











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkFusarium (Ascomycota, Pezizomycotina, Sordariomycetes, Hypocreomycetidae, Hypocreales, Nectriaceae) is a ubiquitous fungus widely distributed in soil, plants, and organic substrates. Currently, infections by Fusarium are the second most frequent cause by environmental molds and they are associated with high rates of mortality in immunosuppressed patients including those with AIDS and cancer, prolonged use of corticosteroids, and transplanted organs 1-2. The pathogenicity by Fusarium is associated with the production of mycotoxins, proteases, collagenases, and keratinases 3-4.
Pathologies associated with Fusarium in humans include Fusarium keratitis, onychomycosis, endophthalmitis, peritonitis, invasive Fusarium disease, and skin infections. Less frequent infections include osteomyelitis, arthritis, otitis, sinusitis, and brain abscesses 1-2,5. The most common pathology among immunosuppressed patients is invasive or disseminated reaching a 70% to 100% mortality and infection sources are skin and nail lesions 6. This invasive nature, as well as its role in fungal keratitis and its difficult treatment, increase its prevalence in the clinical setting 7-8. The most frequent human pathogens of Fusarium are the species complexes: F. solani (Mart.) Sacc. (FSSC), considered as the most resistant, followed by F. oxysporum Smith & Swingle (FOSC) 2,9. Other species such as F. dimerum Penz. 1882 (FDSC) 10, F. proliferatum (Matsush.) Nirenberg 1976 11, F. incarnatum-equiseti (Desm.) Sacc. 1886 (FIESC) 12,F.sacchari (E. J. Butler & Hafiz Khan) W. Gams 1971 13, F.fujikuroi Nirenberg 1976 (FFSC) 12, F. chlamydosporum Wollenw & Reinking 1925 (FCSC) 12, F. moniliforme J. Sheld 1904 14,15 and F. sporotrichioides Sherb. 1915 15 have also been reported.
The taxonomy of the species complexes in this genus is not a trivial task and there have been many proposals of reclassifying it into multiple genera 16. In the last years, FSSC has been the object of a debate on its reclassification to the genus Neocosmospora17 with arguments in favor of each option. However, the large amount of information about fusariosis shows the importance of keeping the FSSC classification for medical professionals 16. In Colombia, the identification of Fusarium at the species level for epidemiological purposes is not common 18,19, although, in the last years, there have been more studies to characterize clinical isolates at this level 20-23. The lack of this type of characterization has been a serious problem for physicians as proper identification is necessary for an adequate treatment 24. These fungi have variable susceptibility to antifungals and some of them are resistant to all the available treatments 9,12,25. Furthermore, it is necessary to compare the susceptibility of each isolate with a given genotype given the high variability at the intraspecific level 8. This implies characterizing the populations at the highest resolution possible while keeping the simplicity and efficiency required for diagnosis in clinical laboratories in the country.
Fusarium identification traditionally relied on morphological characters; however, these vary at the intraspecific level and are influenced by culture conditions 26. On the other hand, the description of some characters is subjective and the procedure can be lengthy and expensive and it needs trained mycologists 14,26, which prompted the development of molecular techniques for the identification of the genus 26. DNA-based techniques are useful for determining Fusarium taxonomy as they do not have the difficulties arising from the use of morphological characters and, besides, they reveal the genetic diversity 26. Different molecular markers have been proposed for Fusarium genotyping and classification including random species-specific primers and the sequencing of different genes 5,27-30. Currently, the highest resolution marker is the translation elongation factor 1-α (TEF1-α) gene 31 now complemented with multilocus sequence typing (MLST) through the addition of two partial sequences from the two largest DNA-directed RNA polymerase subunits (RPB1 and RPB2) 29,32. Despite these resources, there is still a need for other molecular methods to complement current approaches and provide more information about the genetic diversity of the isolates and their correlation with phenotypical traits 23. Therefore, it is necessary to evaluate molecular markers for the identification of Fusarium species supplying enough information for phylogenetic inference, population genetics, and epidemiology 8.
Our study centered on genotyping clinical isolates of this group of fungi obtained from patients in Medellin, Colombia, to establish the utility of two molecular markers which may provide resolution at the species level and measure the genetic diversity of Fusarium populations. We also evaluated the correlation between the genotypes and relevant phenotypic traits for epidemiological purposes including the isolation tissue and the susceptibility to antifungals. We evaluated two primers according to their gel fingerprints: a mini-satellite probe generated from the M13 bacteriophage and the (GACA)4 simple DNA repeat sequence 5. We compared their performance using sequencing and PCR-based methods already reported by our group 22,23. The genotyping results using these markers were clustered and the classifications compared with other molecular markers including the partial sequencing of the 28S ribosomal subunit (rDNA 28S) 30 and the transcription elongation factor 1-alpha (TEF1-α), widely accepted as reference for this group, as well as the PCR-based identification using 0X31/0X32 primers 27 and Fusofor/Fusorev 28.
Materials and methods
Isolates
We obtained isolates from 101 patients referred by health providers to the Laboratorio de Micología Médica y Experimental at the Corporación para Investigaciones Biológicas (CIB) in Medellin after obtaining their informed consent. Before we took the samples, we gathered the data on their sex, age, place of residence, and previous use of antifungals. The samples were then obtained from hands and feet fingernails, cornea, sinuses, skin, and secretions by non-invasive procedures warranting the chain of custody at the institutions to avoid cross contamination. Samples were cultured in three different media: Sabouraud agar (Thermo Scientific, USA), potato dextrose agar (Thermo Scientific, USA), and mycosel agar (Fisher Scientific, USA) and 10 inoculation points, at room temperature for one to three weeks. Fusarium was considered the etiological agent based on two criteria: 1 the growth of the isolate in the media was observed in more than five of the inoculation points and 2 the same isolate was obtained in at least two of the three media. Once growth was observed, the macroscopic colonies identified as Fusarium sp. were stored in sterile vials 25.
DNA extraction and purification
The isolates were cultured for 10 days in Sabouraud medium (Thermo Scientific, USA) and, subsequently, a portion of the mycelia was macerated with liquid nitrogen in a laminar flow chamber for DNA extraction with the DNeasy Plant Mini-kit (Qiagen) with the following modifications: a vortex of the sample with buffer API for 30 minutes and incubation with RNAse A for another 30 minutes. The DNA was suspended in 80 µl of water for subsequent quantification at 260 nm in a Nanodrop 1000 spectrophotometer (Thermo Scientific, USA).
Molecular genotyping
The molecular genotyping of Fusarium isolates was carried out by PCR using two random primers: a 15-bp minisatellite probe (5’-GAGGGTGGCGGTTCT-3´) from the M13 bacteriophage and a simple DNA repeat sequence (GACA)45. These markers have been widely used to discriminate between related strains of a wide variety of pathogenic microorganisms 5,33-35. As a negative control, we used a PCR mix with no DNA.
The PCR reaction mix of 50 pi included: 5 μl of 10X PCR buffer with 15 mM MgCI2, 3.0 μl dNTP mix (10 μmol/L each), 30 ng of primer, and 2.5 U of Taq DNA polymerase 5. Initial denaturation was at 94 °C for 20 sec followed by 35 cycles of denaturation at 94 °C for 20 sec, annealing at 50 °C for 1 min, amplification at 72 °C for 20 sec, and a final extension at 72 °C for 4 min in an MJ Research thermocycler. The PCR patterns were resolved by electrophoresis on 1.4 % agarose gels in Tris-borate-EDTA (TBE) buffer at 70 V for one hour and a half using the DNA 1 Kb plus DNA ladder as size reference (Fermentas). These were detected by ethidium bromide staining and were visualized on a BioDoc Analyze transilluminator (Biometra). The images were digitally recorded for further analysis.
Fingerprint analysis
The PCR patterns obtained by the amplification of DNA fragments of different sizes for each isolate were termed as M13 and (GACA)4 genotypes. These fingerprints were analyzed by GelCompar II (Applied Maths NV, Kortrijk, Belgium) using the Pearson correlation coefficient, which considers each band pattern as a densitometric curve. Two dendrograms were built with the results of M13 and (GACA)4 using the UPGMA algorithm. These dendrograms and pairwise matrices calculated using Euclidean distance were then exported for posterior analysis.
Genotypes were clustered by their degree of similarity in each dendrogram. The isolates with identical band patterns were established as the same genotype and assigned the same number; those differing in one or two bands were considered closely related genotypes (with the same number but differentiated by a lowercase letter), and differing in three or more bands, as different genotypes (with different numbers). The identity of most of the strains (e.g. FOSC or FSSC), was determined in a previous work by TEF1-α partial sequence 23. Dendrogram annotation was done using the clustering classification, the isolation tissue, and the molecular identification at the species level.
Hunter Gaston discriminatory index (HGDI) calculation
Hunter Gaston discriminatory index (HGDI) was calculated as proposed by Hunter, et al. and Sola, et al. 36-37:
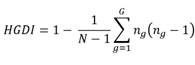
where N is the total number of strains in the population, is the total number of distinct patterns given by each genotyping method, and is the number of strains corresponding to the pattern . We took the clustering classifications obtained by M13 and (GACA)4 markers at the different levels of resolution given by the clusters (low resolution) and the genotypes (high resolution). For the sequencing methods (rDNA 28S and TEF1-α), we calculated the indexes grouping by the different haplotypes obtained using the Arlequín 3.5 software 38 as the haplotypic diversity. For the PCR-based identification, we grouped the results from the two pairs of primers 0X31/ 0X32 and Fusofor/Fusorev considering that they only provide a binary classification (i.e., presence/absence of amplification).
Antifungal susceptibility tests
We evaluated a group of 44 isolates for their susceptibility to the in vitro activity of amphotericin b, itraconazole, and voriconazole using the disk diffusion test according to the CLSI M38-A methods as previously reported 25. Isolates were grown on PDA agar for a week at room temperature. Mycelia were resuspended with distilled water Into a 10 ml tube and sedimented for 20 min. The supernatant was collected and normalized to a concentration of 106 UFC/ml; 200 pi of this inoculum was added to RPMI agar plates with MOPS buffer (AES Laboratories, Paris, France) and dried for 15- 30 minutes. We performed an E-test following the manufacturer’s instructions (Etest®-AB Biomérieux). We placed on the agar plate two strips of antifungal with a concentration gradient in the range of 256 to 0.016 pg/ml opposite from the inoculum and incubated it at 28 ºC for 48 hours. Susceptibility values were recorded as the lowest antifungal concentration inhibiting the fungal growth.
Clustering enrichment analysis
To test the clustering classifications obtained with the M13 and (GACA)4 markers and their associations with epidemiological variables we used a hypergeometric test. We evaluated the isolation tissue, the sequencing-based identification at the species level, the antimycotic sensibility in µg/ml, and the antimycotic phenotype (resistant, sensible-dose-dependent, and sensible) as reported in a previous characterization of these isolates 25. We controlled for false discovery rate adjusting by the Benjamini-Hochberg procedure 39. We also plotted some of these variable distributions using ggplot2 40.
Analysis of molecular variance (AMOVA) and correlation reference classification
The distance matrix obtained with the M13 marker was used in an AMOVA 41 analysis in search of population structures given the clustering and then compared to the TEF1-α and rDNA 28S haplotype reference classification. The correlation between the distance matrices computed with these methods was compared using a Mantel test 42.
The R code used to perform all the statistical analyses and cluster association figures can be found at https://github.com/welasqz/Fusarium_Genotyping
Results
Fingerprinting clusters from M13 and (GACA) 4 showed differences in their resolution.
A total of 32 and 58 genotypes were revealed by the markers (GACA)4 and M13, respectively (figure 1).The M13 marker showed more power of resolution, and its band patterns were more complex and depicted a higher number of bands. The most frequent genotype by M13 was number 58, with 16 isolates, all belonging to FOSC. The second most frequent genotype was 51, with 6 isolates (of FOSC) followed by genotype 43 with 5 isolates (of FOSC), and genotypes 5 and 24 of FSSC and 52 of FOSC, with 4 isolates each one. Genotypes 22, 23, and 49 had 3 isolates each and the genotypes 38, 54, 56, and 57 had 2 isolates each. Unique genotypes (with only one isolate associated) were obtained for 48 (82.7%) isolates.
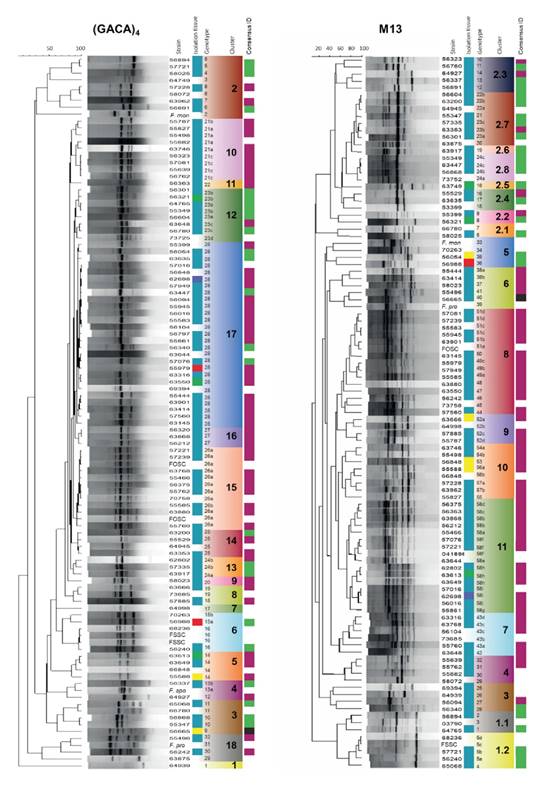
Figure 1 (GACA)4 and M13 dendrograms obtained by the UPGMA method using the Pearson correlation. The first column of each dendrogram correspond to the strain number. Several refence strains were included and abbreviated as FSSC (F.solani (Mart.) Sacc.), FOSC (F. oxysporum Smith & Swingle), F.mon (F. moniliforme J. Sheld. 1904), F.pro (F. proliferatum (Matsush.) Nirenberg 1976), and F.spo (F. sporotrichioides Sherb. 1915). The second column refers to the isolation tissue (blue: toenails, green: fingernails, purple: abdomen, red: cornea, and yellow: skin). The third column shows the genotypes classification with their number and clusters (differentiated by color and number). The last column refers to the consensussequencing identification (green: FSSC, purple: FOSC, and black: Fusarium incarnatum).
The most frequent genotype revealed by the (GACA)4 marker was number 28 assigned to 27 isolates followed by genotype 26 with 12 isolates; genotype 21 with 9; 23 with 8,14,16, and 25, each with 4; 5, 24, and 27 with 3, and genotypes 8,10,11,13,15, and 19 with 2 isolates each. Unique genotypes by (GACA)4 were obtained for 16 isolates (50%). Compared to M13, the majority of the (GACA)4 clusters had a mixture of different Fusarium species.
M13 and (GACA) 4 showed the highest discriminatory power as compared to other genotyping methods for Fusarium
HGDI values were computed for the different resolution levels obtained from the M13 and (GACA)4 markers, the sequencing-based genotyping using rDNA 28S and TEF1-α, and the PCR-based methods. Reference values 37 indicate a method is highly discriminatory if h>0.6, moderately discriminatory if 0.3<h<0.6, and poorly discriminatory if h<0.3. The highest discriminatory power was obtained by the genotype and high-resolution cluster level of M13 with 0.9970 and 0.9269, respectively. The (GACA)4 genotype and cluster classifications followed with 0.9156 and 0.8932, and then, M13 low-resolution clusters with 0.8693. TEF1-α had a value of 0.8305 followed by rDNA 28S with 0.7778, and, finally, the lowest value reported by PCR-based methods with 0.5235.
The M13 marker was associated with epidemiological variables.
We plotted the clustering classifications computed with the M13 and (GACA)4 markers associated with several epidemiological variables as shown in figure 2. We looked for statistical associations among the levels of these variables and each cluster using a hypergeometric test. The M13 marker showed stronger associations with all the variables: 1) with the molecular identification given by TEF1-α and rDNA 28S, with clusters 1-5 linked with FSSC, with cluster 6 as a transition group (FSSC, FOSC, and Fusarium spp), and with clusters 7-11 containing strains identified as FOSC; 2) with isolation tissues, especially cluster 5 which grouped strains from the cornea and skin tissues, and cluster 11 containing all abdomen isolates; 3) with the voriconazole response showing resistant phenotypes associated with clusters 1-5 (FSSC) and susceptible-dose-dependent phenotypes in the 6-11 clusters (FOSC). The M13 marker was then taken forward as a useful genotyping method for Fusarium.
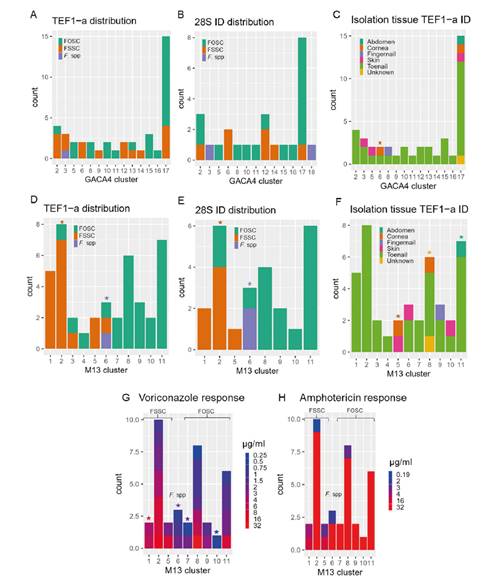
Figure 2 (GACA)4 and M13 association with several epidemiological variables. Cluster associations with the sequencing-based identification of the isolates for (GACA)4 (A, B) and M13 (D, E). Associations with the isolation tissue (C, F) for each marker clusters and associations between M13 and the response to antifungal compounds (G, H). M13 clusters 1 to 5 are enriched with isolates classified as FSSC, cluster 6 with Fusarium spp, and clusters 7 to 11 with FOSC. Significant hypergeometric values (p-adjusted <0.001) reported for category-mixed clusters indicating the significant category with the same color as in the histogram.
M13 measured population structure and had a high correlation with the reference Fusarium identification method.
After exploring the M13 marker clustering when characterizing Fusarium populations from clinical samples, we assessed the genetic variability of the marker and its correlation with the genotyping obtained from sequencing- based methods. Haplotypes from TEF1-α and rDNA 28S were calculated for the FSSC and FOSC populations and inputted with the M13 Euclidean distance matrix to an AMOVA test. The highest haplotype diversity was found for FSSC with values of 0.8097 for rDNA 28S and 0.8917 for TEF1-α. FOSC had values of 0.5385 and 0.5435, respectively. Concatenated haplotypes were used as input data for the AMOVA and we found a population structure using the M13 fingerprint data with a p-value of 0.0099 and a φ statistic of 0.2509.
To confirm the correlation between the two methods, we performed a Mantel test on the distance matrices and found a strong correlation between them (p-value=0.02). These analyses established M13 as a discriminative marker for Fusarium molecular characterization maintaining the correlation and structure of the reference molecular identification and a higher discriminatory power.
Discussion
Fusarium fungi are opportunistic pathogens causative agents of human mycoses; in agriculture, they cause a decrease in crop yields and economic losses. They are commonly characterized as a group of saprophytes in many environments and as characteristic inhabitants of the soil 43. Studies have called attention to the increasing cases of onychomycosis by non-dermatophyte fungi like Fusarium around the world 7; hypotheses explaining such an increase include better identification of these organisms as onychomycosis primary agents and the recent Fusarium spp. detection as emerging pathogens in growing immunocompromised populations 7.
Fusarium has great genetic variability and is difficult to classify 26,44. One of the major problems is the lack of diagnostic morphological characters, which makes the correct species identification and taxonomic classification very complex. However, the available molecular tools can improve the resolution at species level identification 26,44. The phylogenetic species recognition of Fusarium clinical isolates has been proposed based on the partial sequences of the TEF1-α, RPB1, and RPB2 genes 32. This method, also called multilocus sequence typing (MLST), is currently recognized as the best strategy for genetic population studies and to identify Fusarium species complexes 8,32. Our HGDI calculations indicate that M13 has a similar discrimination power to the MLST method documented for Fusarium, with 0.9970 for M13 vs 0.991 for MSLT 32. Besides, when a huge geographic diversity is involved, a large number of isolates of Fusarium from different environments, hosts, and tissues are available for biology, clinical, and evolutionary studies. Therefore, a simple, fast, cheap, and effective methodology is always required to select the most representative isolates for further studies. Here we confirmed that the M13 marker meets these qualifications.
More than 35.000 strains isolated from various substrates around the world are accessioned in the Fusarium Research Center (FRC) and the USDA- ARS National Center for Agricultural Utilization Research (NCAUR) Culture Collection, which makes this genus the best-preserved fungal group 45. Using this rich strain resource, extensive molecular phylogenetic studies have been conducted resulting in data covering most agriculturally and/or medically important species complexes. Despite these advances, a significant amount of diversity has yet to be explored and some species complexes are quite poorly characterized phylogenetically 45.
Here we were able to find a huge diversity of Fusarium genotypes (58 out of 101 isolates) using the M13 marker and the genotyping was consistent with the identification by TEF1-α for most of the isolates (figure 1) with a significant correlation between the two markers. We found associations between the clusters and multiple epidemiological variables (figure 2) such as the isolation tissue and the response to antimycotics, which supports the population structure given by the marker using AMOVA and demonstrates the utility of the marker in assessing the genetic variability of the isolates at the population level.
The most heterogeneous and basal genotypes were those belonging to FSSC, in agreement with the haplotypic diversity measurements from sequencing-based methods and other work 46,47. Then we found that the M13 cluster 6 is a transition group with multiple species followed by 5 clusters comprising the FOSC isolates. Noteworthy, most of the isolates we studied were identified as FOSC (53/77; 69%) and only 31% (24/77) as FSSC, which contrasts with other reports from Tropical areas (even with another one from Colombia) where FSSC was found to be the main human pathogen group of Fusarium12 probably due to the fact that FOSC is a prevalent plant pathogen and some genotypes rapidly disseminate and adapt in our geography. Noteworthy, many of the FOSC isolates had identical or closely related genotypes (e.g., clusters 7-11 in figure 1), and lower haplotypic diversity.
Besides the species complex identification, we showed associations between the M13 classifications and several epidemiological variables including the isolation tissue and the antifungal susceptibility. The associations with the isolation tissues indicated that the M13 cluster 5 was related to more unique tissues, such as cornea and skin, and that the genotypes isolated from these were closer to each other than the rest. In contrast, the most abundant tissues, such as toenails, had a broad distribution and diversity. When evaluating the response to antifungals, only the voriconazole presented a wide range of susceptibilities to test the associations with the classifications. We found that the response to voriconazole was associated with the species and, therefore, with the M13 classification. These results suggest that further research to develop new antifungal molecules is required for this group of fungi.
The M13 probe was established as a marker in 1987 to detect hypervariable DNA minisatellite regions in humans and animals 48. It has been used for studying the genetic epidemiology of pathogenic fungi 49 and was the first reported for typing isolates responsible for fungal keratitis belonging to FOSC and FSSC 50. Undoubtedly, considering the number of clinical isolates we characterized in this study and the low cost of the PCR- based methodology, the M13 marker is recommended to detect Fusarium genetic variability. We propose its use to choose the most diverse and/or frequent clinical isolates that should be analyzed using the MLST scheme and different omics such as genome sequencing. A local-blast analysis involving the 15-bp M13 sequence and the four Fusarium cured-genomes reported in the Fusarium Comparative Genomics Platform (FCGP) could reveal the extent of the genome coverage of this marker.

