










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.10 no.2 Bogotá Dec. 2005
CICLOS DE POSTURA ANUAL ex situ DE Rhinoclemmys melanosterna, R. diademata Y DE SUS HÍBRIDOS (Reptilia: Testudines: Emydidae: Batagurinae)
Ex situ Annual Egg—Laying Cycles of Rhinoclemmys melanosterna, R. diademata and their Hybrids(Reptilia: Testudines: Emydidae: Batagurinae)
JAIME RAMÍREZ-PERILLA
Departamento de Biología (Unidad de Biología Animal), Facultad de Ciencias, Universidad Nacional de Colombia. Sede Bogotá.
Presentado febrero 15 de 2005, aceptado junio 13 de 2005, correcciones agosto 22 de 2005.
RESUMEN
Registros históricos de postura ex situ de huevos de tortugas del género Rhinoclemmys en la Estación de Biología Tropical Roberto Franco (EBTRF), correspondientes a los años 19911999, fueron analizados en relación con factores climáticos locales. Rhinoclemmys melanosterna, Rhinoclemmys diademata y sus híbridos presentan ciclos de postura continuos durante el año con máxima frecuencia durante la estación lluviosa. La época de postura máxima durante un ciclo anual del híbrido R. melanosterna x R. diademata es más temprana que la de sus progenitores de origen. No parece existir relación directa entre la oferta de alimento ad libitum y los ciclos de postura de Rhinoclemmys sp.
Palabras claves: Reptilia, Testudines, Rhinoclemmys, reproducción, ciclos reproductivos.
ABSTRACT
R. melanosterna, R. diademata and their hybrids (R. melanosterna x R. diademata) present annual continuous egglaying cycles with maximum frequency during the rainy season. The period of maximal annual egglaying for hybrid R. melanosterna x R. diademata is earlier than its parents. Aparently there is not direct relation between the ad libitum food offer and Rhinoclemmys sp egglaying cycles.
Key words: Reptilia, Testudines, Rhinoclemmys, reproduction, reproductive cycles.
INTRODUCCIÓN
MATERIALES Y MÉTODOS
LUGAR DE LA INVESTIGACIÓN
Estación de Biología Tropical Roberto Franco, EBTRF (Facultad de Ciencias, Universidad Nacional de Colombia) en la ciudad de Villavicencio (4°09’12’’ latitud norte y 73°38’06’’ longitud oeste), Meta, Colombia.
ANIMALES Y MANEJO ALIMENTARIO
La colonia de animales objeto de este estudio fue implantada hace 30 años por Federico Medem en la EBTRF bajo condiciones de cautividad con manejo alimentario oportunista ad libitum. A partir de 1990 se implementó un régimen alimentario diario, a discreción según disponibilidad, basado en concentrados peletizados comerciales para gallinas ponedoras, complementado con frutas y verduras. Se han mantenido separadas las líneas parentales, Fo, tanto de R. melanosterna como de R. diademata,y generaciones sucesivas de R. melanosterna x R. diademata se han venido reproduciendo exitosamente en cautividad.
CICLOS DE POSTURA EN RELACIÓN CON VARIABLES CLIMÁTICAS
A partir de datos históricos de postura de R. melanosterna, R. diademata y sus híbridos (R. melanosterna x R. diademata), registrados diariamente con excepción de días feriados, se construyó una base de datos de porcentaje de ovipostura/mes, entre los años 1991 y 1999, que fue cotejada con información meteorológica mensual correspondiente al mismo período, suministradas por el Servicio del Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM) de la estación del aeropuerto “Vanguardia” de la ciudad de Villavicencio. Las variables climáticas promedio mes seleccionadas para el período 1991-1999 fueron temperatura (°C), precipitación (mm), humedad relativa (%) y brillo solar (horas/luz). Se hizo un análisis estadístico descriptivo de los ciclos de postura anual con base en promedios/mes de frecuencias relativas de postura, durante el lapso indicado y, mediante la técnica de correlación de Pearson se determinó si existía algún grado de asociación entre la ovipostura y cada una de las variables climáticas seleccionadas. Igualmente se determinó si existía correlación entre las posturas/mes de la especies estudiadas.
RESULTADOS Y DISCUSIÓN
CONDICIONES CLIMÁTICAS
Los indicadores climáticos promedio en el aeropuerto Vanguardia (IDEAM) de la ciudad de Villavicencio correspondientes al área de ubicación geográfica de la EBTRF, entre los años 1991 y 1999 (3.287 días, 108 meses) son los siguientes: temperatura promedio diurna de 25,6 °C ± 0,99, (máxima 29,2 °C y mínima 23,5 °C); pluviosidad promedio mes de 389,7 mm ± 209,43, (máximo 877,1 mm y mínimo 13,2 mm); humedad relativa promedio de 77,6% ± 5,88, (máximo 87% y mínimo 55%); horas totales promedio luz mes de 134,96 ± 29,96, (máximo 222,2 y mínimo 76,1 horas luz). Como las variaciones de temperatura señaladas son promedio de valores horarios medios diurnos/mes, éstos no reflejan las fluctuaciones reales de la temperatura cuando se confrontan valores mínimos absolutos de temperatura en la noche, antes del amanecer (17 °C) frente a los máximos absolutos diurnos (36,8 °C) de un día soleado. El promedio de temperatura diurna/mes a lo largo del año no varía significativamente entre meses, sean estos de baja o de alta precipitación.
En Villavicencio el régimen pluviométrico es de tipo unimodal (Fig. 1A) con un período de baja precipitación entre los meses de diciembre y marzo (rango de 77,53 a 265,10 mm promedio de precipitación/mes) y picos máximos en los meses de abril, mayo y junio (567 mm, 655 mm y 541 mm respectivamente) seguido de una leve disminución de las lluvias en el mes de agosto (412 mm) que se incrementa en el segundo semestre en los meses de septiembre a noviembre (492,6 mm; 491,6 mm; 405,7 mm respectivamente) sin llegar a los mismos niveles de intensidad del primer semestre. Los análisis de correlación múltiple (índice de Pearson) entre las variables climáticas señaladas indican asociaciones esperadas (Fig. 1B) de tal forma que, en general, existen tendencias de correlación negativa entre precipitación/mes vs. temperatura/mes (p = 0,57); entre el brillo solar (horas luz/mes) vs. precipitación/mes (p =0,26) y entre el brillo solar/mes vs. humedad relativa/mes (p = 0,40). Por el contrario, tendencias de correlación entre variables de precipitación vs. humedad relativa (p =0,72) o entre el brillo solar vs. temperatura (p = 0,42) son positivas. El comportamiento climático es propio de la región ecuatorial donde las variaciones dependen del régimen modal de las lluvias, a diferencia de las zonas templadas donde la estacionalidad es termodependiente.
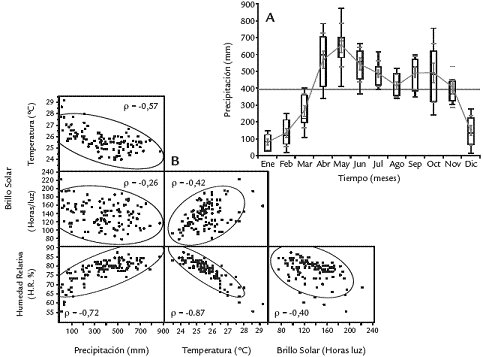
Figura 1. Régimen pluviométrico (mm) promedio mes. A. Entre los años 1991-1999 en la ciudad de Villavicencio, Colombia. B. Índices de correlación múltiple (coeficientes de Pearson) entre variables climáticas.
CICLOS DE POSTURA
Durante los 3.287 días monitoreados, R. diademata presentó 264 eventos de postura en 170 días diferentes (promedio de 1,5 huevos/día), de los cuales, en dos fechas diferentes, 13 de julio de 1992 y 19 de diciembre de 1995 hubo posturas máximas de seis huevos. La postura total para R. melanosterna fue de 967 huevos en 383 días (promedio de 2,5 huevos/día), con un evento de 11 huevos puestos el día 6 de mayo de 1996. Por su parte R. diademata x R. melanosterna registró 237 días de postura con un total de 515 huevos (2,2 huevos/día), con máxima postura/día de siete huevos en agosto 11 de 1991, marzo 31 de 1993. Es de suponer que la concentración de posturas en un solo día son evidencia de eventos independientes de posturas individuales simultáneas, o de un número de individuos proporcional al número máximo de huevos que pueden poner en un único evento. Para el caso de R. melanosterna se ha determinado posturas/individuo de uno a dos huevos, rara vez tres (Castaño Mora y Medem, 1983). No se sabe con qué intervalo de tiempo sucede una postura después de otra por individuo, tampoco es fácil discernir qué mecanismos ecofisiológicos regulan la maduración de folículos, su vitelogénesis, ovulación y eventos subsecuentes que conllevan a la postura de huevos fecundados o sin fecundar. En las condiciones de la EBTRF, R. melanosterna, R. diademata e híbridos ponen huevos todo el año (Fig. 2). En general, R. melanosterna concentra su postura durante el segundo semestre del año igual que R. diademata (81,47% y 70,93% de la postura total/año, Figs. 2A y 2B, respectivamente), mientras los híbridoslo hacen entre el segundo y tercer trimestre del año (80,72% de la postura, Fig. 2C).

Las frecuencias relativas promedio de posturas son mínimas en R. melanosterna entre los meses de enero y abril (Fig. 2A) con porcentajes que fluctúan entre 2,34% y 3,34%; a partir de mayo (6,91%) la postura se incrementa sucesivamente mes a mes hasta un máximo de 19,81% en el mes de octubre; noviembre y diciembre son progresivamente declinantes (10,78% y 7,65%, respectivamente). Un comportamiento similar se presenta en R. diademata (Fig. 2B). Los híbridos, R. melanosterna x R. diademata, difieren un poco del patrón descrito. En éstos, (Fig. 2C), la postura se anticipa ligeramente comenzando en marzo (7,5% de posturas) con frecuencias incrementando sucesivamente hasta el mes de agosto (19,39%) y que comienzan a declinar en septiembre (7,43%); los meses de postura mínima son entre octubre (1,77%) y febrero (3,28%) del año siguiente (las frecuencias mínima y máxima en este período son 1,19% y 3,85%).
Es evidente que los ciclos de postura de R. melanosterna y R. diademata son similares a través del año (p = 0,902; correlación de Pearson) y no existe ningún grado de asociación temporal entre los ciclos de postura de híbridos, R. melanosterna x R. diademata, con R. melanosterna (p = 0,118), ni con R. diademata (p = 0,013). Desde el punto de vista de la heredabilidad genética esperada no es razonable que los híbridos tengan una temporada de postura diferente a la de sus especies de origen si todas están expuestas a condiciones climáticas y manejo alimentario similares. La información de postura descrita para R. melanosterna es coherente tanto con los registros de postura en los meses de diciembre a enero hechos por Dahl y Medem (1964) en el bajo Sinú, noroeste de Colombia; como por los publicados por Castaño Mora y Medem (1983) quienes, sobre 299 huevos puestos en el lapso de 1965 a 1983 en la EBTRF, encontraron que las frecuencias de postura fueron máximas entre los meses junio a septiembre.
CICLOS DE POSTURA Y FACTORES CLIMÁTICOS
Un análisis de correlación múltiple (coeficiente de Pearson) entre frecuencias relativas de postura de Rhinoclemmys sp. en función de variables climáticas/mes, bajo las condiciones de manejo en la EBTRF, revela una correlación positiva mayor con los fenómenos climáticos asociados a la lluvia.
R. diademata, R. melanosterna e híbridos (R. diademata x R. melanosterna) presentan índices de correlación relativamente significantes entre postura vs. precipitación/mes (p = 0,60; p = 0,95 y p = 0,87, respectivamente) comparados con coeficientes de correlación menores en función de la temperatura (p = 0,37; p = 0,60 y p = 0,42, respectivamente). Excepcionalmente, el híbrido R. diademata x R. melanosterna muestra una alta correlación (p = 0,9) entre sus posturas y el número de días lluviosos/mes, que es coherente con la mayor pluviosidad en el primer semestre del año cuando se concentra la postura de las especies híbridas (Fig. 2C). Es contradictorio la correlación muy positiva entre posturas vs. horas luz solar (p = 0,84).
DISCUSIÓN
Las especies del género Rhinoclemmys y sus híbridos, aquí analizados, se comportan conforme a la definición de ciclos reproductivos continuos, propios de organismos de hábitat tropicales, que se diferencian de los ciclos reproductivos denominados asociados o disociados, propios de organismos de lugares de latitudes altas con estacionalidad climática (Pough et al., 1998). Una situación evidente en R. melanosterna, R. diademata y sus híbridos es que sus ciclos de postura, bajo las condiciones de la EBTRF, no son directamente dependientes de la disponibilidad de alimento ya que las frecuencias de ovipostura disminuyen dramáticamente en verano a pesar que el suministro de alimento se mantiene igualmente disponible durante todo el año. Tampoco son explicables las diferencias entre la época de mayor postura de los híbridos (R. melanosterna x R. diademata) y la época de postura de la especie R. melanosterna o de R. diademata. melanosterna, propia de la costa pacífica y del norte de Colombia (Medem, 1958; Mittermeier et al., 1980) y R. diademata de la cuenca del río Catatumbo en Colombia y del noroeste de Venezuela (Ernst y Barbour, 1989) con un mismo origen filogenético, se diferenciaron probablemente como consecuencia de su aislamiento reproductivo mediado por la cordillera oriental que se extiende de sur a norte en Colombia hasta Venezuela. El hábitat de una y otra especie son similares en cuanto a que son bosques tropicales húmedos, y es de suponer que sus ciclos de postura a lo largo del año sean similares en vida silvestre, dado que están expuestas al mismo régimen pluviométrico, porque al parecer ocupan el mismo rango de la franja latitudinal influida por los ciclos anuales de confluencia de nubosidad intertropical. Nicéforo (1958) registró en el mes de enero de 1958, en la cuenca del río Zulia, Catatumbo, a un ejemplar de Geomyda punctularia diademata (sinónimo de R. diademata) que tenía dos huevos con cáscara dura y dos óvulos, mientras Medem (1962a) halló 12 óvulos pequeños en un ejemplar muerto de Geoclemmys melanosterna (sinónimo de R. melanosterna) lo cual podría significar un mecanismo fisiológico de poliovulación simultánea seguido de maduración de óvulos, diferidos temporalmente unos de otros, de tal manera que posteriormente pueden suceder posturas de uno, dos o tres huevos simultáneamente con frecuencias distintas a lo largo de un ciclo anual.
El surgimiento de los híbridos en condiciones de hábitat artificiales propios de la cautividad, y con dependencia externa de suministro de alimento, que bien pueden ser causa del desarrollo de estrategias reproductivas propias un tanto diferentes, como mecanismo adaptativo a su hábitat y nichos particulares. Es bien conocido que factores externos como el clima y la disponibilidad de recursos en el medio ambiente o mecanismos endógenos de carácter fisiológico (metabólicos y neuroendocrinos), y sus interacciones, regulan la madurez gonadal (óvulos y espermatozoides) y contribuyen a modelar patrones de conducta complejos asociados a la reproducción (Crews, 1978; Alcock, 1998) bajo condiciones ecológicas distintas. En animales poiquilotermos, que es el caso de las tortugas, cuando la temperatura ambiente se incrementa, el metabolismo también lo hace, de tal forma que la energía consumida en el alimento se gasta más en mantenimiento, como gasto metabólico, presentándose un probable déficit para el crecimiento, y con mayor razón para la reproducción como ha sido demostrado en estudios de peces (Brett, 1971) y anfibios (Lillywhite et al., 1973). Esta podría ser una razón por la que especies del género Rhinoclemmys disminuye su postura en época de verano en la EBTRF. También es de esperar que con la baja de temperatura ambiente la apetencia disminuya, tal como se observó en un estudio sobre consumo y apetencia de juveniles de R. melanosterna, que redujeron sus tasas de crecimiento debido a un menor consumo cuando la temperatura ambiental era menor, así estas relaciones no fueran estadísticamente evidentes (Rangel, 2002). Aunque el ciclo de postura ex situ de Rhinoclemmys sp. es continuo, con meses de mayor porcentaje de postura durante las temporadas de lluvias y bajas posturas en verano, a pesar del suministro permanente de alimento de buena calidad, es de suponer que la falta de correspondencia entre la disponibilidad de alimento y las frecuencias de postura se debe a altas variaciones en las tasas metabólicas propias de animales poiquilotermos (Nagy, 1982; 1983). Sin embargo, dado el origen tropical de Rhinoclemmys es posible que estas especies se asemejen más a otras tortugas con modelos metabólicos en los que a mayor disponibilidad de alimento sea posible incrementar el esfuerzo reproductivo (Swingland, 1977), asumiendo condiciones ambientales estables dentro de límites de confort térmico. Esta sería una situación muy diferente a lo evidenciado en tortugas del desierto, Gopherus agassizzii, en la que la postura de huevos es independiente de la lluvia y de la disponibilidad de alimento debido a que las hembras almacenan energía antes del invierno y usan estas reservas para asegurar las posturas en primavera (Henen, 2002). La subfamilia Batugurinae es la más primitiva del grupo Emydidae y se caracteriza por la producción constante de posturas con muy pocos huevos extremadamente grandes que garantiza una mayor sobrevivencia de los recién nacidos (Pritchard y Trebbau, 1984). Es de suponer que bajo condiciones de humedad relativa alta, disponibilidad permanente de alimento y temperaturas constantes propias del promedio de la franja tropical las especies del género Rhinoclemmys incrementen el esfuerzo reproductivo (número de huevos por postura) con mayor frecuencia de posturas todo el año, dentro de los límites genéticofisiológicos máximos posibles. Situaciones de esta naturaleza deberían ser objeto de experimentación, sobre todo con propósitos de conservación dado el estatus de especie vulnerable de R. diademata y de casi amenazada de R. melanosterna (Castaño Mora y Medem, 2002a, 2002b).
AGRADECIMIENTOS
A la División de Investigación de la Universidad Nacional de Colombia, sede Bogotá. A César Urueña y a Willington Martínez asistentes de la Estación de Biología Tropical Roberto Franco, por su colaboración en la recuperación, depuración y sistematización de la base de datos.
BIBLIOGRAFÍA
ALCOCK J. Animal Behavior: An Evolutionary Approach. Sunderland, Massachusetts: Sinauer Associates, Inc. Publishers; 1998. [ Links ]
BRETT JR. Energetic Reponse of Salmon to Temperature. A Study of Some Thermal Relations in the Physiology and Freshwater Ecology of Sockeye Salmon (Oncorhynchus nerka). Am Zool. 1971;11:99-114. [ Links ]
CASTAÑO MORA O, MEDEM F. Datos preliminares sobre la reproducción de Rhinoclemmys melanosterna Gray (Reptilia: Quelonia: Emydidae). LOZANIA (Acta Zoológica Colombiana) 1983;47:1-6 [ Links ]
CASTAÑO MORA O. Rhinoclemmys diademata. En: Castaño Mora OV. Editor. Libro rojo de reptiles de Colombia. Libros rojos de especies amenazadas de Colombia. Bogotá, Colombia: Instituto de Ciencias Naturales Universidad Nacional de Colombia, Ministerio del Medio Ambiente, Conservación Internacional Colombia; 2002a. [ Links ]
CASTAÑO MORA O. Rhinoclemmys melanosterna. En: Castaño Mora OV, Editor. Libro rojo de reptiles de Colombia. Libros rojos de especies amenazadas de Colombia. Bogotá, Colombia: Instituto de Ciencias Naturales Universidad Nacional de Colombia, Ministerio del Medio Ambiente, Conservación Internacional Colombia; 2002b. [ Links ]
CREWS D. Integration of Internal and External Stimuli in the Regulation of Lizard Reproduction. En: Greenberg N, MacLean P, Editores. Behavior and Neurology of Lizards. Rockville, Maryland, USA: National Institute of Mental Health; 1978. p. 149-172. [ Links ]
DAHL G, MEDEM F. Informe sobre la fauna acuática del río Sinú. Corp Auton Reg Valles Magdalena y Sinú (CMV); 1964. p. 41-44. [ Links ]
ERNST CH, BARBOUR RW. Turtles of the World. Washington, USA: Smithsonian Institution Press; 1989. [ Links ]
HENEN BT. Reproductive Effort and Reproductive Nutrition of Female Desert Tortoises: Essential Field Methods. Integr Comp Biol. 2002;42:43-50. [ Links ]
LILLYWHITE HB, LICHT P, CHELGREN P. The Role of Behavioral Thermorregulation in the Growth Energetics of the Toad, Bufo Boreas. Ecology. 1973;4:375-383. [ Links ]
MEDEM F. Informe sobre reptiles colombianos (II). El conocimiento actual sobre la distribución de Testudinata en Colombia. Bol Mus Cienc Nat Caracas. 1958;23(14):13-45. [ Links ]
MEDEM F. La distribución geográfica y ecología de los Crocodylia y Testudinata en el departamento de Chocó. Rev Acad Colomb Cienc Exact Fis Nat. 1962a;(11):279-303. [ Links ]
MEDEM F. Informe final sobre la comisión realizada a los ríos Atrato, San Juan y Baudó. Consejo Nacional de Política Económica y Planeación. Documentos Desarrollo Chocó. Plan Fomento Regional 1959-1969. Cali: Impr. Edit. Norma; 1962b. p. 684-693. [ Links ]
MITTERMEIER RA, MEDEM F, RHODIN AGJ. Vernacular Names of South American Turtles Society for the Study of Amphibians and Reptiles. Herpetological. 1980;Circular 9:1-45. [ Links ]
NAGY KA. Energy Requirements of Free-Living Iiguanid Lizards. In: Burggghardt GM and Rand AS, Editores. Iguanas of the World: Their Behavior, Ecology and Conservation. New Yersey, USA: Noyes publications, Park Ride; 1982. p. 49-59. [ Links ]
NAGY KA. Ecological Energetics. En: Huey RB, et al. Editors. Lizard Ecology: Studies of a Model Organism. Cambridge, Massachussets: Harvard University Press; 1983. [ Links ]
NICÉFORO MARÍA HNO. Contribución al estudio de los testudineos de Colombia. Familia Emydidae, Geomyda punctularia diademata Mertens. San José, Medellín. Bol Cult Col. 1958;23:31-37. [ Links ]
PAOLILLO A. Description of a New Subspecies of the Turtle Rhinoclemmys Punctularia (Daudin) (Testudines:Emydidae) From Southern Venezuela. Amphibia-Reptilia. 1985;6:293-305. [ Links ]
POUGH FH, ANDREWS RM, CADLE JE, CRUMP ML, SAVITZKY AH, WELLS KD. Herpetology. New Jersey, USA: Prentice-Hall, Inc.; 1998. [ Links ]
PRITCHARD PC. Encyclopedia of Turtles. New Jersey: T.F.H. Publ., Inc., Neptune; 1979. [ Links ]
PRITCHARD PC, TREBBAU P. The Turtles of Venezuela. Oxford, OH: Society for the Study of Amphibians and Reptiles; 1984. [ Links ]
RANGEL MENDOZA A. Consumo y apetencia en juveniles d la tortuga palmera (Rhinoclemmys melanosterna) mantenidos en cautividad bajo diferentes dietas y frecuencias de alimentación [trabajo de pregrado]. Bogotá, Colombia: Universidad Nacional de Colombia, Departamento de Biología; 2002. [ Links ]
RAMÍREZ-PERILLA J. Técnicas de recolección y control de calidad de huevos de tortugas y cocodrilos. Apéndice 1: ciclos de postura y épocas de cosecha. Zoodivulgación. (Estación de Biología Tropical Roberto Franco). 1999;1:31-32. [ Links ]
SWINGLAND IR. Reproductive Effort and Life History Strategy of the Aldabran Giant Tortoise. Nature. 1977;269:402-404. [ Links ]

