorphisms in Colombian patients with different clinical presentations of tuberculosis. Tuberculosis (Edinb). 2006;86:11-9. http://dx.doi.org/10.1016/j.tube.2005.03.001 [ Links ]
43. Ottenhoff TH, De Boer T, van Dissel JT, Verreck FA. Human deficiencies in type-1 cytokine receptors reveal the essential role of type-1 cytokines in immunity to intracellular bacteria. Adv Exp Med Biol. 2003;531:279-94. [ Links ]
44. Wang J, Tang S, Shen H. Association of genetic polymorphisms in the IL12-IFNG pathway with susceptibility to and prognosis of pulmonary tuberculosis in a Chinese population. Eur J Clin Microbiol Infect Dis. 2010;29:1291-5. http://dx.doi.org/10.1007/s10096-010-0985-0 [ Links ]
45. Lugo-Villarino G, Maldonado-López R, Possemato R, Peñaranda C, Glimcher LH. T-bet is required for optimal production of IFN-gamma and antigen-specific T cell activation by dendritic cells. Proc Natl Acad Sci USA.2003;100:7749-54. http://dx.doi.org/10.1073/pnas.1332767100 [ Links ]
46. Xia Z, Liu Q, Berger CT, Keenan BT, Kaliszewska A, Cheney PC, et al. A 17q12 allele is associated with altered NK cell subsets and function. J Immunol. 2012;188:3315-22. http://dx.doi.org/10.4049/jimmunol.1102775 [ Links ]
47. Szabo SJ, Sullivan BM, Stemmann C, Satoskar AR, Sleckman BP, Glimcher LH. Distinct effects of T-bet in TH1 lineage commitment and IFN-gamma production in CD4 and CD8 T cells. Science. 2002;295:338-42. http://dx.doi.org/10.1126/science.1065543 [ Links ]
48. Sullivan BM, Jobe O, Lazarevic V, Vásquez K, Bronson R, Glimcher LH, et al. Increased susceptibility of mice lacking T-bet to infection with Mycobacterium tuberculosis correlates with increased IL-10 and decreased IFN-gamma production. J Immunol. 2005;175:4593-602. [ Links ]
49. Roberts T, Beyers N, Aguirre A, Walzl G. Immunosuppression during active tuberculosis is characterized by decreased interferon-gamma production and CD25 expression with elevated forkhead box P3, transforming growth factor- beta and interleukin-4 mRNA levels. J Infect Dis. 2007;195:870-8. http://dx.doi.org/10.1086/511277 [ Links ]
50. van Crevel R, Karyadi E, Preyers F, Leenders M, Kullberg BJ, Nelwan RH, et al. Increased production of interleukin 4 by CD4+ and CD8+ T cells from patients with tuberculosis is related to the presence of pulmonary cavities. The J Infect Dis. 2000;181:1194-7. http://dx.doi.org/10.1086/315325 [ Links ]
51. Marín ND, París SC, Vélez VM, Rojas CA, Rojas M, García LF. Regulatory T cell frequency and modulation of IFN-gamma and IL-17 in active and latent tuberculosis. Tuberculosis (Edinb).2010;90:252-61. http://dx.doi.org/10.1016/j.tube.2010.05.003 [ Links ]
52. Hohler T, Reuss E, Adams P, Bartsch B, Weigmann B, Worns M, et al. A genetic basis for IFN-gamma production and T-bet expression in humans. J Immunol. 2005;175:5457- 62. [ Links ]
53. Munthe-Kaas MC, Carlsen KH, Haland G, Devulapalli CS, Gervin K, Egeland T, et al. T cell-specific T-box transcription factor haplotype is associated with allergic asthma in children. J Allergy Clin Immunol. 2008;121:51-6. http://dx.doi.org/10.1016/j.jaci.2007.07.068 [ Links ]
54. Suttner K, Rosenstiel P, Depner M, Schedel M, Pinto LA, Ruether A, et al. TBX21 gene variants increase childhood asthma risk in combination with HLX1 variants. J Allergy Clin Immunol. 2009;123:1062-8.e8. http://dx.doi.org/10.1016/j.jaci.2009.02.025 [ Links ]
55. Sasaki Y, Ihara K, Matsuura N, Kohno H, Nagafuchi S, Kuromaru R, et al. Identification of a novel type 1 diabetes susceptibility gene, T-bet. Hum Genet. 2004;115:177-84. http://dx.doi.org/10.1007/s00439-004-1146-2 [ Links ]
56. Gourh P, Agarwal SK, Divecha D, Assassi S, Paz G, Arora-Singh RK, et al. Polymorphisms in TBX21 and STAT4 increase the risk of systemic sclerosis: Evidence of possible gene-gene interaction and alterations in Th1/Th2 cytokines. Arthritis Rheum. 2009;60:3794-806. http://dx.doi.org/10.1002/art.24958 [ Links ]
57. Morita M, Watanabe M, Inoue N, Inaoka C, Akamizu T, Tatsumi KI, et al. Functional polymorphisms in TBX21 and HLX are associated with development and prognosis of Graves' disease. Autoimmunity. 2012;45:129-36. http://dx.doi.org/10.3109/08916934.2011.622013 [ Links ]
58. Yang P, Qiu G, Wang S, Su Z, Chen J, Wang S, et al. The mutations of Th1 cell-specific T-box transcription factor may be associated with a predominant Th2 phenotype in gastric cancers. Int J Immunogenet. 2010;37:111-5. http://dx.doi.org/10.1111/j.1744-313X.2010.00899.x [ Links ]
59. Zhang LH, Li Q, Li P, Zhu ST, Wang J, Yang HL, et al. Association between gastric cancer and -1993 polymorphism of TBX21 gene. World J Gastroenterol. 2012;18:1117-22. http://dx.doi.org/10.3748/wjg.v18.i10.1117 [ Links ]
60. Cao B, Yang H, Ding H, Qi S, Gao L, Cui H, et al. Association analysis of hepatitis virus B infection with haplotypes of the TBX21 gene promoter region in the Chinese population. Clin Chem Lab Med. 2007;45:333-8. http://dx.doi.org/10.1515/CCLM.2007.062 [ Links ]
61. Svensson A, Bergin AM, Lowhagen GB, Tunback P, Bellner L, Padyukov L, et al. A 3'-untranslated region polymorphism in the TBX21 gene encoding T-bet is a risk factor for genital herpes simplex virus type 2 infection in humans. J Gen Virol. 2008;89:2262-8. http://dx.doi.org/10.1099/vir.0.2008/001305-0 [ Links ]
62. Ko FW, Lun SW, Wong CK, Szeto CC, Lam CW, Leung TF, et al. Decreased T-bet expression and changes in chemokine levels in adults with asthma. Clin Exp Immunol. 2007;147:526-32. http://dx.doi.org/10.1111/j.1365-2249.2006.03315.x [ Links ]
63. Chung HT, Kim LH, Park BL, Lee JH, Park HS, Choi BW, et al . Association analysis of novel TBX21 variants with asthma phenotypes. Hum Mutat. 2003;22:257. http://dx.doi.org/10.1002/humu.9169 [ Links ]
64. Ylikoski E, Kinos R, Sirkkanen N, Pykalainen M, Savolainen J, Laitinen LA, et al. Association study of 15 novel single-nucleotide polymorphisms of the T-bet locus among Finnish asthma families. Clin Exp Allergy. 2004;34:1049-55. http://dx.doi.org/10.1111/j.1365-2222.2004.01995.x [ Links ]
