










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.11 no.1 Bogotá June 2006
ÉXITO DE ANIDACIÓN DE LA GARZA REAL Egretta alba (Aves, Ardeidae) EN EL DEPARTAMENTO DE CÓRDOBA, COLOMBIA
Nesting Success of the Great-White Heron (Egretta alba) in the Department of Córdoba, Colombia
JAVIER RODRÍGUEZ-BARRIOS1, FRANCISCO TRONCOSO2
1Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá.
2Fundación Ornitología Sierra Nevada FOSIN.
Presentado agosto 22 de 2005, aceptado octubre 30 de 2005, correcciones enero 16 de 2006.
RESUMEN
El éxito de anidación de la garza real (Egretta alba) durante un período reproductivo, se determinó en una camaronera en el departamento de Córdoba (Noroeste de Colombia) entre los meses de julio y noviembre de 2001. Examinamos la influencia de la disposición de los nidos hacia el interior de la colonia y su localización vertical en los árboles, sobre el éxito de anidación de esta garza, encontrando que los dos factores están significativamente relacionados con el porcentaje de éxito de anidación. El análisis de regresión lineal mostró una relación positiva entre el éxito de anidación y la distancia hacia el interior del manglar. Al final del período de anidación el 52% de las parejas anidantes fueron exitosas, con un aumento en el tamaño de la población cercano al 100%.
Palabras clave: anidación, garza real, Córdoba, Colombia.
ABSTRACT
Between July and November 2001, we evaluated the nesting success of the Great-white Heron (Egretta alba) during one breeding period at a shrimp pond in northeastern Colombia. We examined the relationship between nesting success and two nest variables: position within the colony (peripheral vs. central), and vertical location on sampled trees, finding a significant relationship between each pair of variables. Regression analysis showed a positive relation between nesting success and distance from the center of the mangrove. By the end of the nesting period, 52% of the nesting couples were successful, increasing the population size by 100%.
Key words: Nesting, Great-white Heron, Córdoba, Colombia.
INTRODUCCIÓN
Las aves silvestres viven en un medio ambiente fluctuante y poseen un patrón temporal no uniforme de requerimientos de energía (Alfaro y Russi, 1986). Su supervivencia y éxito reproductivo dependen del desarrollo de un programa de regulación de tiempo eficiente que les permita el ajuste de las funciones fisiológicas más importantes a períodos óptimos. De forma tal que la carga genética de las parejas que produzcan crías en épocas no aptas, será eliminada o reducida por selección natural (Odgen, 1994). La garza real es una especie solitaria, aunque anida en colonias (Arancibia y Sepúlveda, 1991). Esta es una especie residente común de manglares, ríos y lagos que puede vivir desde el nivel del mar hasta 2.600 m de altura (Hilty y Brown, 1986). Su distribución geográfica incluye prácticamente cualquier parte del mundo, desde zonas tropicales hasta Canadá o Chile, siendo una especie cosmopolita y eurictona (Olivares, 1973), pues su área de dispersión ocupa en los continentes porciones dilatadas. Se reportan hasta dos períodos reproductivos. La primera época se presenta de enero a julio y la segunda se presenta a finales de julio y dura hasta inicios de septiembre (revisado por: Hilty y Brown, 1986). Amontona gran cantidad de palos delgados y secos hasta formar una plataforma de estructura sencilla pero resistente (Olivares, 1973), por encima de los dos metros en mangles, cactus o grandes árboles (Hilty y Brown, 1986). Pone de dos a cinco huevos de color azul celeste, los cuales miden 51 a 63 mm de largo y de 39 a 42 mm de ancho (Hilty y Olivares, 1973; Brown, 1986). La ubicación de los nidos es un factor de mucha importancia en cuanto a las colonias mixtas se refiere como ocurre con las poblaciones de esta especie que acostumbra a desarrollar sus procesos de anidación junto a otras especies de manera simultánea (Alfaro y Russi, 1986). En cuanto al éxito de los polluelos se ha comprobado que existe un mayor porcentaje de supervivencia entre los dos primeros polluelos con relación al tercero y al cuarto (Alfaro y Russi, 1986; Custer, 1989; Parker et al., 1989; Guster y Peterson, 1991). El éxito de anidación de esta especie ha sido utilizado como un indicador de las condiciones en que se encuentran los humedales ocupados por ella (Custer y Osborn, 1977), proporcionando información valiosa en los períodos críticos como las sequías extremas, los aumentos en la salinidad de los estuarios y efectos antrópicos, con base en el comportamiento temporal de sus fases reproductivas en orden de magnitud y ubicación espacial (Frederick y Splanding, 1994). La dinámica de las poblaciones de esta garza, se usa para explicar, densidad de los bosques y el nivel de las aguas en los humedales (Hofman et al., 1994; Frederick y Loftus, 1993). Con este trabajo se evaluó el éxito de anidación de la garza real E. alba en una colonia de anidación ubicada en el sector de Agrosoledad, bahía de Cispata (golfo de Morrosquillo, Colombia) durante la segunda fase de anidación del año 2001 que corresponde a los meses de julio a noviembre. Esto se desarrolló tratando de relacionar dicho éxito con dos factores en relación a los nidos: (a) disposición hacia el interior de la colonia y (b) su localización vertical en los árboles.
METODOLOGÍA
ÁREA DE ESTUDIO
El estudio se llevó a cabo en el sector de Agrosoledad, situado en la costa caribe de Colombia (golfo de Morrosquillo), al este del río Sinú, en el municipio de Cispata. La laguna costera es el antiguo estuario de la desembocadura del río Sinú, el cual actualmente desemboca en el mar Caribe al este de Montería y al oeste del área de estudio (Fig. 1).
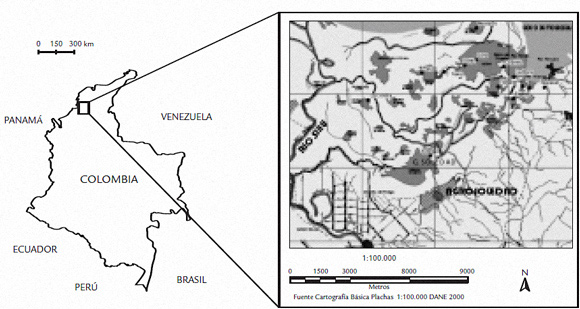
Figura 1. Zona de estudio. Fuente: INVEMAR 2001-1002. Plan de seguimiento y monitoreo del taico estuarino del río Sinú.
La colonia de reproducción de la garza real en este sitio se encuentra dentro de uno de los cuatro biofiltros (biofiltro el Garcero) que purifican aguas para levante de langostinos (Pennaeus bahamei). El área de incubación (7 Ha) corresponde únicamente a la población de Mangle Bobo (Laguncularia racemosa). Esta especie de mangle, es la más abundante en el biofiltro y se encuentra sumergida aproximadamente en 1,10 m de agua proveniente de los estanques camaroneros (Fig. 2). La profundidad de las lagunas de manglar del biofiltro, brinda protección a las garzas contra depredadores terrestres potenciales para sus nidos.
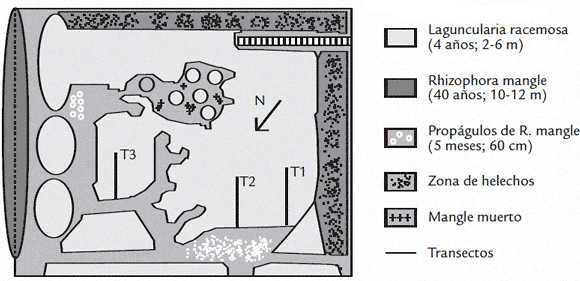
Figura 2.Ubicación del garcero dentro del biofiltro.
FASE DE CAMPO
En campo, se desarrollaron los muestreos con una periodicidad de ocho días por mes (primera semana de cada mes). Las faenas de campo se desarrollaron por lo general de las 06:00 a las 12:00 y de las 14:00 a las 18:30 durante todos los días, utilizando un bote para transitar entre los caños del garcero. Para estimar el número total de aves adultas en el área de estudio, se utilizó el método de parar y contar para recorridos en accesos posibles con repeticiones (ocho repeticiones en promedio) y así disminuir el error de la estimación para ubicar los sitios de anidación. Por medio de este método se hicieron recorridos en la zona correspondiente a la colonia, durante el primero y último muestreo (julio y noviembre), deteniéndose en puntos equidistantes para contar individuos adultos y juveniles de la especie en estudio (Hennig, 1997). Como resultado se obtuvo el tamaño inicial de la población de garzas y el aporte de nuevos individuos al final de este período de anidación (juveniles exitosos). Se ubicaron tres transectos cuya longitud dependió de la presencia de árboles ocupados por nidos. Estos transectos abarcaron un área igual a un cuarto del área total del garcero según lo propuesto por Hennig (1997). Dentro de estos transectos, se registraron los árboles de incubación, en una extensión aproximada de 25 m a la izquierda y 25 m a la derecha de cada transecto. De esta manera se hizo una estimación de la población de individuos anidantes y su ubicación dentro del garcero. En 50 m de transecto se registró por lo tanto un área de 1/4 de hectárea con repeticiones para disminuir el error de la estimación.
ESTIMACIÓN DEL ÉXITO DE ANIDACIÓN
Para analizar la fenología de la puesta, en cuanto a la distribución de las nidadas, se seleccionó y marcó (con cintas de colores) un máximo de 14 árboles al azar por transecto. La numeración de estos árboles se hizo de un extremo (orilla de los caños) hacia el centro de la colonia. Posteriormente, se realizó un registro del número de huevos y número de polluelos en los árboles correspondientes a cada transecto (primer factor) y la altura de los nidos en los árboles escogidos (segundo factor). Se estimó el porcentaje de éxito de anidación (E) para cada transecto, con base en la relación entre el número máximo de huevos iniciales para los nidos en los árboles muestreados (Nmáx) y el total de polluelos de esos nidos, que lograron volar (Ni) en cada mes de muestreo, como:
E = ——— x 10
Nmáx
Con los datos de éxito temporal promedio para cada transecto, se realizó un estimativo del éxito promedio de toda la colonia de garzas reales (EP) correspondiente al período de anidación muestreado, como:
ET1 + ET2 + ET3
E = ——————————
3
ET1, ET2 y ET3 correspondió al éxito promedio de los transectos.
Finalmente, los valores obtenidos de porcentaje de éxito de anidación (E) para cada transecto en los diferentes árboles y nidos, se relacionaron con los valores obtenidos para los factores limitantes (disposición de los nidos hacia el interior de la colonia y localización vertical de los nidos).
ANÁLISIS DE LA INFORMACIÓN
Se realizó un análisis de regresión lineal y un ANDEVA (modelo II) para encontrar relaciones y diferencias entre el porcentaje de éxito de anidación y los factores evaluados respectivamente, utilizando el paquete estadístico Statgraphics 4.0.
RESULTADOS
Un total de 4.900 aves fue estimado al inicio del período de anidación con errores de estimación de 50 individuos (aproximadamente). Este error se disminuyó por el conteo repetido en el mismo punto, a menos de diez individuos. Al finalizar el período de muestreo (noviembre) se estimó un tamaño de población de garzas de 9.126 (adultos junto juveniles exitosos). Con el registro de aproximadamente un cuarto del área total correspondiente a 754 parejas (Tabla 1), resultó una población reproductiva calculada de 3.016 parejas.
| Transecto | N° de Parejas en anidación | N° de Árboles | Porcentaje |
| 1 | 339 | 46 | 42 |
| 2 | 298 | 38 | 35 |
| 3 | 117 | 25 | 23 |
| Total | 754 | 129 | 100 |
Tabla 1. Total de parejas de anidación por transecto (se incluyen todas las parejas anidantes reportadas para cada transecto).
Con base a las ecuaciones planteadas para el calculó del porcentaje de éxito de anidación se estimó que el 51,9% de los individuos fueron exitosos (Tabla 2), lo que corresponde a 1.565 parejas exitosas (cuyos polluelos lograron volar y salir del nido). Estas 1.565 parejas y el promedio de éxito de puesta de 2,17 huevos (Tabla 3), dan como resultado 4.226 juveniles exitosos. Sumando este número total de juveniles al calculado de la población en el primer mes de muestreo que fue de 4.900 individuos, da como resultado un tamaño poblacional máximo de 9.126 garzas reales entre juveniles exitosos y adultos en la colonia, al final del período de anidación (Tabla 3). Se obtuvieron datos para eventos fenológicos reproductivos, correspondientes a los meses de julio a noviembre del año 2001 en los cuales se encontraron con más frecuencia puestas de dos, seguido por puestas de tres huevos (Tabla 4).
| N | Mínimo | Máximo | Promedio | Desviación |
| 34 | 1 | 4 | 2,17 | 0,019 |
Tabla 2. Éxito de postura de la garza real (registrado para 42 árboles). N: número de árboles muestreados al azar. Mínimo y Máximo: frecuencia mínima y máxima respectivamente de postura por nido.
| Total parejas exitosas | Éxito de anidación (%) | Juveniles exitosos | Tamaño de la población al final del período |
| 1565/ 3016* | 51.9 | 4226/ 8142 | 9126 |
Tabla 3. Resultados generales para la población de garzas reales al final del período de muestreo. *1.565 parejas exitosas de 3.016 parejas en total.
| Tamaño de la puesta | Frecuencia | % Éxito |
| Puesta de 1 Huevo | 40 | 17,0 |
| Puesta de 2 Huevos | 103 | 43,8 |
| Puesta de 3 Huevos | 65 | 27,7 |
| Puesta de 4 Huevos | 27 | 11,5 |
Tabla 4. Tamaño de la nidada determinado en nidos exitosos.
ANÁLISIS DE LA INFORMACIÓN
Se encontraron diferencias entre el porcentaje de éxito de anidación para los árboles muestreados (F13,1078=28,17, p<0,001), observándose gráficamente un incremento significativo del porcentaje de éxito hacia al interior de la colonia (Fig. 3A). Se da una tendencia en el incremento del porcentaje de éxito de anidación con la altura de los nidos sobre la superficie del agua (F32,1059=26,72, p<0,001) (Fig. 3B). De igual forma, se presentaron diferencias en el porcentaje de éxito de anidación de los diferentes transectos (F2,1089=18,8, p<0,001) mostrando una disminución marcada hacia el tercer transecto que corresponde a la zona noreste del garcerío (Fig. 3C).
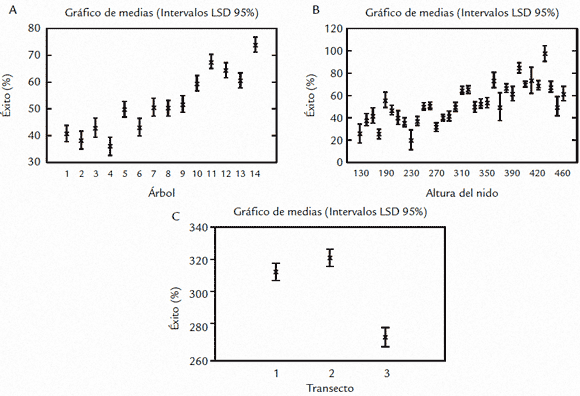
Figura 3. A. Aumento del éxito de anidación hacia el centro de la colonia (primer factor). B. Aumento del porcentaje de éxito en los nidos ubicados a mayor altura. C. Diferencias en el porcentaje de éxito para los tres transectos muestreados.
Se registra una relación lineal en los dos factores evaluados (ubicación de los nidos hacia el interior de la colonia y la posición vertical de éstos) y el porcentaje de éxito de anidación (F1,1090=299,4, p<0,001 y F1,1090=426,2, p<0,001 respectivamente; Figs. 4A y 4B). Ésto sugiere que el éxito de anidación es una función lineal del los factores evaluados, los cuales presentan características importantes en sentido espacial (vertical y horizontalmente) por lo que son considerados como limitantes de este éxito.
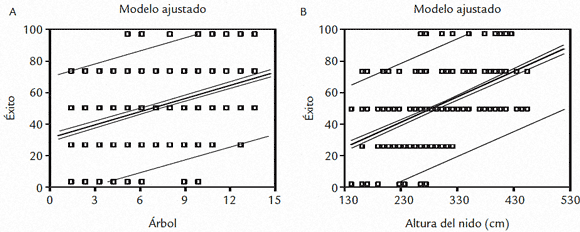
Figura 4. A. Relación directa del éxito reproductivo hacia el centro de la colonia. B. Relación directa del incremento del éxito de anidación con en nidos ubicados a mayor altura.
DISCUSIÓN
El incremento poblacional estimado de E. alba durante este período de anidación fue de de 4.900 a 9.126 individuos contando los juveniles exitosos, lo cual representa un aumento del 93% de la población. Según Frederick y Spalding (1994) y Rodgers (1980) este incremento acelerado de poblaciones de garzas se debe a varios factores ambientales los cuales no fueron evaluados en este estudio, como son: el aporte óptimo de espacio para ubicar los nidos, buen suministro de alimento para los polluelos, baja competencia por recursos y protección contra predadores. Baxter (1994) menciona que ecosistemas de manglar que se encuentran en estados sucesionales tempranos como el estudiado en nuestro trabajo, se caracterizan por presentar una dinámica poblacional de crecimiento rápido en tiempo, debido a las condiciones óptimas en cuanto al aporte de recursos, para que estos organismos sean exitosos en sus historias de vida. Otro aspecto de importancia es la ubicación de los árboles de incubación sobre una columna de agua de aproximadamente 1,3 m de profundidad, lo cual es una condición clave para el éxito de anidación como lo reportan Frederick y Collopy (1989a) y Scharf (1989) quienes encontraron que las colonias que se construyen al menos a 5 a 10 cm encima de la columna de agua, tienen mayor éxito de anidación ya que se protegen contra predadores terrestres como los mapaches, zorros y ratas los cuales afectan altamente a las colonias. Scharf (1989) reporta un cambio de ubicación de las colonias de anidación de la garza real desde las islas y penínsulas hacia los sistemas lóticos y lénticos que presentan poblaciones de manglar parcialmente sumergido para protegerse de los predadores y de disturbios antrópicos.
Varios estudios han demostrado que el abandono y desaparición de estos sitios de anidación se deben principalmente a la perturbación ocasionada por el hombre entre otros factores como los cambios meteorológicos e hidrológicos como lo reporta Frederick y Collopy (1989b), Merrifield y Naughton (1992), entre otros. Si se mira este fenómeno a mediano plazo, se puede considerar a estas colonias como agentes perturbantes en sectores como los estudiados, observando que la proyección anual es a su incremento y además que en las épocas de anidación el nivel de contaminación de las aguas es muy elevado debido a la cantidad de materia orgánica que excretan en forma de guano. Teóricamente se esperaría que este crecimiento acelerado se estabilice cuando el sistema supere su capacidad de carga y el recurso espacio y alimento sea limitante como ocurre con la mayoría de las poblaciones biológicas (Smith y Collopy, 1995; Schogolev, 1996). Esto muestra una relación clara con la zona estudiada, la cual presenta características ideales para que los procesos de anidación y crecimiento poblacional al final de un período de anidación sean exitosos, con protección ante disturbios antrópicos, posible suministro de alimento de las piscinas camaroneras y un ecosistema de manglar con poblaciones de L. racemosa en un estadío sucesional temprano (la edad promedio del bosque del garcero es de ocho años).
Pueden existir eventos que afectan la homogeneidad de los procesos de anidación en sentido espacial, al observar el patrón de incremento del éxito de anidación hacia el centro de la colonia, correspondiente a los árboles de mayor altura. Baxter (1994) estudió varias colonias de garzas en humedales de Australia al igual que trabajos realizados por Chavez-Ramírez y Snack (1995). Éstos reportan la existencia de competencia por espacio de estas poblaciones y de otras aves acuáticas al inicio de los períodos reproductivos. Como resultado los individuos más competitivos ubican sus nidos hacia el centro del humedal, el cual les brinda mayor protección contra predadores terrestres y además es un sitio que corresponde a árboles de mayor tamaño lo cual les facilita ubicar sus nidos a una altura mayor. Finalmente, aparecen individuos menos competitivos que tardarán más tiempo en iniciar su proceso reproductivo, los cuales se ubicarán hacia el borde del manglar. El número de huevos por nido reportado en los árboles de los diferentes transectos fue en promedio de dos huevos, lo cual corresponde a un valor bajo comparado con otros reportes del país que fluctúan entre 2-5 huevos por nido, como lo reporta Olivares (1973), Hilty y Brown (1986) y para Norte América en los Everglades de la Florida (Frederick y Collopy, 189). Se presenta una estratificación vertical de varias especies de garzas para ubicar los nidos en un mismo árbol de incubación. Este patrón fue reportado por Pratt (1972) en las costas de San Francisco y de igual forma Alfaro y Russi (1986) en un humedal de Cartagena. El patrón de ubicación de nidos de la especie E. alba correspondió a la parte superior copa de los árboles. Hacia la parte media y baja del follaje de estos árboles se ubicaban otras especies de garzas de menor tamaño como Egretta thula y Bubulcus ibis. Posiblemente esta sea una distribución del recurso espacio como resultado de algún tipo de competencia interespecífica en la cual, garzas de mayor tamaño escogen la parte alta de los árboles, seguida por garzas de menor tamaño que se ubican en estratos inferiores del mangle (Alfaro y Russi, 1986).
CONCLUSIONES
Los conteos realizados antes y después de la fase de anidación muestran que la población de la garza real aumentó cerca del 100% (de 4.900 individuos hasta 9.126 al final de la etapa). Estos resultados generales revelan de forma clara que el sector estudiado presenta condiciones óptimas para el desarrollo de un proceso exitoso de anidación. Al extrapolar en número de parejas anidantes estimadas en un cuarto de el área total a toda la población (3.016 parejas = 6.032 adultos), el número de individuos calculado es muy superior al determinado en los conteos poblacionales durante el primer mes de muestreo (4.900 individuos aproximadamente). Esto es debido a que la ubicación de las nidadas en la colonia no presenta una distribución homogénea y que más bien, existen lugares en los que se puede encontrar mayor o menor número de parejas anidantes con relación al sector muestreado por los transecto. En promedio se construyeron ocho nidos por árbol. El número máximo está alrededor de 13 nidos por árbol de puesta, lo que muestra claramente la poca cobertura foliar de estos árboles jóvenes. La mayor frecuencia de postura durante esta etapa de anidación correspondió a nidos con dos huevos y muy esporádicamente puestas de cuatro huevos. Este valor es consecuente con el tamaño de los árboles, ya que los árboles de la zona de estudio presentaban un promedio de altura de 4 m (del nivel superficial de la columna de agua hasta el dosel). Claramente se pudo observar la preferencia de esta especie por ubicar sus nidos en los árboles que se encontraban parcialmente sumergidos en la columna de agua, como posible mecanismo de protección contra depredadores terrestres, que en cualquier momento podrían atacar sus nidos. A pesar de que los árboles de incubación son muy jóvenes y el área de anidación es de tan solo 12 hectáreas, el número de individuos pertenecientes a la especie en estudio puede considerarse elevado tanto al comienzo como al final de la fase de anidación (9.126 individuos). Ésto se relaciona debido a que se conjugan varios factores que hacen de este un lugar ideal para la anidación exitosa de esta y otras especies de aves acuáticas del garcero anteriormente mencionados.
La fuente alimenticia fue proporcionada particularmente por las piscinas camaroneras del sector, en las cuales se pudo encontrar poblaciones de adultos extensas en busca del alimento para sus polluelos, el cual consistía en el langostino Pennaeus bahamei el cual es cultivado en Agrosoledad. Se encontró un incremento del éxito de anidación hacia el interior de la colonia, determinado por la ubicación de los árboles a lo largo de los transectos, lo cual corresponde a los árboles de mayor tamaño ubicados hacia el sur y (Figs.2 y 3C). Se encontraron diferencias del éxito de anidación en el transecto 1 y 2 con relación al 3, el cual corresponde a árboles de menor tamaño y los valores más bajos de porcentaje de éxito (Fig. 3C) con un valor de probabilidad p que representa una diferencia significativa muy marcada de este transecto con el resto. No existió una tendencia clara entre el éxito de anidación y la ubicación de los nidos en estrato vertical del los árboles (Fig. 3B), lo cual se puede explicar, debido a que esta colonia se encontraba formada sobre árboles de poca altura que no presentaban diferencias muy marcadas entre sus alturas, aunque gráficamente se observa una tendencia por ubicar los nidos en alturas que oscilan entre dos y tres metros sobre el nivel del agua (Fig. 3B), y en su mayoría en los árboles de mayor tamaño ubicados al sur de la colonia (Fig. 3C), por tener una mayor altura representada en su cobertura foliar, muestran mayor cantidad de nidos por árbol (Fig. 5).
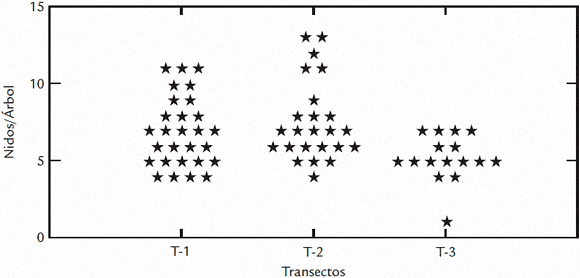
Figura 5. Densidad de nidos en los tres transectos escogidos al azar. (N = 68).
AGRADECIMIENTOS
Al Instituto de Investigaciones Marinas (INVEMAR) por facilitar el transporte hasta la zona de estudio. A la Hacienda Camaronera AGROSOLEDAD S.A. por permitir el desarrollo exitoso de los muestreos y la estadía dentro de sus predios.
Al doctor Óscar David Solano, investigador del Instituto de Investigaciones Marinas (INVEMAR) y todo su grupo de investigación por facilitarme el transporte hasta la zona de estudio.
BIBLIOGRAFÍA
1. ALFARO M, RUSSI D. Estimación del éxito de anidamiento de la garza morena Egretta tricolor (Gosse, 1847) en la laguna de San Lorenzo, Cartagena (Colombia). Boletín Ecotrópica. 1986;19:3-15.
2. ARANCIBIA E, SEPÚLVEDA A. Guía de campo de las aves de Chile (online). Universitaria: Santiago de Chile. 1991. (Citado 15 de febrero de 2000). Disponible por Internet: http://www.ccpo.odu.edu/~andres /aves/089.0.html. [ Links ]
3. BAXTER G. The Influence of Synchronous Breeding, Natal Tree Position, and Rainfall on Egret Nesting Success. Colonial Waterbirds. 1994;17(2):120-129. [ Links ]
4. CHAVEZ-RAMÍREZ F, SLACK, R. Differential Use of Coastal Marsh Habitats by Nonbreeding Wading Birds. Colonial Waterbirds. 1995;18(2):166-171. [ Links ]
5. CUSTER T. Egg Size and Laying Order in Black-Crowned Night-Herons, Snowy Egrets, and Great Egrets. Pacific Seabird Group Bulletin Baxter. 1989;16:24. [ Links ]
6. ____________, OSBORN R. Wading Birds as Biological Indicators: 1975 Colony Survey. U.S.A. Fish Wildl Serv Spec Sci. 1977;206:46-68. [ Links ]
7. FREDERICK P, COLLOPY M. The Role of Predation in Determining Reproductive Success of Colonially Nesting Wading Birds in the Florida Everglades. Condor. 1989a;91(4):860-867. [ Links ]
8. ____________. Nesting Success of Five Ciconiiform Species in Relation to Water Conditions in the Florida Everglades. Auk. 1989b;106(4):625-634. [ Links ]
9. ____________, LOFTUS F. Responses of Marsh Fishes and Breeding Wading Birds to Low Temperatures: A Possible Behavioral Link Between Predator and Prey. Estuaries. 1993;16(2):216-122.
10. ____________, SPALDING M. Factors Affecting Reproductive Succes of Wading Birds (Ciconiformes) in the Everglades Ecosystem. Everglades Ecosystem and its Restoration. Edit. St. Lucie Press. The United States; 1994;659-690. [ Links ]
11. GUSTER T, PETERSON D. Growth Rates of Great Egret, Snowy Egret and Black- Crowned Night-Heron Chicks. Colonial Waterbirds. 1991;14:46-50. [ Links ]
12. HENNIG V. Investigaciones sobre la biología del cormorán o pato cuervo (Phalacrocorax b. brasilianus) en la ciénaga grande de Santa Marta, Colombia. Documento. Corporación Autonoma Regional del Magdalena. Biblioteca General; 1997;48. [ Links ]
13. HILTY S, BROWN W. A Guide to the Birds of Colombia. Princeton University Press. Princeton (USA); 1986;836. [ Links ]
14. HOFMAN W, BANCROFT G, STONG A, SAWICKI R, HOFFMAN W, JEWELL S. Relationships Among Wading Bird Foraging Patterns, Colony Locations, and Hydrology in the Everglades. Everglades the Ecosystem and its Restoration. Ed. Lucie-Press. Florida, The United States; 1994;615-657. [ Links ]
15. MERRIFIELD K, NAUGHTON M. Waterbird Censuses of Cabell Marsh, and 1993. In: Northwest Sci. 1994;68(2):139. [ Links ]
16. ODGEN J. A Comparison of Wading Bird Nesting Colony Dynamics (1931-1946 and 1974-1989) as an Indication at Ecosystem Condition in the Southern Everglades. Everglades Ecosystem and its Restoration. Edit. St. Lucie Press. Florida, The United States; 1994;533-568. [ Links ]
17. OLIVARES A. Las ciconiiformes colombianas. Ed. PROYSER Proyectos y Servicios. Colombia; 1973;303. [ Links ]
18. PARKER G, MOCK D, LAMEY T. How Selfish Should Stronger Sibs Be. Am Nat. 1989;133(6):846-868. [ Links ]
19. PRATT H. Nesting Success of Common Egrets and Great Blue Herons in the San Francisco Bay Region. Condor. 1972;74:447-463. [ Links ]
20. RODGERS J. Reproductive Success of Three Heron Species on the West Coast of Florida. Fla Field Nat. 1980;(8):37-40. [ Links ]
21. SCHARF W. Coastal Great Heron and Great Egret colonies of the Michigan Great Lakes. En: The Jack-Pine Warbler. 1989;67(2):53-65. [ Links ]
22. SCHOGOLEV I. Fluctuations and Trends in Breeding Populations of Colonial Waterbirds in the Dnestr Delta, Ukraine, Black Sea. Colonial Waterbirds. 1996;19(S):91-97. [ Links ]
23. SMITH J, COLLOPY W. Colony Turnover, Nest Success and Productivity, and Causes of Nest Failure Among Wading Birds (Ciconiiformes) at Lake Okeechobee, Florida (1989-1992). Florida USA. Ecological Studies on the Littoral and Pelagic Systems of Lake Okeechobee. 1995;45:287-316. [ Links ]

