



![Ribonucleases: [subtitle]Theurapetical potential on Viral Infections](/img/en/prev.gif)






Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.11 no.2 Bogotá June 2006
CARACTERIZACIÓN DE LA COMUNIDAD DE MACROINVERTEBRADOS DE LA QUEBRADA PALOBLANCO DE LA CUENCA DEL RÍO OTÚN (RISARALDA, COLOMBIA)
Characterization of the Macroinvertebrate Community of Paloblanco Gulch, Otun River Basin (Risaralda, Colombia)
ELEONORA BERNAL, DUBERNEY GARCÍA, MIGUEL A. NOVOA, ATTICUS PINZÓN Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá.
Presentado septiembre 7 de 2005, aceptado octubre 13 de 2005, correcciones enero 11 de 2006.
RESUMEN
Se realizó un estudio de las comunidades de macroinvertebrados presentes en la quebrada Paloblanco, ubicada en la cuenca del río Otún (Risaralda, Colombia). Los puntos de muestreo seleccionados corresponden a la cabecera y la zona cercana a la desembocadura en el río Otún, ambos con características geomorfológicas de corrientes de primer orden. En cada punto se colectaron los macroinvertebrados de las unidades funcionales hojarasca, epiliton, musgo y grava, junto con una muestra de deriva y fauna general (sin discriminar unidad funcional). Se encontraron 42 familias de Insecta, además de Acari, Annelida, Crustacea e Hirudinea. Igualmente se encontraron familias de presencia exclusiva en cada punto, mostrando diferencias en la composición de las comunidades. Los índices ecológicos empleados mostraron una mayor diversidad en punto bajo y diferencias entre algunas de las unidades funcionales, relacionados con el medio y los hábitos presentes. Para el análisis se tuvieron en cuenta los conceptos de continuo y zonación de los ríos. A pesar de tener condiciones de primer orden, existen variaciones a pequeña escala entre los puntos de muestreo, que están relacionadas con la intervención antrópica en la zona aledaña a punto bajo.
Palabras clave: macroinvertebrados, quebrada Paloblanco, deriva, continuo, Colombia.
ABSTRACT
A research on the macroinvertebrate community was made in the Paloblanco gulch, located in the Otun River basin, (Risaralda, Colombia). Two sample points were selected, the headwaters and the mouth’s closer zone in the Otun River and both presented first order river geomorphology characteristics. The functional units (litter, stone, moss and gravel) were collected and conjunctly with derive and general fauna samples (not including discrimination in the functional units) were taken at each point. 42 Insecta families were founded, also Acari, Annelida, Crustacea and Hirudinea.
Equally, exclusive families were found in each point, showing a difference in the community composition. The ecological indexes were employed to show high diversity in the low point and differences among some functional groups, related with the environment and the present habits. Furthermore, the continuum concept and zonation application in the Paloblanco gulch were analyzed. Despite that sample points are first order, small scale variations were found between them which are related to the antropic intervention in the low point surrounding zone.
Key words: macroinvertebrates, Paloblanco gulch, drift, continuum, Colombia.
INTRODUCCIÓN
Las cuencas están caracterizadas por las interacciones geomorfológicas, hidrológicas y biológicas que determinan las comunidades presentes en sus ríos. Observando los cambios graduales en composición taxonómica y características físicas y químicas aparecen dos conceptos importantes: continuo del río referente a la distribución de especies en un río desde su nacimiento hasta su desembocadura (Vannote et al., 1980), y zonación de los ríos, que se refiere a tramos definidos dentro del río en los que el paisaje determina la composición de las comunidades (Illies y Botosaneanu, 1963; Hynes, 1971). El continuo del río habla de cómo los organismos se distribuyen en un río de acuerdo a las características del agua, la disponibilidad de luz, la oferta de alimento, la presencia de depredadores, etc; estos factores determinan la presencia de ciertas familias de organismos en unos sitios y su reemplazo por familias diferentes en otros sitios, pero que cumplen la misma función en el sistema. De otro lado la zonación de los ríos habla de cómo el paisaje altera esta distribución; si es un río de montaña o de llanura, si hay un lago o una cascada, características geomorfológicas del paisaje que cambian suave o drásticamente la distribución de organismos en un río. Otro concepto que se emplea en este trabajo es el de unidades funcionales (UF), las cuales facilitan la descripción de la heterogeneidad espacial del río como un conjunto de sustratos funcionales discretos y cubiertos por agua. Cada unidad está constituida por distintas fracciones de materia orgánica y en su conjunto representan la estructura física y la oferta de recursos alimenticios para la biota de la quebrada (Ospina et al., 2004). Las UF consideradas en este trabajo fueron: Hojarasca. Conformada por acumulaciones sumergidas de hojas y madera de diferentes tamaños y en diferente estado de descomposición que son aprovechadas por hongos, bacterias, meso y macroinvertebrados (Andersson y Sedell, 1979). Epiliton. Comprende fundamentalmente arreglos complejos de bacterias, hongos, protozoos y algas embebidos en una matriz de polisácaridos denominada biopelícula (Lock et al., 1984), además de enzimas extracelulares que se acumulan en la biopelícula y que son las responsables de la conversión de moléculas de alto peso molecular a moléculas de bajo peso molecular que pueden ser asimiladas por los heterótrofos (revisado por Sabater y Romani, 2001). Esta UF constituye buena parte de la producción primaria en ecosistemas lóticos y por consiguiente suele ser visitada por invertebrados en actividades de forrajeo. Grava. Incluye rocas que están en un rango de tamaño de 264 mm (revisado por Ospina et al., 2004) y que son depositadas por la quebrada en las zonas de remansos o localizadas en zonas de corriente moderada. A diferencia de las rocas grandes este sustrato no ofrece la estabilidad mecánica suficiente para la configuración de abundantes comunidades de algas y macrófitas, pero en cambio dispone de muchos espacios intersticiales que son aprovechados por una comunidad compleja de autótrofos y heterótrofos. Musgo. Reune el conjunto de musgos y hepáticas sumergidos y se localizan en zonas expuestas a radiación solar y que igualmente son aprovechadas por diversos grupos de herbívoros acuáticos.
Es así como los estudios de limnología se hacen indispensables para la comprensión de los ríos en el neotrópico, en donde se han encontrado diferencias en su hidrología, fauna y flora con respecto a las del trópico asiático y africano. En Suramérica, los estudios se han concentrado en Brasil (Oliveira 1988; Roldán, 1992) y estudios sobre gradientes (Baptista et al., 2001) y evaluación de la biodiversidad (Galdean et al., 2001). Aunque los estudios realizados han sido en diferentes lugares de Brasil se concentran en ecología y limnología de la región amazónica. Venezuela, por su parte, ha realizado estudios sobre el flujo de energía, químicos y estructura de comunidades, aunque su mayor aporte se centra en el río Orinoco (revisado por Roldán, 1992). Otros países como Perú y Ecuador han realizado estudios sobre fitoplancton en lagos (Roldán, 1992), y sobre cambios altitudinales para macroinvertebrados (Jacobsen, 2003), respectivamente.
En Colombia, los estudios demoraron en comenzar, siendo el primer reporte en los años 50, sobre mediciones fisicoquímicas del agua y observaciones del plancton por Molano. Posteriormente, se realizaron una serie de estudios sobre calidad de agua en los alrededores de Bogotá y Medellín, productividad de las ciénagas del país y del río Magdalena y fauna de macroinvertebrados en Antioquia, así como de manejo de embalses (Roldán, 1992). En los últimos años se ha enfatizado en la bioindicación de la calidad del agua (Posada et al., 2000; Roldán, 2003), en las comunidades de plancton (RojasGil, 2003) y en las comunidades de macroinvertebrados (Laython, 2003; Ospina et al., 2004). En el presente estudio se pretende caracterizar la comunidad de macroinvertebrados de la quebrada Paloblanco del Santuario de Fauna y Flora Otún Quimbaya, realizando la comparación de dos puntos en dicha quebrada: un punto alto y un punto bajo, ambos con características de ríos de primer orden. Mediante la determinación de las familias de macroinvertebrados presentes en muestras generales, muestras definidas por UF y muestras de deriva se buscó constatar que las poblaciones de macroinvertebrados se sitúan y se relevan en la quebrada de acuerdo con la disponibilidad de luz, alimento y espacio, tal como lo describe el concepto del continuo de los ríos. En el caso de este estudio, el concepto de zonación se aprecia de una manera limitada ya que el sitio de muestreo es un río de montaña y sin mucha diferencia altitudinal.
MATERIALES Y MÉTODOS
El sitio de muestreo fue la quebrada Paloblanco, situada dentro del Santuario de Flora y Fauna Otún Quimbaya, ubicado en el flanco occidental de la cordillera Central, que junto con el PRN Ucumarí hace parte de la zona de amortiguación del PNN Los Nevados. Este santuario tiene una extensión de 410,88 ha, dentro de la zona mediaalta de la cuenca del río Otún en su vertiente izquierda, del departamento del Risaralda, municipio de Pereira (vereda La Suiza), con un rango altitudinal entre 1.778 y 2.270 msnm (Rico et al., 2005). La geomorfología se origina especialmente por la cuenca del río Otún, donde en uno de sus costados se localiza el santuario. Los suelos corresponden al dominio Podogenénetico Andino, cuya estructuración se ha dado por la participación de materiales volcánicos, piroplásticos, y aquellos producto de la ablación y la erosión por glaciares de materiales de la franja subandina. Los suelos de la zona son conocidos según la clasificación francesa como Andisoles. Para la zona de estudio el régimen de distribución de lluvias es de tipo bimodaltetraestacional, con dos períodos de lluvias, el primero desde abril hasta mayo, y el segundo desde octubre hasta noviembre, durante el cual cae la mayor cantidad de lluvias. Los meses más secos son febrero y julio y los meses más húmedos son abril y octubre. Los promedios mensual y anual de precipitación son de 250,2 mm y 3.075 mm, respectivamente. La temperatura presenta un comportamiento isotérmico (media mensual entre 14,4 y 15,3 ºC). La vegetación es característica de un bosque muy húmedo montano bajo, existiendo un predomino de las familias de plantas con hábito herbáceo, epífito o arbustivo. En el interior del Santuario, la totalidad de los recursos naturales son aprovechados para efectos de conservación. El recurso hídrico de la cuenca del río Otún es usado para el suministro del acueducto municipal de la ciudad de Pereira, así mismo algunas quebradas y afluentes del Otún, surten agua a los pobladores de la vereda La Suiza (Rico et al., 2005).
Se realizaron dos muestreos, uno en la desembocadura de la quebrada en el río Otún (punto bajo), el día 2 de marzo de 2005, y otro un punto alto, cercano a la cabecera de la quebrada, el día 3 de marzo de 2005, ambos en las horas de la mañana y de la tarde. Para los dos puntos se realizaron los siguientes muestreos: (1) fauna general, con una red rectangular de 1 m x 3 m, con un poro de 1 mm, que se ponía de un lado a otro del cauce durante 5 min, en los cuales se lavaban piedras grandes y se removía el sustrato, para incrementar la efectividad del muestreo. La longitud efectiva de la malla en el punto alto era de 1,5 m y en el punto bajo era de 3 m aproximadamente;
(2) muestras de fauna por grupos funcionales: hojarasca, grava, musgo y epiliton, cada una con tres réplicas, que se tomaron mediante el lavado y raspado de material de acuerdo a la unidad funcional, recogido en redes redondas con un poro de 250 µm; (3) muestras de material de deriva que se tomaron con mallas redondas y cuadradas que eran parcialmente sumergidas en partes de la quebrada que tuvieran una corriente considerable. Las redes redondas se usaron para analizar la materia orgánica a la deriva (materia orgánica gruesaMOG), y las redes cuadradas se usaron para analizar la fauna que se encontraba a la deriva. De cada una se realizaron tres réplicas. (4) Variables fisicoquímicas: pH, temperatura, contenido de oxígeno y conductividad, que fueron tomadas con un multiparámetro (Schott Handylab Multi12 /SET). Además, para el punto bajo se realizó un mapeo de las unidades funcionales: epiliton, hojarasca, musgo, grava arena y sedimento, con una cuadrícula de 1 m2, dividida cada 10 cm, con la que se expresaba la cobertura de cada unidad funcional en un área de 7 m2; también se tomó una muestra de agua (300 mL) para realizar el cálculo de materia orgánica fina (MOF) en este punto. Todas las muestras de MOG, MOF, fauna general y grupos funcionales, fueron fijadas con alcohol al 70% para su posterior manipulación y análisis en los laboratorios de Invertebrados, Ecología y Limnología, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá. Para el análisis de las variables fisicoquímicas del punto bajo, se utilizó un espectrofotómetro (HACH DR/2000 Direct Reading Spectrophotometer) para obtener los valores de fosfatos, sulfatos, nitratos y nitritos. Para hacer los cálculos de MOG, se filtraron las muestras correspondientes en un tamiz de 1 mm, se secaron estas muestras en un horno a 50 °C por 24 horas, se tomó el peso seco de la muestra en una balanza Sauter y se incineraron en una mufla MLW Electro durante tres horas. Una vez enfriado, se pesaron los restos y por diferencia se obtuvo la MOG. Para la MOF, se filtró en campo con un tamiz de 1 mm, se tomaron 200 mL de este filtrado en el laboratorio y se filtraron con un ultrafiltrador (Vacubrand Membrane Pump). Posteriormente se dejó secar el microfiltro en un horno Heraeus a 50 °C por 24 horas. Se tomó el peso seco y se siguió el procedimiento anteriormente descrito con la mufla. Para la determinación de los macroinvertebrados presentes en las muestras de fauna general, grupos funcionales y deriva, se realizó la separación manual bajo estereoscopio (Leika Galen III), la utilización de claves hasta familia y la colección en frascos con alcohol al 70% con las respectivas etiquetas. Se determinaron los hábitos de las familias presentes en la quebrada (Byers, 1984; Coffman, 1984; Edmunds, 1984; Harper y Stewart, 1984; Lange, 1984; Newson, 1984; Peterson, 1984; Polhemus, 1984; Teskey, 1984; Westfall, 1984; White et al., 1984; Wiggins, 1984; Stehr, 1991; Brusca y Brusca, 1990).
Para los cálculos de deriva se utilizó la fórmula de densidad de la población de macroinvertebrados derivantes (Ramírez y Pringle, 2001): D = Ab/t.V.A. Donde D es la densidad de la población de macroinvertebrados derivantes, Ab es la abundancia de la población, t es el tiempo de muestreo, V es la velocidad de la corriente en la boca de la red y A es el área de la boca de la red que permanece sumergida durante el muestreo. Análisis de datos Fauna general y UF: prueba de Kruskal Wallis para determinar si hay diferencias significativas en el número de individuos entre los dos puntos y por cada unidad funcional, y se determinaron los índices de diversidad alfa, Shannon y PEI (probabilidad de encuentro interespecífico) y curvas de abundancia relativa. Deriva: se calculó la desviación estándar para el resultado de MOG y MOF. Se realizó una prueba de Kruskal Wallis para los macroinvertebrados derivantes, con el fin de determinar diferencias entre los dos puntos y entre las familias en cada punto. El índice alfa fue obtenido por medio del programa Diversity (Pérez, 2005) y el índice de Shannon y PEI fue calculado por medio del software EXCEL®. Todas las pruebas estadísticas fueron corridas en el software de análisis de datos SAS V8®.
RESULTADOS
Se encontraron 42 familias de Insecta, además de Acari, Annelida, Crustacea e Hirudinea en los dos puntos muestreados (Tabla 1). De los macroinvertebrados encontrados se observa la presencia exclusiva de algunas familias dentro de cada punto muestreado.
Es así como para punto alto se encontraron 29 familias y para punto bajo 40 familias, de las que Ephydridae, Gyrinidae, Helodidae, Heteroceridae, Limnichidae, Perlidae y Tabanidae son únicas en punto alto y Caenidae, Calopterygidae, Corduliidae, Culicidae, Dytiscidae, Glossosomatidae, Hyaellidae, Libellulidae, Siphlonuridae, Veliidae, Carabidae y Curculionidae son exclusivas para punto bajo, aunque las dos últimas familias pueden proceder de ambientes terrestres y por lo tanto, no hacer parte de la comunidad acuática. Esto a gran escala muestra diferencias en la composición de las comunidades presentes en la quebrada Paloblanco.
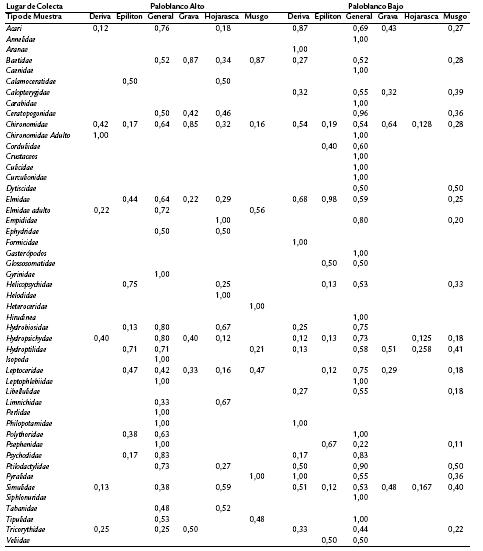
Tabla 1. Porcentaje de macroinvertebrados por familia, para las unidades funcionales y fauna general, para los sitios de muestreo.
En la tabla 2 se presentan los índices de biodiversidad de alfa, Shannon y PEI. En fauna general no se encontraron diferencias significativas (Pr > ⎯2 = 0,21) en el número de individuos entre los puntos muestreados. Para las unidades funcionales se encontraron diferencias significativas en el número de individuos para epiliton (Pr > ⎯2 = 0,036), hojarasca (Pr > ⎯2= < 0,0001) y musgo (Pr > ⎯2= < 0,0001) entre los dos puntos muestreados, mientras que para grava no hubo diferencia significativa (Pr > ⎯2 = 0,29).
| Shannon | PEI | Alfa | |
|---|---|---|---|
| Fauna General | 2,443 | 0,853 | 8,032 |
| Punto alto | 2,064 | 0,723 | 5,575 |
| Epiliton | 1,945 | 0,853 | 5,442 |
| Grava | 1,408 | 0,672 | 3,228 |
| Hojarasca | 1,524 | 0,616 | 4,090 |
| Musgo | 1,882 | 0,837 | 4,201 |
| Punto bajo | 2,249 | 0,822 | 6,871 |
| Epiliton | 1,781 | 0,807 | 2,943 |
| Grava | 1,672 | 0,828 | 4,321 |
| Hojarasca | 0,871 | 0,486 | 1,503 |
| Musgo | 1,594 | 0,980 | 3,379 |
Tabla 2. Índice de biodiversidad alfa para fauna general y las unidades funcionales para los dos sitios de muestreo (punto alto y punto bajo) en la quebrada Paloblanco.
Para las densidades de deriva de las familias de macroinvertebrados no se encontraron diferencias significativas en el número de individuos dentro de punto alto (Pr > ⎯2= 0,976), mientras que dentro de punto bajo (Pr > ⎯2= 0,023) y entre los dos puntos (Pr > ⎯2 = 0,0229), sí se encontraron diferencias. A partir de los cálculos realizados se encontraron los valores de deriva para los macroinvertebrados que se presentan en la tabla 3 y MOF, MOG en la tabla 4. Las curvas de abundancia relativa mostraron que la fauna general tiene un comportamiento geométrico (Fig. 1A), al igual que punto bajo (Fig. 1B), sin embargo, el comportamiento para punto alto es de Paloquebrado (Fig. 1C). La distribución de los hábitos de las familias presentes en cada punto se representa en la figura 2. Finalmente, se presentan los valores de las variables fisicoquímicas en la tabla 5.
DISCUSIÓN
Basados en las características geomorfológicas de los sitios de muestreo (mayor tamaño de las rocas, pendiente pronunciada, amplitud y profundidad bajas, volumen de la zona hiporreica pequeño), se han considerado ambos puntos como corrientes de cabecera (Vannote et al., 1980; Gomi et al., 2002); además, los dos puntos comparten características fisicoquímicas (oxígeno disuelto y pH) similares (Tabla 5) y presentan una diferencia altitudinal de solo 400 m. A pesar de lo anterior, se presentan en punto bajo, características que difieren de las de un río de primer orden: dosel no cerrado, menor aporte de MOG (Tabla 4) y menor conductividad (Tabla 5). Al mismo tiempo, la distribución de macroinvertebrados constituye la mayor diferencia entre los dos puntos. Al evaluar las condiciones de los sitios de muestreo se encuentran dos factores que influyen directa e indirectamente en las diferencias halladas entre los dos puntos. El primero es la deforestación de la zona aledaña al punto bajo, lo que disminuye drásticamente la vegetación riparia, así como la densidad del dosel, reduciendo de igual manera el aporte de MOG a manera de hojarasca y aumentando en gran medida la cantidad de luz que entra al río en ese punto. El segundo es el aporte antrópico de MOF a partir de residuos provenientes de las viviendas aledañas al río en el punto bajo. Los índices de diversidad para la fauna general reflejan similitud pero no igualdad, siendo ligeramente mayor el índice para punto bajo. Pese a que en fauna general no se observa una diferencia significativa, los valores de los índices de diversidad cambian entre los dos puntos de acuerdo a las UF.
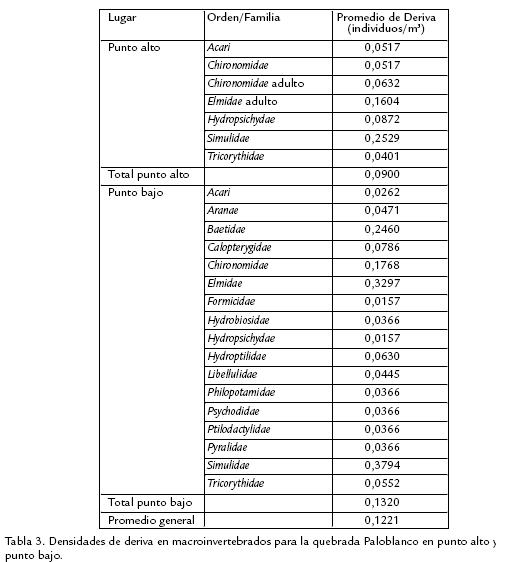
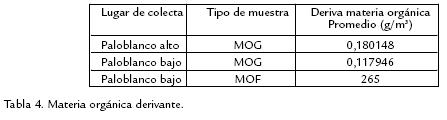
Se observa que en hojarasca hay una gran diferencia entre los dos puntos en el número de individuos; esto se puede explicar con la mayor cantidad de MOG que entra al sistema en el punto alto comparada con el punto bajo (Tabla 4). Al presentar mayor cantidad de MOG en el punto alto se facilita el establecimiento de microorganismos descomponedores, que en compañía de los macroinvertebrados inician los procesos de degradación de la misma. Ésto se contrasta con el punto bajo, en el cual se presenta un menor aporte alóctono de MOG representada fundamentalmente en la que ha sido previamente procesada en el punto alto, la cual es arrastrada con mayor facilidad por la corriente, influyendo negativamente en el establecimiento de organismos en la misma. En musgo, los índices de diversidad tienden a ser mayores en el punto alto. Esta diferencia se debe a que el número de individuos Simuliidae es muy alto en este punto, disminuyendo el valor de los índices. Sin embargo, los valores de riqueza tienden a ser mayores en el punto bajo. Teniendo en cuenta que esta UF sirve como trampa de la MOF (Anderson y Sedell, 1979) y al encontrarse mayor cantidad de MOF con respecto a la MOG (Tabla 4), hay una mayor disponibilidad de este recurso, aumentando el número de taxones y su abundancia en esta UF. Estos resultados concuerdan con otros estudios en sistemas lóticos suramericanos (Galdean, et al., 2001), en donde la mayor proporción de macroinvertebrados está asociada con agrupaciones de musgos. La diversidad en epiliton tiende a ser mayor en punto bajo ya que al aumentar la proliferación de algas debido a la mayor entrada de luz y disponibilidad de nutrientes, la oferta de recursos aprovechados por la comunidad de macroinvertebrados es mayor.
GRUPOS FUNCIONALES
Pese a determinar a nivel de familia, se genera un sesgo hacia ciertos grupos funcionales ya que se tiene incertidumbre en cuanto al hábito específico de los organismos; por lo tanto, la discusión de este aspecto podría variar en la medida que se tuviera con mayor certeza los distintos hábitos para cada uno de los organismos. El grupo funcional dominante en ambos puntos muestreados fue recolector (Fig. 2A), que en conjunto con el grupo filtrador indican que buena parte de la MO en el medio es MOF. Este resultado se contrasta con estudios similares en ríos tropicales de montaña de orden 1 (Ospina et al., 2004) en los cuales se ha reportado prevalencia de individuos recolectores y mayor cantidad de MOF. Esta particularidad puede atribuirse a que, aunque la quebrada Paloblanco es una corriente de primer orden donde se espera una mayor cantidad de MOG, las tasas de descomposición de este componente están dadas por microorganismos, pues el sistema se encuentra en una región tropical en la cual la temperatura promedio del agua es mayor en comparación con la de las regiones templadas (Irons et al., 1994). A su vez, la MOF presenta buena proporción de microorganismos que son aprovechados por los recolectores y filtradores, aumentando su abundancia, como se observa en los resultados. Se encontró una mayor cantidad de raspadores en el punto bajo (Fig. 2), debido a una mayor cantidad de algas, ya que las condiciones de elevada luminosidad, debido a una menor densidad de vegetación riparia, y mayor cantidad de nutrientes, originados en gran medida por fuentes antrópicas (aportes de aguas residuales de las viviendas aledañas a la quebrada), permiten una mayor proliferación de grupos autótrofos, en comparación con las condiciones de baja luminosidad del punto alto, que por lo tanto, disminuyen la abundancia de biopelículas, y asimismo, la cantidad de raspadores en ese punto. Esto se basa en estudios que indican una correlación positiva entre la abundancia de algas y raspadores, lo cual sugiere una dependencia por este recurso (Wallace y Webster, 1996).
| Lugar de Colecta | pH | Conductividad ( uSeals/L) | Oxígeno Disuelto (mg/L) |
|---|---|---|---|
| Paloblanco Alto | 7,22 | 37 | 7,76 |
| Paloblanco Bajo | 6,9 | 4 | 7,92 |
Tabla 5. Variables fisicoquímicas en la quebrada Paloblanco.
El porcentaje de individuos filtradores es mayor en punto bajo, siendo la familia Simuliidae la más representativa (Fig. 2), lo cual sería lo esperado al presentarse una mayor cantidad de MOF. Asimismo, la alta abundancia de este grupo funcional, está dada por su bajo gasto energético para buscar alimento, al ser organismos que emplean la energía cinética del río, de tal manera que al encontrar sus requerimientos ecológicos (suficiente velocidad de corriente, alta calidad y concentración de sestón orgánico) pueden soportar biomasas superiores por unidad de área (Wallace y Webster, 1996). Como organismos cuya función principal es la remoción y fijación de MOF (Wallace y Merrit, 1980), los filtradores aportan al medio otra fuente de materiales aprovechables para las algas e incrementan su abundancia, siendo esta, otra causa del mayor porcentaje de raspadores discutido anteriormente.
La diferencia de porcentajes de depredadores encontrados (Fig. 2) se basa en parte a la mayor cantidad de presas disponibles, en este caso en particular, una mayor cantidad de colectores en punto alto, los cuales son las presas más frecuentes de este grupo de organismos (Wallace y Webster, 1996). Otro factor que puede estar influenciando la abundancia de los depredadores es la presencia de grupos de organismos que los depreden en el punto bajo (se encontraron peces en el muestreo), a diferencia del punto alto donde la ausencia de un factor limitante como éste permite un incremento en la población de depredadores. Pese a la mayor cantidad de depredadores en punto alto, se presenta una mayor diversidad en punto bajo que, junto con las curvas de abundancia relativa (Fig. 1) indican una mayor variedad de recursos que la sustentan. Al excluir a Chironomidae del grupo de depredadores (datos no mostrados), el porcentaje cambia en gran medida, pasando a ser similar entre los dos puntos.
El mapeo que se hizo en punto bajo refleja como están distribuidas las unidades funcionales en un área de 7 m2. Se observa que la unidad predominante es epiliton, seguida de grava, emergente y musgo, hojarasca y arena en igual proporción. Aunque en el área muestreada la unidad funcional que más se encontró fue el epiliton, la cantidad de organismos en ella es mínima probablemente porque este hábitat no ofrece mayores beneficios en cuanto a retención de nutrientes y condiciones de asentamiento. En contraste, las unidades funcionales con menor cobertura presentan un gran número de organismos ya que, a diferencia del epiliton, son hábitats que facilitan el establecimiento de las comunidades de macroinvertebrados, los cuales ofrecen una fuente de retención de nutrientes y de estabilidad estructural y nutricional para los organismos. De acuerdo con las curvas de abundancia relativa (Fig. 1) se observa que en punto bajo se presentan pocas familias comunes y todas las demás son raras, mientras que en punto alto la distribución de las familias en la comunidad se adecua a la repartición de recursos, de tal manera que existe una especialización por parte de los grupos presentes en el punto (University of Vermont, 2004). Esto obedece a una restricción en los recursos alimenticios en relación con el punto bajo, cuya fuente principal es de origen alóctono y que son degradados y transportados rápidamente por la corriente, lo cual limita el número de familias que se pueden establecer allí.
DERIVA
La diferencia significativa encontrada entre ambos puntos se da porque al presentarse una mayor riqueza y abundancia en punto bajo hay mayor competencia y los organismos aumentan su movilidad en busca de recursos, aumentando la probabilidad de que un individuo sea arrastrado por la corriente. Se encontró que la familia que más deriva tuvo en los dos puntos de muestreo es Simulidae, (Tabla 3), cuyo hábito filtrador impone exposición directa a la corriente, y por su abundancia, tiene mayor probabilidad de ser arrastrada. Las siguientes familias con mayor deriva corresponden a Elmidae (adulto) y Baetidae para los puntos alto y bajo, respectivamente, ambas de hábito raspador, indicando que la movilidad permanente en busca de alimento aumenta la exposición a la corriente, favoreciendo su arrastre.
CONTINUO
Resulta difícil relacionar el presente trabajo con el concepto del continuo desde el punto de vista de los grupos funcionales de macroinvertebrados, ya que las proporciones entre raspadores, colectores, depredadores, etc., son expresadas a partir de la biomasa y en nuestro estudio se hizo a partir de abundancias. Asimismo, hay una marcada intervención antrópica en uno de los sitios de muestreo que supone cambios en la composición de las comunidades de macroinvertebrados con respecto a sistemas naturales. Teniendo en cuenta las consideraciones anteriores, este modelo pasa a un segundo plano dentro de la discusión de los resultados.
A partir de los resultados se puede afirmar que las diferencias encontradas entre los dos puntos de muestreo son causadas en gran medida por una intervención antrópica (deforestación de la rivera del río), que implica cambios en las condiciones abióticas y los proceso biológicos que alteran la composición de las comunidades de macroinvertebrados y los grupos funcionales dominantes en ríos de primer orden.
AGRADECIMIENTOS
A la Universidad Nacional de Colombia por el apoyo logístico para el desplazamiento hasta el lugar de muestreo. A la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales y al Santuario de Fauna y Flora Otún Quimbaya, por permitirnos realizar este trabajo en los terrenos del santuario y por toda la información y apoyo que recibimos durante nuestra estadía allí. A Carolina Mora, Ximena Galíndez, Camilo Loaiza, Luis Carlos Peña, Lizeth Cifuentes, Dalí Rojas, Margarita Victoria, Simón Cortés y Catalina Palacios por su esfuerzo y dedicación durante los muestreos y a Diego Mendivelso y Soraya Villalobos, además, por su ayuda en las determinaciones. Finalmente a Javier Rodríguez y al Profesor Rodulfo Ospina por su apoyo y asesoría.
BIBLIOGRAFÍA
ANDERSON NH, SEDELL JR. Detritus Processing by Bajo in Stream Ecosystems. Annu Rev Entomol. 1979;24:351357.
[ Links ]BAPTISTA DF, BUSS DF, DORVILLÉ LFM, NESSIMIAN JL. Diversity and Habitat Preference of Aquatic Insects Along the Longitudinal Gradient of the Macaé River Basin, Rio de Janeiro, Brazil. Rev Bras Biol. 2001;61(2):249258.
[ Links ]BRUSCA R, BRUSCA G. Invertebrates. Sunderland (MA): Sinauer Associates; 1990.
[ Links ]BYERS GW. Tipulidae. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 491–514.
COFFMAN, WP, FERRINGTON LC. Chironomidae. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal /Hunt; 1984. p. 551–652.
EDMUNDS GF. Ephemeroptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 94–125.
GALDEAN N, CALLISTO M, BARBOSA FAR. Biodiversity Assessment of Benthic Bajo in Altitudinal Lotic Ecosystems of Serra do Cipó (MG, Brazil). Rev Bras Biol. 2001;61(2):239248.
[ Links ]GOMI T, SIDLE RC, RICHARDSON JS. Understanding Processes and Downstream Linkages of Headwater Systems. Bioscience. 2002;52(10):905916.
[ Links ]HARPER PP, STEWART KW. Plecoptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 182–230.
HYNES HBN. Zonation of the Invertebrate Fauna in a West Indian Stream. Hydrobiologia. 1971;38:18.
[ Links ]ILLIES J, BOTOSANEANU L. Problemas et methodes de la classification et de la zonation ecologicque des eaux courantes, considerees surtout du point du vue faunistique. Mitt Int Ver Theor Angew Limnol. 1963;12:157.
[ Links ]IRONS JG III, OSWOOD MW, STOUT RJ, PRINGLE CM. Latitudinal Patterns in Leaf Breakdown: Is Temperature Really Important? Freshw Biol. 1994;32:401411.
[ Links ]JACOBSEN D. Altitudinal Changes in Diversity of Bajo From Small Streams in the Ecuadorian Andes. Arch Hydrobiol. 2003;158(2):145167.
[ Links ]LANGE WH. Aquatic and Semiaquatic Lepidoptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal /Hunt; 1984. p. 348–360.
LAYTHON M. Caracterización de macroinvertebrados acuáticos en los ríos Arzobispo y San Cristóbal (Cerros Orientales, Bogotá/Colombia), en la misma franja altitudinal y con énfasis en quironómidos [trabajo de grado]. Bogotá: Universidad Nacional de Colombia; 2003.
[ Links ]LOCK MA, WALLACE RR, COSTERTON JW, VENTUYO RM, CHARLTON SE. River Epilithon: Toward a StructuralFunctional Model. Oikos. 1984;42:1022.
[ Links ]NEWSON, HD. Culicidae. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 515–533.
OSPINA R, RISS W, GUTIÉRREZ J, MURCIA P, ÁLVAREZ J. Proyecto tipología de pequeños ríos en Cundinamarca y la validez de los conceptos ecológicos actuales en los ríos tropicales de montaña [informe técnico final]. Bogotá (Colombia): ColcienciasUniversidad Nacional de Colombia; 2004.
OLIVEIRA L. Introduçao ao Estudo da limnología 1944, The Amazon Limnology and Landscape Ecology of a Might Tropical River and Its Basin 1984, trabajos de lagunas brasileñas 19151983, II Congreso Brasileño de limnología; 1988
[ Links ]PÉREZ J. Entomología Granadina. 2005 [citado 31 Oct 2005]. Disponible en URL: http://perso.wanadoo.es/jp-l/descargas.html.
[ Links ]PETERSON BV. Simulidae. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 534–550.
POLHEMUS JT. Aquatic and Semiaquatic Hemiptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 231–260.
POSADA JA, ROLDAN G, RAMÍREZ J. Caracterización fisicoquímica y biológica de la calidad de aguas de la cuenca de la quebrada Piedras Blancas, Antioquia, Colombia. Rev BiolTrop. 2000;48(1):5970.
[ Links ]RAMÍREZ A, PRINGLE C. Spatial and Temporal Patterns of Invertebrate Drift in Streams Draining a Neotropical Landscape. Freshw Biol. 2001;46:4762.
[ Links ]RICO V, PINZÓN A, MEDINA J editores. Estudios ecológicos regionales del Santuario de Fauna y Flora Otún Quimbaya Risaralda, Colombia. Bogotá (Colombia): Universidad Nacional de Colombia; 2005.
[ Links ]ROJASGIL YC. Comparación de la estructura de la comunidad de fitoplancton en varios lagos y ríos de la cuenca del Putumayo y Caquetá (Amazonía colombiana) [trabajo de grado]. Bogotá: Universidad Nacional de Colombia; 2003.
[ Links ]ROLDÁN, GA. Fundamentos de limnología neotropical. Medellín (Antioquia): Universidad de Antioquia; 1992.
[ Links ]ROLDÁN GA. Bioindicación de la calidad del agua en Colombia. Uso del método BMWP/Col. Medellín (Antioquia): Universidad de Antioquia; 2003.
[ Links ]SABATER SY, ROMANI AM. Structural and Activity of Rock and Sand Biofilm in a Mediterranean Stream. Ecology. 2001;82(11):32323245.
[ Links ]STEHR, FW. Immature Insects. Dubuque (IA): Kendal/Hunt; 1991.
[ Links ]TESKEY HJ. Part One. Larvae of Aquatic Diptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal /Hunt; 1984. p. 448–466.
UNIVERSITY OF VERMONT (US). Diversity of Stream Organisms. 2004. [citado 31 de Oct 2005] Disponible en URL: http://www.uvm.edu/~biology/Classes/102/PDF/ Streams.pdf.
[ Links ]VANNOTE RL, MINSHALL GW, CUMMINS KW, SEDELL JR, CUSHING CE. The River Continuum Concept. Aquat Sci. 1980;37:130137.
[ Links ]WALLACE JB, WEBSTER JR. The Role of Bajo in Stream Ecosystem Function. Annu Rev Entomol. 1996;41:115139.
[ Links ]WALLACE JB, MERRITT RW. FilterFeeding Ecology of Aquatic Insects. Annu Rev Entomol. 1980;25:103132.
[ Links ]WESTFALL MJ. Odonata. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 12676.
[ Links ]WHITE DS, BRIGHAM WU, DOYEN JT. Aquatic Coleoptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 361437.
[ Links ]WIGGINS GB. Trichoptera. In: Merrit RW, Cummins, KW. An Introduction to the Aquatic Insects of North America. 2 ed. Iowa: Kendal/Hunt; 1984. p. 271–311.