










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.11 no.1 Bogotá June 2006
CRECIMIENTO Y DESARROLLO DE Colletotrichum gloeosporioides f. alatae DURANTE SU CULTIVO EN MEDIOS LÍQUIDOS
Growth and Development of Colletotrichum gloeosporioides f. alatae During Culture in Liquid Medium
LAURA E. CERÓN RINCÓN1, BLANCA L. HIGUERA M.2, JIMENA SÁNCHEZ N.1, SILVIA BUSTAMANTE3, GUSTAVO BUITRAGO3
1Departamento de Biología,
2Departamento de Química,
3Instituto de Biotecnología,
Universidad Nacional de Colombia, Sede Bogotá.
Presentado agosto 1 de 2005, aceptado octubre 21 de 2005, correcciones diciembre 9 de 2005.
RESUMEN
Algunas características descritas como factores de virulencia para el género Colletotrichum sp. tales como la masa del micelio producido, la esporulación, la actividad poligalacturonasa y el pH del medio, fueron evaluadas durante el cultivo de C. gloeosporioides f. alatae en tres medios líquidos (Czapeck, caldo Martin y caldo papa), utilizados comúnmente para el desarrollo de hongos y en el medio de cultivo Czapeck adicionado con extracto de tubérculo de ñame como única fuente de carbono. Al cabo de 17 días de crecimiento, se obtuvieron los niveles máximos de los parámetros mencionados, al utilizar este último medio, respecto a los otros medios de cultivo evaluados. El medio de cultivo implementado con extracto de tubérculo de ñame, suministró los requerimientos nutricionales del hongo para el desarrollo de factores relacionados con los mecanismos de infección que pueden participar en su patogenicidad.
Palabras clave: antracnosis, ñame, factores de virulencia, Colletotrichum sp., Dioscorea sp.
ABSTRACT
Some characteristics known as virulence factors for Colletotrichum sp. genus, like: weight of the produced mycelium, sporulation, poligalacturonase activity and pH medium were evaluated during the growth of C. gloeosporioides f. alatae in three liquid medium commonly used for fungi culture (Czapeck, Martin broth and potato broth) and additionally (Czapeck) with yam extract as the only source of carbon. After of 17 days of growth, maximum values were obtained for the above parameters in the last medium, compared with others growth media evaluated. The implemented medium with yam extract, supply nutritional requirements of the pathogen for the development of characteristic factors related with mechanism of infections that may play a role in the pathogenesis.
Key words: Anthracnosis, yam, virulence factor, Colletotrichum sp., Dioscorea sp.
INTRODUCCIÓN
Dentro de las especies vegetales que se cultivan tradicionalmente en Colombia, el ñame (Dioscorea sp.) dejó de ser utilizado, solamente para el abastecimiento del mercado local, pues constituye un alimento básico para la economía campesina de la Costa Atlántica, para convertirse en un cultivo de exportación. En 1989 se sembraron más de 25.000 hectáreas de ñame; sin embargo, la producción se vio drásticamente afectada por una infección masiva del hongo Colletotrichum gloeosporioides causante de la antracnosis (Singh et al., 1966; Winch et al., 1984; Abang et al., 2003; Pérez et al., 2003), ocasionando la reducción del área sembrada, que en 1990 llegó a mil hectáreas. La alta incidencia de la antracnosis en la Costa Atlántica colombiana ha afectado drásticamente la producción llegando a ocasionar pérdidas hasta en un 85% (Green et al., 2000), por lo cual esta enfermedad se considera como el factor limitante más importante de la producción colombiana. En Nigeria, donde se produce el 75% del ñame del mundo, la antracnosis se ha encontrado a lo largo de la zona de cultivo y particularmente frecuente y severa en la zona agroecológica del bosque húmedo (Green et al., 2000). La enfermedad se puede presentar en tubérculos, hojas, pecíolos y/o tallos. Inicialmente las hojas afectadas presentan en el haz puntos rojizos de apariencia hendida con halo amarillo. En el envés se observa ennegrecimiento de las nervaduras. Luego las lesiones crecen en forma irregular y se unen entre sí ocasionando finalmente necrosis en la hoja. Otro síntoma de la enfermedad es el ennegrecimiento, muerte apical de los tallos, ocasionando finalmente la muerte descendente de la planta. En algunos casos ocurre necrosis en el pecíolo produciendo defoliación de la planta (Alvarez, 2000). La severidad de esta enfermedad, conlleva el excesivo uso de fungicidas resultando una práctica costosa, poco eficaz y nociva para el ambiente (Pérez et al., 2003). Se suma a estas dificultades, la escasa investigación que existe sobre este cultivo y la biología de sus patógenos.
Las especies de Colletotrichum sp. causantes de la antracnosis en diversos cultivos, exhiben dos fases principales de nutrición, durante la colonización de la planta; la fase inicial biotrófica en la cual se obtienen los alimentos de las células vivas huésped, y la segunda fase tardía necrotrófica donde los alimentos se obtienen de las células hospederas muertas a causa del ataque del patógeno (Bailey et al., 1992). La fase biótrofa es de corta duración y en ésta se asegura el establecimiento del patógeno, sin daños severos en el tejido vegetal. La expresión enzimática para degradar la pared vegetal está estrictamente limitada durante esta fase y la planta hospedera parece no reconocer al patógeno. En consecuencia no se desencadena respuesta de defensa (Perfect et al., 1999). A la fase necrótrofa se asocia la aparición de los síntomas de la antracnosis, con una estrecha relación entre dicha aparición, el incremento en la expresión enzimática para degradar la pared celular vegetal y la virulencia del patógeno (Centis et al., 1997). El crecimiento micelial in vitro de Colletotrichum sp. presenta algunas similitudes con el desarrollo necrótrofo, en lo que se refiere a la expresión enzimática (Shih et al., 2000), la rápida colonización y utilización de la fuente de alimento y la producción de amonio en respuesta a pH ácido in vitro, incrementando de esta forma el pH hasta obtener el nivel óptimo para la actividad pectato liasa (Mendgen y Hahn, 2000). Se ha reportado (Drori et al., 2003; Prusky et al., 2001; Yakoby et al., 2000), para algunos hongos asociados a la antracnosis (Colletotrichum sp.) pre y poscosecha de frutos, el incremento del pH del medio, tanto en el crecimiento necrótrofo como en el saprófito, la relación directa que existe entre este fenómeno y la activación de enzimas relacionadas con la degradación de pectinas; así como (Benito et al., 2000) la importancia de estos factores en el desarrollo de la patogenicidad de hongos y bacterias. Sin embargo, también se reporta (Centis et al., 1997) que el gen clpg2 de Colletotrichum lindemuthianum, que codifica una endopoligalacturonasa, se expresa durante todos los estadíos de la infección y es inducible en cultivo por la adición de pectina incluso a pH ácido. La búsqueda de alternativas duraderas para el control de enfermedades vegetales y que además sean amables con el ambiente, como es el caso del uso de elicitores fúngicos (moléculas derivadas del hongo que son capaces de estimular mecanismos metabólicos de defensa vegetal) requiere, entre otros, del conocimiento de la biología y de las características óptimas de cultivo del patógeno. El objetivo de este trabajo fue evaluar algunos parámetros de desarrollo del patógeno C. gloeosporioides f. alatae como son la masa del micelio producido, la esporulación, la actividad de la enzima poligalacturonasa (E.C. 3.2.2.15) excretada al medio y el pH del medio, durante su cultivo en varios medios líquidos. Los resultados obtenidos generan conocimiento acerca del desarrollo del patógeno causal de la antracnosis del ñame y contribuyen a la búsqueda de alternativas para el control de la enfermedad y al desarrollo de la ciencia y la agricultura locales.
MATERIALES Y MÉTODOS
EL PATÓGENO
Colletotrichum gloespoiroides f. alatae se obtuvo del banco de germoplasma mundial de hongos localizado en la ciudad de Ámsterdam-Holanda. El hongo se conservó en medio de cultivo agar papa dextrosa (PDA) en oscuridad y a una temperatura de 25º C.
Se reactivó la virulencia del hongo inoculando plantas de ñame (Dioscorea rotundata var. Espino) cultivadas in vitro, con 5 mL de una suspensión de 1,5 x 108 conidias/mL, que se inyectó con una jeringa (29G ½'') por el envés de cinco peciolos por plántula. Las plántulas fueron suministradas por el Laboratorio de Cultivos Vegetales del Departamento de Biología de la Universidad Nacional de Colombia, sede Bogotá. La suspensión de conidias se obtuvo lavando con agua destilada estéril cultivos de 15 días de crecimiento en caja de Petri con agar PDA y determinando la concentración por conteo (Madigan et al., 2000), en cámara de Neubauer.
ESTABLECIMIENTO DE CULTIVO DE Colletotrichum gloeosporioides f. alatae EN MEDIOS LÍQUIDOS
Los medios utilizados para el cultivo del hongo fueron: 1) caldo papa dextrosa (Difco); 2) caldo Martin (1 g K2HPO4, 0,5 g MgSO4*H2O, 5 g peptona y 10 g sacarosa, para 1 L de medio); 3) Czapeck (1 g K2HPO4, 0,5 g NaCl, 13,3 mg EDTA, 10 mg FeSO4*7H2O, 0,5 g MgSO4*H2O, 2 g NH4NO3, 6,9 mg MnSO4*4H2O, 3,54 mg ZnSO4*7H2O, 0,01 mg CuSO4*5H2O y 30 g sacarosa, para 1 L de medio) y 4) Czapeckñame (con extracto del tubérculo de ñame como única fuente de carbono, a una concentración en azúcares de 30 g/L de medio). El extracto se obtuvo a partir de tubérculo de Dioscorea rotundata variedad espino de aproximadamente dos meses posteriores a la cosecha. Para ello el tubérculo se peló, se sumergió en agua en ebullición por dos minutos; posteriormente se homogeneizó en licuadora y la suspensión resultante se centrifugó a 10.000 r.p.m. por 10 minutos; se tomó el sobrenadante y se le determinó azúcares totales por el método de Dubois. Los medios se ajustaron a pH 3,8 con ácido láctico, se sirvieron en frascos de 25 mL de capacidad (5 mL/frasco) y se esterilizaron en autoclave por 15 minutos a 121º C y 15 psi. Los cultivos se iniciaron en una concentración de 0,5 x 106 conidias/mL, obtenidas a partir de hongo al cual se le activó previamente su virulencia con 15 días de crecimiento en caja de Petri, y se mantuvieron durante 39 días en oscuridad a 25º C y con agitación constante (150 r.p.m.) siguiendo lo descrito por Shih et al. (2000).
MUESTREOS Y TRATAMIENTOS DE RESULTADOS
A partir de los cultivos del patógeno en los diferentes medios líquidos se determinaron, como parámetros de crecimiento y desarrollo, la masa seca de micelio y la esporulación y como parámetros fisiológicos, la actividad poligalacturonasa (E.C. 3.2.2.15) excretada al medio y el pH del mismo, por los métodos que se describen a continuación. El material experimental (frascos) se dispuso en un diseño completamente al azar con cuatro tratamientos (medios de cultivo) y tres réplicas para cada uno. Después de establecer los medios de cultivo (tratamientos) como se describió anteriormente, se recolectaron tres (réplicas) por cada medio a los 4, 8, 10, 14, 17, 20, 25, 29, 35 y 39 días de cultivo. Para comparar las variables fisiológicas, de crecimiento y desarrollo, el análisis de los resultados se realizó por análisis no paramétrico de Kruskal-Wallis (p<0,05) usando el programa estadístico STATGRAFICS 6.0.
DETERMINACIÓN DE CRECIMIENTO MICELIAR Y ESPORULACIÓN
Para establecer las curvas de crecimiento del patógeno se determinó la masa seca del micelio obtenido de los cuatro medios líquidos evaluados. El micelio se separó del medio de cultivo por centrifugación (12.000 r.p.m. x 10 min a 4º C), siguiendo lo reportado por Yakoby et al. (2000). A continuación se lavó dos veces con agua destilada y se secó a una temperatura de 40º C, hasta peso constante. Para evaluar la producción de conidias, éstas se separaron del micelio por agitación en vortex durante un minuto y su concentración (conidias/ mL de medio) se determinó por conteo (Madigan et al., 2000) en cámara de Neubauer.
DETERMINACIÓN DE ACTIVIDAD POLIGALACTURONASA (E.C. 3.2.2.15) Y PH DEL MEDIO
La determinación de la actividad poligalacturonasa requiere que sean retirados previamente los azúcares reductores presentes en el medio de cultivo, pues su presencia interfiere con el ensayo de cuantificación. Por lo tanto, una vez se retiró el micelio del medio de cultivo por centrifugación, como se describió anteriormente, el sobrenadante obtenido se sometió a diálisis contra agua desionizada, con cuatro cambios a 4º C durante 24 horas. El ensayo se realizó por duplicado siguiendo el protocolo descrito por Boundart et al. (2003), con algunas modificaciones. El método de determinación de la actividad poligalacturonasa se basa en la determinación de azúcares reductores liberados a partir de un sustrato de la enzima como el ácido poligalacturónico. La determinación de los azúcares reductores se realizó por el método colorimétrico de Nelson, (1944) modificado por Somogy (1945). El pH de los medios de cultivo se determinó por triplicado a cada tiempo evaluado, usando electrodo de referencia Ag/Ag (Schoot®).
RESULTADOS Y DISCUSIÓN
ACTIVACIÓN DE LA VIRULENCIA DE Colletotrichum gloeospoiroides f. alatae
Se reactivó la virulencia del hongo inoculando plántulas de D. rotundata var. Espino. Los signos y síntomas característicos de la enfermedad (lesiones necróticas) se registraron sobre las hojas a los cuatro días posteriores a la inoculación, como se muestra en la figura 1). Los órganos vegetales afectados se colocaron en medio PDA de donde se aisló el patógeno, con lo cual se confirmó su virulencia. El hongo se cultivó nuevamente en medio sólido PDA en el que se incubó por 15 días. A partir de estos cultivos, posteriormente se obtuvo la suspensión de conidias para inocular los medios, por los procedimientos descritos anteriormente.
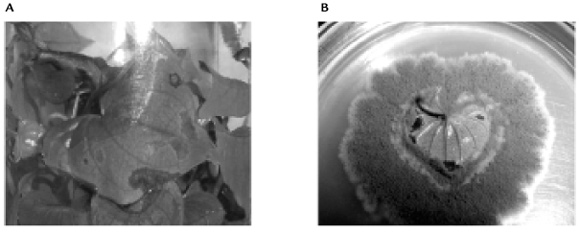
Figura 1. Activación de la virulencia de C. gloeosporioides f. alatae. A. Observación de signos y síntomas (lesiones necróticas) registrados cuatro días después de la inoculación. B. Cultivo en PDA obtenido a partir de los órganos vegetales afectados.
CRECIMIENTO MICELIAR, ESPORULACIÓN, PH Y ACTIVIDAD POLIGALACTURONASA DURANTE EL CULTIVO DE Colletotrichum gloeosporioides f. alatae EN MEDIOS LÍQUIDOS
Como parámetro de crecimiento se determinó la masa del micelio (Fig. 2), obteniéndose valores que fueron significativamente diferentes (p = 4,7 x 10-9) a través del período de estudio. Como se puede observar en la figura 2, en el medio Czapeck-ñame no solo se obtuvo la mayor masa miceliar (20 días), sino que el desarrollo de ésta fue superior (alrededor del doble), con relación al obtenido con los demás medios empleados. Aunque inicialmente la masa de micelio obtenido en Czapeck y Czapeck-ñame fue similar (no hay diferencias significativas entre 4 a 14 días), a partir del día 14 la masa miceliar obtenida en Czapeck se mantuvo constante hasta el final del experimento, correspondiendo en promedio solo al 58% del peso que se obtuvo con Czapeck-ñame. A partir del día 20, el micelio obtenido en czapeck-ñame se mantuvo sin diferencias significativas hasta el final de experimento. Los resultados obtenidos en los caldos Martin y papa no presentaron diferencias durante el experimento y alcanzaron en promedio entre 32 a 36% del crecimiento observado en Czapeck-ñame.
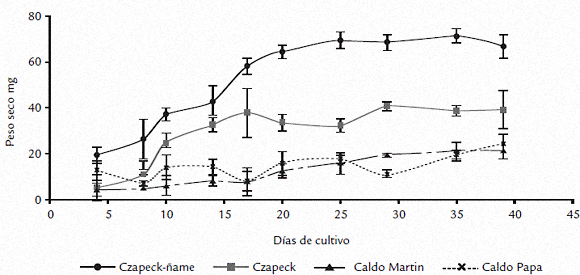
Figura 2. Curvas de crecimiento de C. gloeosporioides f. alatae en los medios de cultivo líquidos empleados Czapeck-ñame, Czapeck, caldo Martin y caldo papa. Los datos se presentan como el promedio de tres réplicas y las barras verticales representan la desviación estándar.
Otro parámetro de desarrollo evaluado fue la producción de esporas (conidias/mL de medio), resultados que se ilustran en la figura 3. La esporulación fue significativamente diferente (p = 6,35 x 10-9) a través del período de estudio. Durante el desarrollo del experimento el conteo de conidias en Czapeck-ñame no solo fue significativamente mayor en todos los tiempos evaluados, sino que llegó alrededor de 108 conidias/mL (día 17), mientras que en los demás medios de cultivo, no superó 106 conidias/mL durante el tiempo que duró el estudio. Entre los cuatro y 20 días de cultivo no hubo diferencias entre los conteos de los medios Czapeck, caldo Martin y caldo papa. Las diferencias se registraron a partir del día 25 en el caldo Martin en el que se alcanzaron en promedio 2 x 106 conidias/mL y a partir del día 30 en Czapeck, en el que se obtuvieron 2 x 107 conidias/mL. En el caldo papa no se registró aumento en esta variable sino hasta el final del experimento (día 39), cuando fue significativamente diferente para los cuatro medios empleados. En este tiempo, en el caldo papa se alcanzaron valores del orden de magnitud de los que se observaron en el caldo Martin a los 25 y 30 días. Se sugirió como factor de virulencia para Colletotrichum sp. la secreción de amonio al medio con el consecuente incremento en el pH (Drori et al., 2003; Prusky et al., 2001; Yakoby et al., 2000) y se demostró para el caso C. gloeosporioides (Penz.) una relación directa entre este fenómeno y la activación de enzimas relacionadas con la degradación de pectinas (Yakoby et al., 2000) y en general, en otros patosistemas, para la activación de enzimas degradadoras de pared vegetal. Igualmente, se estudiaron la variación del pH y de la actividad poligalacturonasa en los medios líquidos evaluados, durante el cultivo de C. gloeosporioides f. alatae, resultados que se muestran en las figuras 4 y 5 respectivamente. El análisis estadístico mostró que el pH de los medios fue significativamente diferente (p = 1,4 x 10-9) a través del período de estudio. Como se ilustra en la figura 4 el hongo cultivado en el medio Czapeck-ñame, a los 4 días incrementó significativamente el pH desde 3,8 (t=0) hasta 5,6. Esta variable siguió en aumento hasta alcanzar pH 8 a los 17 días y a partir de este momento los incrementos por el tiempo de cultivo no fueron significativos. Los medios Czapeck y caldo Martin alcanzaron el nivel de pH obtenido en Czapeck-ñame a los cuatro días, pero a los 10 y 29 días respectivamente. Esta variable continuó en ascenso en estos medios, alcanzando un mismo nivel al final del experimento (día 39), nivel que fue sin embargo, significativamente menor que el valor máximo obtenido para Czapeck-ñame. En el caldo papa se dieron incrementos en el pH a partir de los 29 días y hasta el final (día 39), pero todos fueron significativamente menores con respecto a Czapeck-ñame, czapeck y caldo Martin.
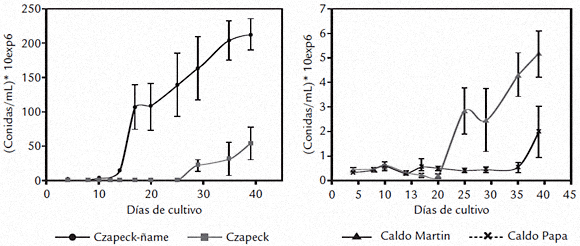
Figura 3. Esporulación durante el cultivo de C. gloeosporioides f. alatae en diferentes medios de cultivo líquidos.
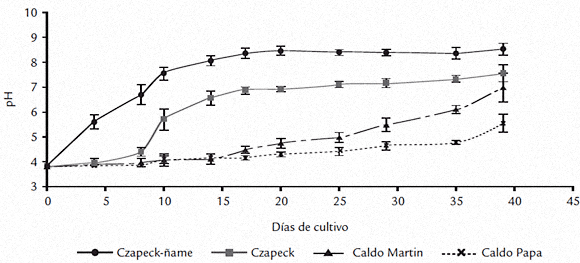
Figura 4. Variación del pH en medio líquido durante el cultivo de C. gloeosporioides f. alatae. (0 días pH = 3,8)
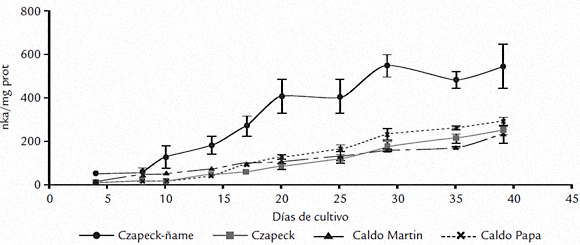
Figura 5. Actividad poligalacturonasa de C. gloeosporioides f. alatae durante cultivo en medio líquido.
Se determinó el efecto del pH del medio, en la producción y secreción de proteínas en varios microorganismos y en gran número de especies del género Colletotrichum sp. (revisado por: Drori et al., 2003; Yakoby et al., 2000), particularmente en antracnosis pre y poscosecha de frutos como tomate, aguacate, manzana, entre otros. La alcalinización del tejido hasta un pH óptimo para la actividad pectato liasa, se consideró como parte de los factores que incrementan la virulencia de C. gloeosporioides y depende de la disponibilidad de nitrógeno para ser convertido a amonio (Prusky et al., 2001). C. gloeosporioides f. alatae fue capaz de utilizar diferentes fuentes de nitrógeno, tanto inorgánicas como orgánicas, para elevar el pH del medio, tal como se observó según el pH final alcanzado en Czapeck y en el caldo Martin, los cuales contienen como fuente de nitrógeno NH4NO3 y peptona, respectivamente. Sin embargo, estos dos medios no alcanzaron el pH obtenido en Czapeck-ñame (pH final > 8) medio que contiene como fuente de nitrógeno NH4NO3 y proteínas de extracto de ñame. En consecuencia, es posible que el comportamiento observado en el pH del cultivo en caldo papa, se debió a que éste es un medio relativamente pobre en nitrógeno respecto a Czapeck, Czapeck-ñame y el caldo Martin. Además, en los dos medios que contienen NH4NO3 se presentaron los mayores valores de pH durante la mayor parte de los tiempos de cultivo evaluados. El efecto del pH del medio sobre la actividad poligalacturonasa no es un factor determinante ya que ésta se puede encontrar a pH ácido (Centis et al., 1997). Sin embargo, su producción durante el cultivo se presenta más rápido en medios que contengan pectina que en aquellos que contienen otro tipo de fuente de carbono. Como se muestra en la figura 5 esta enzima se detectó en todos los medios de cultivo, a lo largo del experimento.
Estos resultados se analizaron estadísticamente (Kruskal-Wallis) encontrándose que la actividad poligalacturonasa excretada a los medios fue significativamente diferente (p = 9,9 x 10-10) a través del período de estudio. Desde los cuatro días la actividad en Czapeck-ñame fue significativamente mayor a la de los demás medios; sin embargo, a los ocho días no hubo diferencias significativas entre Czapeck-ñame y el caldo Martin, pero el comportamiento de la enzima poligalacturonasa en Czapeck-ñame a partir del día diez fue marcadamente diferente a los otros medios utilizados. La actividad en Czapeck-ñame aumentó rápidamente entre los diez y los 20 días, llegando a superar en más de cuatro veces la de los otros medios, y a partir del día 20 se mantuvo sin variaciones significativas, mientras que en los demás medios siguió en aumento hasta el final del experimento (día 39), pero solo llegando a la mitad de la actividad enzimática observada en Czapeck-ñame. Aunque se ha determinado que la actividad poligalacturonasa no es un factor determinante en la infección de C. lindemuthianum, endo PG de este patógeno es capaz de elicitar la síntesis de proteínas relacionadas con patogénesis (PRs) en plantas de tabaco (Boudart et al., 2003) y de algodón (Venere et al., 1984), razón por la cual se ha sugerido su importante papel en la activación de las respuesta de defensa del hospedero. Se ha demostrado que genes que codifican para la actividad endoPG se expresan diferencialmente durante la infección del hospedero y durante el crecimiento saprofítico en medios de cultivo (Centis et al., 1997). En este último, su expresión se activó por la presencia de pectina como fuente de carbono, como se observó en Czapeck-ñame, cuya fuente de carbono seguramente contiene pectina. La disponibilidad de nitrógeno, así como la relación carbono-nitrógeno de los medios empleados, también pudo afectar el grado de esporulación que se observó durante el cultivo de C. gloeosporioides f. alatae. Jackson y Schisler (1992) reportan que distintas variaciones de la relación carbono-nitrógeno en el medio de cultivo, tuvieron influencia en: el rendimiento, la morfología y eficiencia de las esporas de Colletotrichum truncatum para causar enfermedad en plantas de Sesbania exalta. Sin embargo, en Czapeck-ñame no solo la esporulación sino las demás variables estudiadas, mostraron un comportamiento muy distinto en comparación a los otros medios, por lo que se debe tener en cuenta que la influencia sobre estas variables está dada, además, por la compleja composición que puede tener Czapeck-ñame, ya que los mucílagos que se pueden extraer del tubérculo, están compuestos por una serie de proteínas, azúcares, minerales y vitaminas (Perea y Buitrago, 2000).
Se aisló Colletotrichum gloeosporioides de hojas, tallos y tubérculos de ñame que presentaron el síntoma de la antracnosis (Abang et al., 2003; Pérez et al., 2003), sin embargo, no se conocen los factores de virulencia que determinaron el desarrollo del patógeno en los tubérculos, factores que podían estar relacionados con las variables estudiadas, ya que estas mostraron un comportamiento particular en el medio Czapeck-ñame y marcadamente diferente al encontrado en los demás medios empleados. En Czapeck-ñame la dinámica de las variables en estudio fue similar, ya que en el período comprendido entre los 17 a 25 días, se alcanzaron los máximos en el desarrollo del micelio, el grado de esporulación, el incremento del pH y la actividad poligalacturonasa. De este modo se pudo establecer como condiciones de cultivo para Colletotrichum gloeosporioides f. alatae, el medio Czapeck-ñame (pH 3,8) iniciándolo con 0,5 x 106 conidias/mL, y manteniendo el cultivo en la oscuridad, a 25º C y con agitación constante (150 r.p.m.); condiciones adecuadas para la producción masiva de masa miceliar del patógeno, que puede ser utilizada para fines de obtención de elicitores encaminados a activar las respuestas de defensa de la planta.
CONCLUSIONES
Se implementó un medio líquido para el desarrollo de Colletotrichum gloeosporioides f. alatae, consistente en Czapeck y extracto de tubérculo de ñame como única fuente de carbono. Con este medio se obtuvieron los mayores valores para los parámetros de: crecimiento (masa de micelio seco), esporulación, variación de pH del medio y actividad poligalacturonasa extracelular, frente a tres medios líquidos comúnmente utilizados para el cultivo de hongos. Este estudio no solo aportó elementos de criterio para la selección de condiciones de cultivo que permitieran la obtención de micelio fúngico, sino que además, la implementación del medio con extracto de tubérculo de ñame alcanzó los requerimientos nutricionales del patógeno donde fue posible observar el desarrollo de factores característicos de los mecanismos de infección que podían participar en la patogenicidad.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos a la División de Investigaciones de Bogotá (DIB) por la financiación de este trabajo, al Laboratorio de Cultivos Vegetales del Departamento de Biología de la Universidad Nacional de Colombia, Sede Bogotá, dirigido por la profesora Margarita Perea Dallos, por suministrar las plántulas in vitro de ñame Dioscorea rotundata var. espino.
BIBLIOGRAFÍA
1. ABANG MM, WINTER S, MIGNOUNA HD, GREEN KR, ASIEDU R. Molecular Taxonomic, Epidemiological and Population Genetic Approaches to Understanding Yam Anthracnose Disease. Afr J Biotechnol. 2003;2:486-496. [ Links ]
2. ÁLVAREZ A. Prácticas agronómicas para el cultivo del ñame. En: Ñame: producción de semillas por biotecnología. Ed.: Mónica Guzmán y Gustavo Buitrago. Universidad Nacional de Colombia. Editorial Unibiblos. Bogotá, D.C.; 2000;33-39. [ Links ]
3. BAILEY JA, OCONNELL RJ, PRING RJ, NASH C. Infection Strategies of Colletotrichum Species. En: Colletotrichum: Biology, Pathology and Control (Bailey JA y Jeger MJ, Eds.). CAB International Wallingford; 1992;88-120. [ Links ]
4. BENITO EP, ARRANZ M, ESLAVA A. Factores de patogenicidad de Botrytis cinerea. Rev Iberoam Micol. 2000;17:S43-S46. [ Links ]
5. BOUDART G, CHARPENTIER M, LAFITTE C, MARTÍNEZ Y, JAUNEAU A, GAULIN E, ESQUERRE-TUGAYE MT, DUMAS B. Elicitor Activity of a Fungal Endopolygalacturonase in Tobacco Requires a Functional Catalytic Site and Cell Wall Localization. Plant Physiol. 2003;131:93-101. [ Links ]
6. CENTIS SI, GUILLAS N, SÉJALON MT, ESQUERRÉ-TUGAYÉ. Endopolygalacturonase Genes from Colletotrichum: Cloning of clpg2 and Comparison of its Expression to that of clpg1 During Saprophytic and Parasitic Growth of the Fungus. Molecular Plant Microbe Interactions. 1997;10:769-775. [ Links ]
7. DRORI N, KRAMER-HAIMOVICH H, ROLLINS J, DINOOR A, OKON Y, PINES O, PRUSKY D. External pH and Nitrogen Source Affect Secretion of Pectate lyase by Colletotrichum gloeosporioides. Appl Environ Microbiol. 2003;69:3258-3262. [ Links ]
8. GREEN KR, ABANG MM, LLOBA C. A rapid Bioassay for Screening Yam Germplasm for Response to Anthracnose. Tropical Science. 2000;40:132-8. [ Links ]
9. JACKSON MA, SCHISLER DA. The Composition and Attributes of Colletotrichum truncatum Spores are Altered by the Nutritional Environment. Appl Environ Microbiol. 1992;58:2260-1265.
10. MADIGAN MT, MARTINKO JM, PARKER J. Brock Biología de los microorganismos. Octava edición. Prentice Hall Iberia. Madrid; 2000;155. [ Links ]
11. MENDGEN K, HAHN M. Plant Infection and the Establishment of Fungal Biotrophy. Trends Plant Sci; 2000;1-4. [ Links ]
12. PEREA M, BUITRAGO G. Aplicación de la biotecnología agrícola al cultivo de ñame. En: Ñame: producción de semillas por biotecnología. Ed.: M Guzmán y G Buitrago. Universidad Nacional de Colombia. Editorial Unibiblos. Bogotá, D.C.; 2000;17-19. [ Links ]
13. PÉREZ LM, BAQUERO MJ, BELTRAN JD. Caracterización morfológica y patogénica de Colletotrichum spp. como agente causal de la antracnosis en ñame Dioscorea sp. Rev Col Biotecnol. 2003;V:24-35. [ Links ]
14. PERFECT SE, HUGHES HB, OCONNELL RJ, GREEN JR. Colletotrichum: A Model Genus for Studies on Pathology and Fungal-Plant Interactions. Fungal Genet Biol. 1999;27:186-198. [ Links ]
15. PRUSKY D, McEVOY JL, LEVERENTZ B, CONWAY WS. Local Modulation of Host pH by Colletotrichum species as a Mechanism to Increase Virulence. Molecular Plant-Microbe Interaction. 2001;14:1105-1112. [ Links ]
16. SINGH RD, PRASAD N, MATHUR RL. On the Taxonomy of the Fungus Causing Anthracnose of Dioscorea alata L. Indian Phytopathology. 1966;19:65-71. [ Links ]
17. SHIH J, WEI Y, GOODWIN PH. A Comparation of the Pectate lyase Genes Pel-1 and Pel-1 of Colletotrichum gloeosporioides f. sp. malvae and the Relationship Between Their Expression on Culture and During Necrotrophic Infection. Gene. 2000;243:139-150.
18. VERENE RJ, BRINKERHOFF LA, GHOLSON RK. Pectic Enzyme: An Elicitor of Necrosis in Cotton Inoculated with Bacteria. Proc Okla Acad Sci. 1984;64:1-7. [ Links ]
19. WINCH JE, NEWHOOK FJ, JACKSON GVH, COLE JS. Studies of Colletotrichum gloeosporioides disease on Yam, Dioscorea alata, in the Solomon Islands. Plant Pathol. 1984;33:467-77. [ Links ]
20. YAKOBY N, KOBILER I, DINOOR A, PRUSKY D. pH Regulation of Pectate lyase Modulates the Attack of Colletotrichun gloeosporioides on Avocado Fruits. Appl Environ Microbiol. 2000;66:1026-1030. [ Links ]

