










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.11 no.2 Bogotá June 2006
OBSERVACIÓN DE RANGOS DE VUELO DE Bombus Atratus (Hymenoptera: Apidae) EN AMBIENTES URBANOS
Observation of Flight Ranges of Bombus Atratus (Hymenoptera: Apidae) in Urban Environments
LAÍN PARDO, LUDWIG JIMÉNEZ Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá. Colombia.
Presentado agosto 23 de 2005, aceptado abril 3 de 2006, correcciones abril 20 de 2006
RESUMENSe estudió la capacidad de regreso de Bombus atratus a su colonia midiendo la cantidad de individuos que volvieron a ésta después de ser liberadas a diferentes distancias y en cuatro direcciones (norte, sur, este, oeste). Para ello se trasladó una colonia de B. atratus, proveniente de Tenjo Cundinamarca, al Departamento de Biología, Universidad Nacional de Colombia Sede Bogotá, se marcaron y liberaron un total de 100 forrajeras de las cuales regresaron 40. Hubo una relación lineal negativa clara entre la proporción de regresos al nido y las distancias del sitio de liberación, con reducción del número de abejorros capaces de regresar a medida que aumentaba la distancia al nido. El rango máximo observado al cual las abejas pudieron regresan al nido está entre 1.300m y 1.500m y un análisis de regresión lineal predice un rango de vuelo de 1,6 km.
Palabras clave: rango de vuelo, orientación, Bombus atratus.
ABSTRACTThe return capacity of Bombus atratus to its colony was studied by measuring the quantity of individuals that returned to it, after being released at different distances and in four directions (north, south, east, west). We located a colony of B. atratus coming from Tenjo, Cundinamarca, at the Department of Biology, Universidad Nacional de Colombia, Bogotá. We marked and released a total of 100 workers of which 40 returned. There was a clear negative relationship between the proportion of bees returning to its nest and the distance from the released site, decreasing the number of bumblebees able to return as it increased the distance to the nest. The observed maximum range to which the bees found their nest was between 1,300 m and 1,500 m and a lineal regression analysis predicts a flight range of 1.6 km.
Key words: flight range, orientation, Bombus atratus.
INTRODUCCIÓNLos animales han desarrollado mecanismos de orientación que aseguran su regreso a casa después de desplazarse en busca de alimento. En Bombus trasversalis por ejemplo se ha demostrado el uso de rastros en el terreno (Cameron y Witfield, 1996); así como en B. impatients, el uso de pistas próximas y lejanas para el reconocimiento de la entrada (Plowright et al., 1995) o de algún sistema de aprendizaje por asociación en sus preferencias florales respecto al tamaño de éstas (Blarer et al., 2002). Así como orientación geomagnética o planos de luz polarizada en aves y abejas, entre otras (Michener, 1974). Estos mecanismos sin embargo funcionan en un espacio determinado o a una distancia lejos del nido. Esta distancia máxima a la cual los insectos pueden regresar a su casa se le conoce como rango de vuelo o comportamiento de homing y varía de acuerdo a la especie (Goulson y Scout, 2001). En Melipona sp. por ejemplo la máxima distancia registrada es de 2,1 km (Roubik y Aluja, 1983), para Bombus terrestris; Goulson y Stout (2001) reportaron un rango de vuelo hasta de 9,8 km. En Apis mellifera se conoce una distancia nidoindividuo de 13,5 km (Michener, 1974), sin embargo la distancia mayor registrada desde la cual un himenóptero puede regresar a su colonia es de 23 km, para una abeja euglosina (Euplusia surinamensis; Janzen, 1971). Para B. atratus, así como para otras especies suramericanas, las investigaciones sobre su rango de vuelo son casi inexistentes. Sin embargo, hay trabajos que abordan otros aspectos de su biología como el tiempo de forrajeo, el cual varía un promedio de 30 minutos como en Apis, pero se conoce que algunas obreras pueden quedarse por fuera todo el día e incluso la noche (Michener, 1974). Tanto en Bombus spp. como en Apis spp., tiende a haber una constancia en la actividad que realizan en los viajes (Free, 1955), así por ejemplo si salen a colectar néctar harán varios viajes en busca de lo mismo solamente. Se sabe que frecuentan la misma vecindad durante varias visitas, pero al parecer tienen algún mecanismo que les permite reconocer las flores visitadas con el fin de no gastar energía en una flor ya usada, como lo son el reconocimiento de pistas en el terreno y los arreglos florales (Redmon y Plowright, 1996). En este estudio se hicieron pruebas para determinar el rango de vuelo de esta especie de Bombus, las cuales constituyen el primer registro y se convierte en un punto de partida para estudios de comportamiento e historia natural, aspectos poco conocidos en el país.
MATERIALES Y MÉTODOSSe trasladó una colonia de abejorros (Bombus atratus) de Tenjo, Cundinamarca, al laboratorio de Etología, Departamento de Biología, Universidad Nacional de Colombia, que estaba compuesta por aproximadamente 150 individuos con seis reinas y no se observaban machos. Una vez en el laboratorio se acondicionó una caja de icopor y se depositaron dentro de ella. La caja estaba regulada térmicamente por medio de un termostato a una temperatura constante de 28 ºC y comunicada con el exterior a través de un tubo plástico de 4 cm de diámetro que atravesaba una ventana para que los animales salieran constantemente. Antes de la salida se diseñó una recamara para facilitar la captura y marcaje de los animales que entraban y salían. Se permitió más de una semana para que la colonia se familiarizara con su nuevo entorno antes de comenzar con el registro de datos. Para la toma de datos se hizo una modificación de la metodología empleada por Goulson y Stout (2001). El muestreo duró aproximadamente 20 días, y se realizó en días más o menos soleados o secos para facilitar el desplazamiento de los animales. Se capturaban 20 abejorros forrajeros que entraban a la colonia con polen, a los cuales se les pegaba en la superficie dorsal del tórax una lámina plástica, circular y numerada de 2 mm de diámetro y un peso de 0,0012 g. Estos 20 abejorros se distribuyeron equitativamente en cuatro contenedores o cajas negras, fueron llevados en diferentes direcciones (norte, sur, este, oeste) y a una distancia determinada para luego ser liberados, comenzando a 250 m de la colonia. Este procedimiento se repitió aumentando paulatinamente la distancia cada 250 m ó 500 m y terminando en aquella en la cual no se registraron retornos a la colonia. Teniendo así, 20 abejorros en total por cada punto o distancia de liberación, repartidos de a cinco individuos en cada punto cardinal. Se emplearon cajas negras con el fin de evaluar la capacidad de retorno de los abejorros, al limitar algunos de los mecanismos de orientación. Los abejorros se alimentaron antes de ser liberadas con una solución azucarada, por medio de un dispensador instalado en las cajas de transporte para que tuvieran suficiente energía y se liberaron en jardines con flores, para que pudieran forrajear. Luego de la liberación se hicieron observaciones diurnas del nido para registrar las abejas marcadas que lograron regresar al nido.
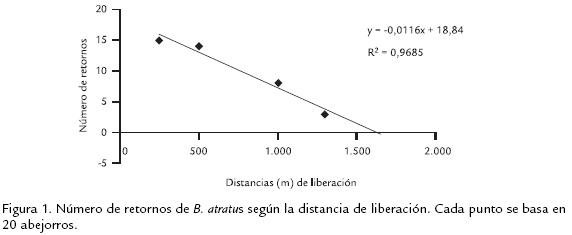
RESULTADOSSe liberaron un total de 100 forrajeras de las cuales regresaron a la colonia 40 de ellas (Tabla 1). La distancia a la cual las abejas de este ensayo no tuvieron la capacidad de ubicar su nido fue 1.500 m. Aunque se quiso evaluar una distancia entre 1.300 m y 1.500 m, no se hizo un ensayo intermedio entre estas distancias, porque el nido no pudo reactivarse luego de la fase normal de declive y no hubo suficientes individuos para hacer otra liberación. Sin embargo, basados en el número absoluto de retornos por cada distancia de liberación, se estableció por medio de una regresión lineal (y = 18,843 0,0116x) (r2 = 0,96, valor crítico de F = 0,01) la distancia máxima a la cual ya no se esperarían retornos, que sería el rango de vuelo teórico, esto es a 1,6 km aproximadamente. Para esta regresión no se consideró la distancia a la cual no llegó ninguna abeja (1.500 m) ya que produciría sesgos en la línea de tendencia (Roubik y Aluja, 1983), porque al incluir más puntos fuera del rango de vuelo encontrado se distorsiona la proporción o la relación de la distancia con el número de retornos (Fig. 1).
| Distancia (m) | Norte | Sur | Este | Oeste | Total/20 |
|---|---|---|---|---|---|
| 250 | 5 | 5 | 3 | 2 | 15 |
| 500 | 3 | 3 | 4 | 4 | 14 |
| 1.000 | 2 | 2 | 2 | 2 | 8 |
| 1.300 | 0 | 2 | 1 | 0 | 3 |
| 1.500 | 0 | 0 | 0 | 0 | 0 |
Tabla 1. Total de retornos de obreras de B. atratus según la orientación de liberación y la distancia.
Con el fin de establecer si había una relación entre el número de abejas que lograban retornar y la dirección a las cuales se liberaban, se realizó una prueba estadística no paramétrica de Friedman (Zar, 1999) que permitió demostrar que no hay diferencias significativas, en otras palabras la capacidad de retorno no está influenciada por la dirección de liberación, (xr2 = 0,082, p = 0,99).
DISCUSIÓNEl rango de vuelo encontrado está dentro de lo descrito para muchas especies en Europa y es alrededor de los 500 m (Goulson y Stout, 2001). Sin embargo, los mismos autores revisan otras especies donde su rango de acción puede ser tan amplio como el encontrado por ellos para Bombus terrestris de 9,8 km. Esto demuestra la gran habilidad de los abejorros para recorrer distancias. Por otro lado, si bien el dato de rango de vuelo ofrecido en este experimento no debe ser confundido con el home range o rango de acción, éste es un buen indicativo que podría aproximar su verdadero rango de acción por dos razones. Primero, se sabe que la probabilidad de regreso de una abeja más allá de su rango de acción es muy limitado, 0,17 cuando esta distancia es 1,2 veces el rango de acción (Roubik y Aluja, 1983). Segundo, teniendo en cuenta que los mecanismos enunciados en general tienen la condición de generar marcas o rastros por los lugares que familiarmente recorren los animales, nos hace pensar que poner a las abejas transportándolas en cajas negras fuera de su rango de acción es lo que hace que pierdan la facultad de orientación para volver a la colonia, es decir las distancias a las cuales las abejas lograron llegar pueden haber sido distancias que ya habían recorrido antes, con las cuales estaban familiarizadas o bien que estaban muy cerca de estas. También se ha registrado que algunos insectos hacen búsquedas sistemáticas de sus rastros para luego ubicar su nido, sin embargo ninguno de estos mecanismos se han investigado en Bombus. El hecho de transportarlos en cajas negras impidiendo la selección de marcas visuales a la largo del camino hace pensar que debe haber algún tipo de mecanismo quizás olfativo o los anteriormente mencionados que permita trazar su ruta de regreso a determinada distancia más allá de su rango de acción. Aunque se trató de observar la dirección que tomaron una vez son liberados, no fue posible porque cuando se les abría la caja de transporte, salían realizando trayectorias circulares al mismo tiempo que ascendían sobre ella y luego parecían alejarse en espiral para tomar una dirección. Este comportamiento también lo registraron Goulson y Stout (2001), para Bombus terrestris, y parece ser necesario para ubicarse en el nuevo sitio desconocido y activar sus procesos de reconocimiento y orientación, que desafortunadamente no están del todo claros. Se pudo establecer, sin embargo, que cada una de las cinco abejas liberadas en cada orientación tomaban rutas distintas, aunque en otras ocasiones varias o todas tomaban una determinada dirección. Se encontró que el tiempo mínimo de regreso desde 250 m, corresponde a 1,5 minutos, lo que sugiere una velocidad aproximada de 10 km/h, la cual consideramos una buena aproximación ya que con este tiempo, se supone que su camino de regreso fue prácticamente directo y continuo, en vista que al haber sido alimentadas previamente no tendrían que detenerse para reabastecerse de néctar o polen. Hay que tener en cuenta que el estimativo del número de retornos está determinado por varios factores no relacionados con la capacidad de llegar al nido. Es decir muchos de los animales liberados pudieron simplemente ser objeto de presa por parte de algún depredador, también pudieron haber bajas por cambios intempestivos de la condiciones climáticas, que puede ser un factor que reduzca la frecuencia de regresos al nido. Así mismo, cabe la posibilidad de que la marca circular se hubiera desprendido. Esto sugiere que no solo se debe suponer que las que no retornaron al nido, no tuvieron la habilidad de orientarse. En lo que respecta a la dirección con la que fueron liberados los grupos de abejorros, no fue encontrada una relación entre el número de retornos y la dirección (x2 = 0,082, p = 0,99), aunque lo hubiéramos esperado como en Melipona fasciata donde se han reportado relaciones entre el número de retornos y la orientación con la que se liberan, especialmente a distancias intermedias (Roubik y Aluja, 1983). Esto puede mostrar que las abejas usan varias formas de orientación y no hay ventajas aparentes por una ruta o por la otra.
CONCLUSIONESEste estudio logró determinar que la distancia máxima desde la cual pueden regresar los abejorros a su nido es de 1,6 km, con la cual se pudo inferir su posible rango de acción, para las condiciones en que se realizaron las pruebas. Éste es un punto de partida que permite indagar varios aspectos sobre su ecología e investigar, entre otras cosas, los mecanismos que permiten a esta especie encontrar su camino de regreso. Sugerimos como paso siguiente realizar un ensayo transportándolas en cajas transparentes para establecer si las claves visuales son importantes para esta especie, así como aumentar el número de colonias y realizarlo en condiciones de vida silvestre, incorporando a los análisis un estudio de las fuentes de alimento presentes.
AGRADECIMIENTOSLos autores agradecen muy especialmente a Wolfgan Hoffman, por su colaboración y enseñanzas sobre el tema. También a los profesores Rodulfo Ospina, German Amat y Enrique Zerda por su valioso apoyo.
BIBLIOGRAFÍABLARER A, KEASAR T, SHMIDA. Possible Mechanisms for the Formation of Flower Size Preferences by Foraging Bumblebees. Ethology. 2002;108(4):341351.
[ Links ]CAMERON S, WHITFIELD J. Use of Walking Trails by Bees. Nature. 1996;379:125.
[ Links ]FREE J. The Collection of Food by Bumble Bees. Insectes Sociaux. 1955;2:303311.
[ Links ]GOULSON D, STOUT J. Homing Ability of the Bumblebee Bombus Terrestris (Hymenoptera: Apidae). Apidologie. 2001;32:105111.
[ Links ]JANZEN D. Euglossine Bees as Long Distance Pollinators of Tropical Plants. Science. 1971;171:203205.
[ Links ]MICHENER D. The Social Behavior of the Bees, a Comparative Study. Cambridge, Massachusset: Belknap Press of Harvard University Press.; 1974.
[ Links ]PLOWRIGHT C, O’CONNELL E, ROBERTS L, REID S. The Use of Proximal and Distal Cues in Nest Entrance Recognition by Bumble Bees. J. Apic Res. 1995;34(2):5764.
REDMON D, PLOWRIGHT C. Flower Revisiting by Foraging Bumble Bees: the Effect of Landmarks and Floral Arrangement. J Apic Res. 1996;35(3/4):96103.
[ Links ]ROUBIK D, ALUJA M. Flight Ranges of Melipona an Trigona in Tropical Forest. J Kans Entomol Soc. 1983;56(2):217222.
[ Links ]ZAR J. Biostatistical Analysis. Englewood Cliffs: PrenticeHall. N.S.; 1974.
[ Links ]
