










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.12 no.1 Bogotá Jan./June 2007
DIVERSIDAD DE ABEJAS (HYMENOPTERA: APOIDEA) EN ESTADOS SUCESIONALES DEL BOSQUE HÚMEDO TROPICAL
Bee Diversity (Hymenoptera: Apoidea) in a Tropical Rainforest Succession
ALLAN SMITH PARDO1, M. Sc., Ph. D.; VICTOR H. GONZALEZ2, B.Sc., Candidato Ph.D. 1Entomología, Departamento de Ciencias Agronómicas, Universidad Nacional de Colombia, Medellín, Colombia. ahsmithp@unal.edu.co 2Department of Ecology and Evolutionary Biology, Haworth Hall, 1200 Sunnyside Avenue, University of Kansas, Lawrence, Kansas 660457523, USA. vhgonza@ku.edu
Presentado 20 de noviembre de 2006, aceptado 13 de diciembre de 2006, correcciones 21 de enero de 2007.
RESUMEN
Los objetivos principales de este trabajo fueron entender cómo la diversidad de abejas cambia durante la sucesión en un bosque húmedo neotropical, y comparar la eficiencia de los cuatro métodos de captura usados. La diversidad y riqueza de abejas fueron estudiadas al nororiente de Medellín (Antioquia, Colombia; 9001.100 m) durante seis meses en cuatro estados sucesionales (pastizal, rastrojo bajo, rastrojo alto y bosque). Las abejas fueron intensiva y sistemáticamente capturadas usando los siguientes métodos: red entomológica, Malaise, metil salicilato para abejas euglosinas (Apidae, Euglossini) y trampa Van Somer con pescado de agua dulce en descomposición. En total se capturaron 15.356 especímenes pertenecientes a cuatro familias y 287 especies de abejas. Esto corresponde al 62% del número total de especies de abejas conocidas para Colombia. Aproximadamente el 50% de todos los individuos fueron abejas sociales sin aguijón (Apidae, Meliponini). Trigona (Trigona) fulviventris fue la especie más abundante (~10%) en todo el muestreo. Augochlora y Megachile fueron los géneros con mayor número de especies. El pastizal y rastrojo alto presentaron los mayores índices de diversidad y riqueza de abejas y fueron significativamente más altos que los del bosque y rastrojo bajo. En todos los estados sucesionales, excepto por el bosque, el número de especies nuevas por muestra tendió a cero, y las curvas de acumulación de especies tendieron a normalizarse con el tiempo y aumento en el área de muestreo. La red entomológica fue el método más eficiente en todos los estados sucesionales, excepto en el bosque, donde el mayor número de especies e individuos fueron capturados con la trampa Van Somer. Sin embargo, en todos los estados sucesionales un mayor porcentaje (50%) de especies raras fueron colectadas con Malaise. El número de especies nuevas colectadas en cada muestra y las curvas de acumulación de especies sugieren que el muestreo fue suficiente para estimar la diversidad de abejas en estados sucesionales tempranos, pero insuficiente para el estudio de la apifauna del bosque. La trampa Van Somer fue un método eficiente para capturar abejas en el bosque y debería ser incluida en muestreos regulares de abejas en bosques tropicales. También se resumen los inventarios de abejas realizados en Colombia y se resalta la importancia de usar otros métodos de muestreos menos comunes para estudiar las abejas en ecosistemas tropicales.
Palabras clave: Apoidea, sucesión, inventarios, Colombia.
ABSTRACT
The purposes of this work were to study the bee diversity across successional stages in a Neotropical rainforest and to test four sampling methods. Bee diversity and richness were studied in northeastern Medellín (Antioquia, Colombia; 9001100 m in altitude) over six months using a chronosequence with four successional stages (abandoned pastures, low shrubs, secondary forest, and mature forest). Bees were intensively and systematically sampled, using the following methods: entomological net, Malaise trap, Methyl Salicylate, and Van Somer trap with decomposed fresh water fish. A total of 15.356 specimens were collected, belonging to four families and 287 species, representing 62% of all bee species found in Colombia. About 50% of all individuals sampled were stingless social bees (Apidae, Meliponini). Trigona (Trigona) fulviventris was the most abundant species (~10%) in the survey. Augochlora and Megachile were the most specious genera. The pasture and secondary forest showed high values of diversity and richness and were significantly higher than those of the mature forest and low shrubs. In all successional stages, except in the mature forest, the number of new species collected in each sample period approached zero and the species accumulation curves tended to stabilize as time and sampling area increased. The net was the most efficient method in all successional stages, except in the forest, where most bee species and individuals were collected with the Van Somer trap. However, a higher percentage (50%) of rare species was collected with the Malaise trap. The number of new species collected in each sampled period and the species accumulation curves suggest that our survey was nearly sufficient to estimate the bee diversity in these early successional stages, but insufficient to study the mature forest apifauna. Due to the high efficiency of the Van Somer trap to attract bees in the forest, this trap should be used regularly in additional bee surveys in tropical rain forests. We also summarize the bee surveys in Colombia and highlight the importance of using other less common sampling methods to study bees from tropical ecosystems.
Key words: Apoidea, succession, surveys, Colombia.
INTRODUCCIÓN
Los objetivos principales de este trabajo fueron entender cómo la diversidad de abejas (Hymenoptera: Apoidea) cambia durante la sucesión en un bosque húmedo tropical y comparar la eficiencia de los métodos de captura usados. Las abejas son uno de los grupos de animales más importantes en la polinización de las plantas y por lo tanto, organismos críticos en la recuperación y mantenimiento de las comunidades vegetales en muchos ecosistemas (Huffaker y Rabb, 1984; Neff y Simpsom, 1993; Michener, 2000). Dado que la riqueza y diversidad estructural de las plantas en un determinado ecosistema aumentan con la edad de la sucesión, esperamos encontrar una mayor diversidad y riqueza de abejas en los estados sucesionales tardíos, ya que ellos ofrecen una mayor diversidad de habitats y recursos (Murdoch et al., 1972; Southwood et al., 1979; BarberenaArias y Aide, 2002). También discutimos brevemente otros métodos de captura, los inventarios de abejas hasta ahora realizados en Colombia y la importancia de los inventarios sistemáticos e intensivos para entender mejor la verdadera diversidad de abejas en las áreas tropicales.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
El trabajo se realizó en un bosque húmedo tropical del área de influencia del proyecto hidroeléctrico Porce II, localizado al nororiente del departamento de Antioquia, en los municipios de Yolombó, Amalfi y Gómez Plata [6°46’N,75°06’W], aproximadamente a 120 km de Medellín. El área de estudio presenta una altitud de 9001.100 m, con una temperatura promedio anual de 22,5 °C, humedad relativa media mensual del 83% y precipitación media anual de 3.050 mm. El régimen de lluvias es bimodal, con un periodo seco durante cinco meses desde fines de noviembre hasta marzo y un período lluvioso de abril a octubre.
INVENTARIO
El inventario de abejas fue hecho en cuatro tipos de sucesión vegetal: pastizal (P), rastrojo bajo (RB), rastrojo alto (RA) y bosque (B). En total se ubicaron tres parcelas representativas de cada estado sucesional. Las coordenadas geográficas y una descripción detallada de la vegetación de cada parcela se encuentran en Pérez y Gutiérrez (1996) y SmithPardo (1999a). En cada parcela se establecieron dos transectos de 100 m de longitud por 2,5 m de ancho para un total de seis transectos por estado sucesional y 24 transectos que representan un área total de 6.000 m2. La recolección de abejas fue realizada durante cuatro días a la semana entre las 8:00 y las 16:00 horas. Cada semana se muestrearon cuatro parcelas de un estado sucesional diferente con el fin de cubrir todas las parcelas de todos los estados sucesionales cada mes. Los muestreos se realizaron en un total de 60 días (15 días por estado sucesional), entre los meses de junio y diciembre de 1997 (720 horas en total; 360 horas por recolector). Las abejas fueron capturadas usando cuatro métodos diferentes: red entomológica, Malaise, trampa Van Somer (TVsp) y trampa para abejas euglosinas (Apidae, Euglossini; TE). Para TVsp se usaron peces enteros de agua dulce en descomposición. El pescado o partes de éste fueron dejadas al aire libre hasta su descomposición, durante 45 días en bolsas plásticas transparentes, herméticamente selladas. La trampa TE es específica para la captura de abejas euglosinas (Roubik, 1989; Roubik y Hanson, 2004). Para TE se usó metil salicilato porque es un atrayente ampliamente usado en la captura de estas abejas. Además, fue la esencia sintética con mayor frecuencia de captura en un muestreo preliminar en el área de estudio (SmithPardo, 1999b). El metil salicilato fue puesto en recipientes plásticos con orificios, cuyos fondos contenían una solución de agua y detergente al 50%. Por cada transecto se usaron dos trampas de cada tipo durante seis horas cada día. Para las recolecciones con la red entomológica, cada transecto fue recorrido durante todo el período de muestreo por dos recolectores. Las abejas fueron recolectadas sobre flores o volando cerca de ellas. En lo posible, se recolectaron abejas con la misma intensidad en todas las flores en un área determinada. Cada dos horas, todos los especímenes recolectados fueron transferidos a viales plásticos. Las abejas fueron sacrificadas y conservadas en acetato de etilo o en alcohol al 70%. Las muestras se preservaron a temperaturas bajo cero en el laboratorio hasta su montaje en seco. Todo el material entomológico recolectado, una colección de referencia de las especies de abejas, y una base de datos electrónica que reúne la información de captura y plantas visitadas para cada individuo, se encuentran en el Museo de Entomología Francisco Luis Gallego, Universidad Nacional de Colombia, Sede Medellín.
ANÁLISIS ESTADÍSTICO
Para efectos de comparación, en todos los análisis los especímenes capturados de abejas solitarias, parásitas y eusociales (Apidae: Meliponini, Bombini y Apini) fueron considerados como individuos. Usando el programa Visual Foxpro, versión 5.0, en un computador personal se calcularon las frecuencias relativas (constancia) y absolutas de las especies, así como los siguientes índices de riqueza, diversidad y similitud: Margalef (IM), Shannon (IS), B. Parker (BP), Jaccard, Sörensen y Bray Curtis. Las especies con frecuencias relativas menores del 2% fueron consideradas especies raras. Para los análisis estadísticos se utilizó el programa estadístico Statgraphics, versión 3 Plus para Windows. La prueba de Tstudent fue usada para comparar los diferentes índices de diversidad estimados y establecer si existían diferencias estadísticas significativas entre ellos (Magurran, 1988).
RESULTADOS
INVENTARIO
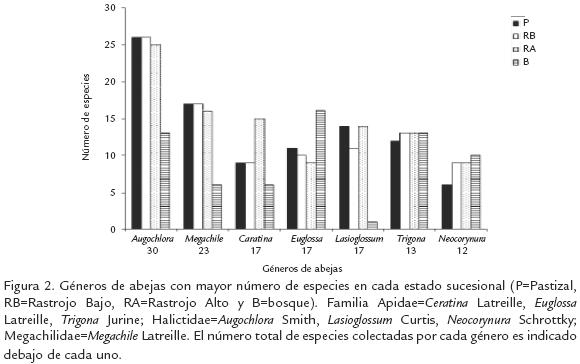
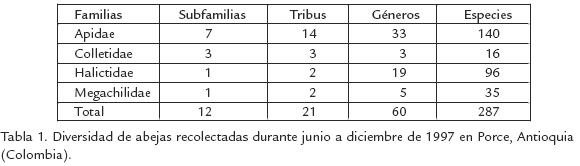
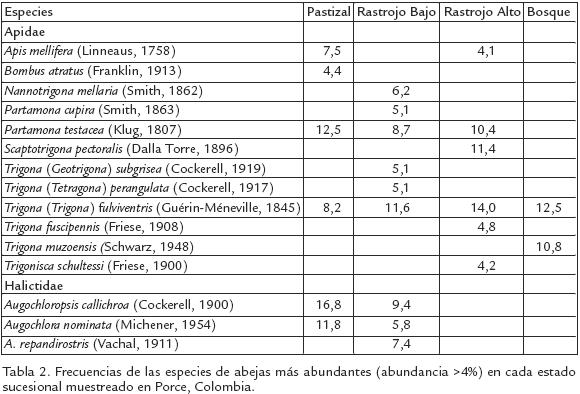
Se capturaron 15.356 especímenes de cuatro de las cinco familias de abejas conocidas para Colombia (Tabla 1). Una lista completa de las especies se encuentra en SmithPardo (1999a). Solamente el 47% de las especies pudieron ser identificadas al nivel de especie, las demás fueron tratadas como morfoespecies. Aproximadamente el 50% de todos los individuos capturados fueron abejas sociales sin aguijón (Meliponini). Junto con Halictidae, ambos grupos representaron cerca del 82% del total de individuos capturados. Las siguientes especies fueron las más abundantes durante el estudio (Tabla 2): Trigona (Trigona) fulviventris (10%), Partamona testacea (5,6%), P. cupira (4%), Augochlora nominata (3,4%) y Augochloropsis callichroa (3,5%). Apis mellifera, la abeja introducida, tuvo una abundancia del 2,6%. Solamente 49 especies (17%) estuvieron representadas por un solo individuo. Augochlora (Halictidae) y Megachile (Megachilidae) fueron los géneros con mayor número de especies (Fig. 2)
ACTIVIDAD DIURNA
La mayor cantidad de abejas fue recolectada entre las 10 y 12 horas, aunque no existieron diferencias estadísticamente significativas entre la cantidad de individuos capturados temprano en la mañana (810 hr) o en la tarde (1416 hr; Tstudent, p > 0,05). No obstante, excluyendo las abejas sociales sin aguijón, la mayoría de individuos fueron capturados en las horas de la tarde.
SUCESIÓN VEGETAL
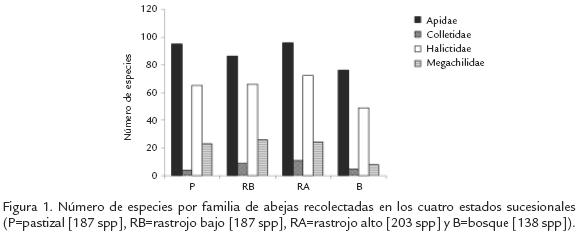
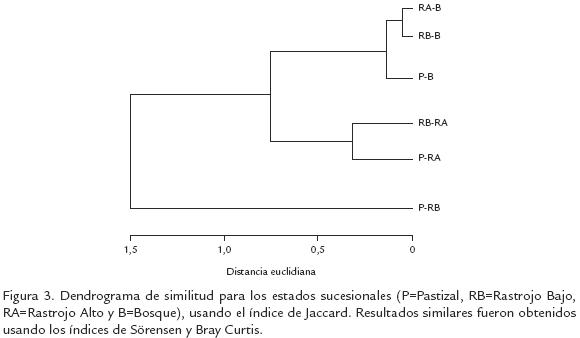
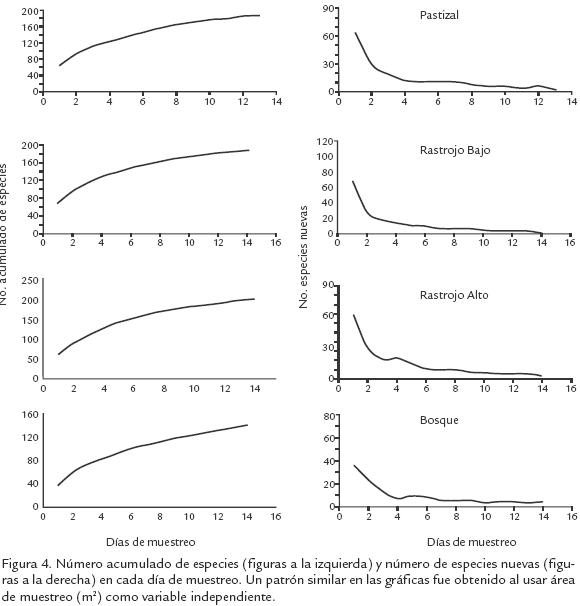
La mayoría de las especies (70%) fueron recolectadas en RA (Fig. 1). El Pastizal y RA presentaron los mayores índices de diversidad y riqueza de abejas (IM=22,8 y 24,2; IS=4,0 y 4,1, respectivamente) y no fueron estadísticamente diferentes entre ellos (Tstudent, p>0,001); sin embargo, estos valores fueron significativamente mayores que los valores estimados para B y RB (IM=17,6, IS=3,8; Tstudent, p<0,09). El Pastizal y RA también presentaron valores bajos de dominancia (BP=0,09). El mayor número de especies exclusivas se encontró en B (28 especies), seguido por P (25), RA (23) y RB (12). En general, las fases iniciales de la vegetación (incluyendo el RA) presentaron mayor similitud, que estos con B (Fig. 3). Las curvas de acumulación de especies en todos los tipos de vegetación, excepto en B, tendieron a normalizarse con el tiempo y aumento en el área de muestreo; además, el número de especies nuevas por muestra tendió a cero (Fig. 4).
EFICIENCIA DE LOS MÉTODOS
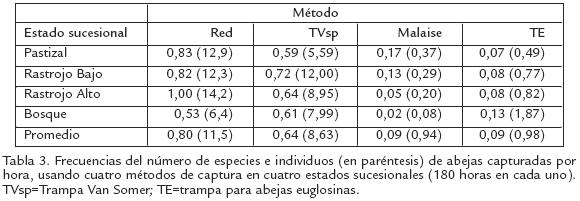
Se encontraron diferencias estadísticamente significativas entre el número de individuos /hora y especies/hora capturados entre los diferentes métodos de muestreo y el estado sucesional (Tabla 3; Tstudent, p<0,001). Más del 80% del número total de especies y el mayor número de especies exclusivas en cada estado de sucesión fueron capturadas con la red entomológica, seguida por TVsp (IM=26,8 y 22,2; IS=4,2 y 3,9, respectivamente). Alrededor del 94% del total de individuos y cerca del 90% de las especies fueron capturadas combinando ambos métodos. La red entomológica fue el método más eficiente en los estados sucesionales, excepto en el bosque donde el mayor número de especies (80%) e individuos fueron capturados con TVsp. Sin embargo, en todos los estados sucesionales un mayor porcentaje (50%) de especies raras fueron recolectadas con Malaise, la mayoría de ellas representadas por un solo individuo. No se encontraron diferencias estadísticamente significativas en la diversidad de abejas capturadas con la red entomológica entre RA y RB, pero sí entre éstos y los demás estados de sucesión (Tstudent, p0,001). Para TVsp se encontraron diferencias significativas entre los valores de diversidad en: RBP, RARB, RAB y PB. No se encontraron diferencias significativas entre la diversidad de abejas capturadas con Malaise y TE, ni entre los tipos de vegetación (T student, p>0,001).
DISCUSIÓN
DIVERSIDAD DE ABEJAS Y SUCESIÓN
La alta diversidad de las familias Apidae y Halictidae (Fig. 1), al igual que los géneros con mayor número de especies encontrados en nuestro estudio (Fig. 2), concuerda con lo observado en otros bosques neotropicales de tierras bajas (p.e., Cure et al., 1992) e incluso en ecosistemas altoandinos tropicales. Al menos en los trópicos, ambas familias son numerosas, abundantes, de hábitos generalistas, contienen especies que pueden ser estacionales, y presentan diversos grados de sociabilidad que van desde solitarias hasta altamente sociales. Las abejas sin aguijón son un buen ejemplo de especies altamente sociales, presentes todo el año y restringidas a las zonas tropicales del mundo (Michener, 1979; Michener, 2000). Los géneros Augochlora, Megachile, Lasioglossum y Neocorynura no solo tienen muchas especies, la gran mayoría de ellas sin describir, sino que también son ecológicamente muy diversos y están ampliamente distribuidos en la región neotropical. Lasioglossum y Neocorynura también son los grupos con mayor número de especies en bosques de niebla y páramos en Colombia (Gonzalez y Engel, 2004).
El bosque, a pesar de tener una alta diversidad y riqueza estructural, no presentó la mayor diversidad y riqueza de abejas. Esto sugeriría que no existe una relación directa entre la riqueza de abejas y la edad de la sucesión. Resultados similares han sido registrados para otros grupos de insectos en bosques húmedos tropicales (BarberenaArias y Aide, 2002). Sin embargo, esta baja diversidad y riqueza de abejas en el bosque estudiado puede ser el resultado de sesgos en los métodos de captura y no una evidencia de un estado sucesional pobre en abejas. Por ejemplo, en espacios abiertos, con vegetación baja y muchas plantas en floración, tales como rastrojos y pastizales, son lugares favorables para el recolector común. De hecho, en estos estados de sucesión la red entomológica fue el método más eficiente en número de individuos y especies recolectados por hora (Tabla 3). En el bosque, los muestreos con la red entomológica estuvieron restringidos al sotobosque (2 m de altura sobre el suelo), donde existen muy pocas plantas en floración. La mayoría de las especies del bosque fueron capturadas con TVsp, una trampa que nunca ha sido usada en muestreos intensivos de abejas. Quizás, el fuerte olor y los fluidos producidos durante la descomposición del pescado son atrayentes fuertes para las abejas y otros insectos, principalmente aquellos que forrajean en estratos superiores del bosque. De hecho, algunas observaciones sugieren que la efectividad de TVsp se incrementa con un aumento en el grado de descomposición del pescado, generalmente alcanzando un máximo entre los seis y ocho días.
El sesgo en los métodos de captura usados en el bosque también es apoyado por las curvas de especies acumuladas y el número de especies nuevas por muestra (Fig. 4). A diferencia de otros estados de sucesión, estas curvas no tendieron a normalizarse con el tiempo, ni con un aumento en el área de muestreo en el bosque. Esto indica que el muestreo realizado fue suficiente para tener una idea de la fauna de abejas en estados sucesionales tempranos pero no en el bosque. Otros métodos de recolección como nidostrampa (Krombein, 1967), platos de colores o trampas de luz ultravioleta (Borror et al., 1989; Leong y Thorp, 1999) pueden ser usados para capturar abejas en bosques tropicales. El uso de tales métodos puede incrementar el número de especies raras y traer resultados interesantes aún en áreas que han sido intensivamente estudiadas. Por ejemplo, usando nidostrampa en el dosel del bosque, Thiele (2002) recolectó Duckeanthidium thielei Michener (Megachilidae), un género previamente conocido de Suramérica. De igual forma, existen varios géneros de abejas en varias familias [Andrenidae (Oxaea Klug), Colletidae (Ptiloglossa Smith) y Halictidae (Megalopta Smith)] que forrajean temprano en la mañana o en las últimas horas de la tarde, cuando la luz es escasa. Estas abejas son fácilmente recolectadas en grandes números usando trampas de luz y por esta razón estuvieron poco o no representadas en nuestro estudio. Al igual que en el bosque, la red entomológica tampoco es el mejor método para recolectar abejas en ecosistemas altoandinos (bosques de niebla o páramos). A tales alturas, las bajas temperaturas, alta radiación y cambios repentinos en las condiciones del clima son factores limitantes para el forrajeo de las abejas. Estas condiciones restringen los tiempos de forrajeo de las abejas a períodos cortos durante el día cuando las condiciones son favorables. Por ejemplo, varias especies de abejas [p.e. Chilicola Spinola (Colletidae), Neocorynura (Halictidae)] raramente son recolectadas usando redes o incluso trampas Malaise, pero pueden ser encontradas fácilmente cuando se buscan sus nidos en el suelo o en ramas secas de las plantas (Gonzalez y Michener, 2004; Gonzalez y Engel, 2004; Gonzalez, 2006). En conjunto, estos ejemplos muestran que para estudiar la diversidad de abejas en ecosistemas tropicales se requiere, además de los métodos tradicionales (p.e. red entomológica, Malaise), la combinación de varios métodos de muestreo. La trampa TVsp fue un método eficiente para capturar abejas en el bosque y debería ser incluída en muestreos regulares de abejas en bosques tropicales.
INVENTARIOS DE ABEJAS EN COLOMBIA
Nuestro estudio es el primer inventario intensivo y sistemático de abejas realizado hasta el momento en Colombia (Tabla 4) y cualquier otro país en Suramérica tropical, excepto Brasil (para Brasil ver: Cure et al., 1992; Carvalho y Bego, 1996; PinheiroMachado et al., 2002). Los pocos inventarios que existen en Colombia están restringidos a bosques tropicales del noroccidente del país. Esto indica que la mayoría de zonas de vida y regiones naturales todavía están por explorar. A pesar de esto, Colombia posee una de las faunas de abejas mejor conocida entre todos los países andinos (NatesParra y Gonzalez, 2000; SmithPardo, 2003). Por ejemplo, Venezuela, Ecuador o Perú carecen completamente de listas de especies. Sin embargo, la mayor parte de este conocimiento en Colombia proviene de registros de literatura e identificación de material recolectado esporádicamente (SmithPardo 2003; Gonzalez y Engel 2004). Esto hace que comparaciones de la diversidad de abejas con otras partes del mundo sean difíciles de realizar, especialmente por las diferencias en los procedimientos e intensidad de la recolección.
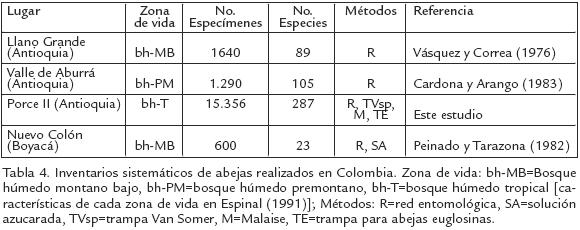
Alrededor del 62% de los géneros y especies actualmente conocidos para Colombia (SmithPardo, 2003) fueron recolectados en este trabajo. Este número de especies es considerablemente alto teniendo en cuenta que provienen de un área relativamente pequeña y de una misma zona de vida (bosque húmedo tropical). Además, a partir de este material se han registrado por primera vez para el país géneros y especies anteriormente conocidos de otras partes de Suramérica, y se han descrito varias especies nuevas y muchas otras están por describir (p.e. Michener, 2002; SmithPardo, 2003; Gonzalez y Ospina, 2006).
DIVERSIDAD DE ABEJAS TROPICALES
A diferencia de otros grupos de organismos, la mayor abundancia y diversidad de abejas en el mundo se encuentra aparentemente en las regiones xéricas de las áreas templadas y no en los trópicos (Michener, 1979; Michener, 2000). Una de las hipótesis para explicar este patrón de distribución es la competencia desproporcionada de las abejas sociales sin aguijón por los recursos disponibles. Las abejas sin aguijón están restringidas a las regiones tropicales y subtropicales del mundo, son muy abundantes y altamente generalistas. Además, tienen sistemas de reclutamiento avanzados que permiten una rápida adquisición de recursos, desplazando así, a otras abejas. Otra de las hipótesis es que los ambientes húmedos de los trópicos pueden afectar la supervivencia de las crías de las abejas, facilitando el crecimiento de hongos y otros microorganismos en las reservas de alimentos, especialmente en las abejas que nidifican en el suelo. Este hecho explicaría por qué los grupos de abejas más exitosos y abundantes en los trópicos (p.e. abejas sin aguijón) nidifican principalmente en la madera. Otra buena explicación es que aún desconocemos en gran medida la verdadera diversidad de los insectos tropicales (Roubik, 1989; Michener, 2000; Gonzalez, 2006).
En resumen, el número de especies de abejas encontrados en este trabajo no solo muestra una alta diversidad de abejas en el área de estudio, sino también que inventarios intensivos y sistemáticos pueden darnos una mejor idea de la verdadera diversidad en los trópicos. Muestreos intensivos de abejas en áreas tropicales son escasos y los pocos existentes demuestran una alta diversidad comparable o superior con las áreas templadas. Por ejemplo, Costa Rica tiene 16 y 32 veces más especies de abejas por unidad de área que México y Estados Unidos, respectivamente (Griswold et al., 1995).
AGRADECIMIENTOSA las Empresas Públicas de Medellín por su apoyo económico y logístico; G. Morales y C. Aguilar, por su asesoría durante el desarrollo de este trabajo, T. Griswold por su ayuda en la identificación de las abejas; M. Camacho, E. Palacios, D. Bennett y dos evaluadores anónimos por sus comentarios y sugerencias que ayudaron a mejorar este manuscrito. A todo el personal del postgrado en Entomología de la Universidad Nacional de Colombia, Sede Medellín, especialmente a J. Abril y J. Quiroz. Apoyo financiero para VG fue obtenido a través de los fondos generales de investigación de la Universidad de Kansas y el proyecto USIsrael Binational Science Foundation grant 2000259 (D. Smith y Lubin).
BIBLIOGRAFÍA
BARBERENAARIAS MF, AIDE TM. Variation in Species and Trophic Composition of Insect communities in Puerto Rico. Biotropica. 2002;34(3):357367.
[ Links ]BORROR DJ, TRIPLEHORN CA, JONSON NF. An Introduction to the Study of Insects. 6th Ed. Saunders College Publishing, Philadelphia, PA; 1989.
[ Links ]CARDONA J, ARANGO CI. Inventario de la fauna Apoidea (Insecta: Hymenoptera) del Valle de Aburrá y sus relaciones con la flora [tesis de pregrado]. Medellín: Departamento de Biología, Universidad de Antioquia; 1983.
[ Links ]CARVALHO AD, BEGO L. Studies on Apoidea fauna of Cerrado Vegetation at the Panga Ecological Reserve, Uberlandia, M. G., Brasil. Rev Bras Entomol. 1996;40(2):147156.
[ Links ]CURE JR, THIENGO M, SILVEIRA FA, ROCHA LB. Levantamento da fauna de abelhas silvestres na zona da mata de Minas Gerais IIIMata secundária na região de Viçosa (HymenopteraApoidea). Rev Bras Entomol. 1992;9(3/4):223239.
[ Links ]ESPINAL LS. Apuntes ecológicos. Universidad Nacional de Colombia, Seccional Medellín; 1991.
[ Links ]GONZALEZ VH, MICHENER CD. A new Chilicola from Colombian Páramo (Hymenoptera, Colletidae, Xeromelissinae). J Hym Res. 2004;13(1):2430.
[ Links ]GONZALEZ VH. Superfamilia Apoidea. En: Fernández F, Sharkey M, editores. Introducción a los Hymenoptera de la región neotropical. Sociedad Colombiana de Entomología y Universidad Nacional de Colombia, Bogotá. 2006. p. 443448.
[ Links ]GONZALEZ VH, ENGEL MS. The Tropical Andean Bee Fauna (Insecta: Hymenoptera: Apoidea), with Examples from Colombia. Entomol Abh (Dres). 2004;62(1):6575.
[ Links ]GONZALEZ VH, OSPINA M. Nuevos registros genéricos de abejas (Hymenoptera: Apoidea) para Colombia. Acta biol Colomb. 2006;11:9394.
[ Links ]GRISWOLD T, PARKER FD, HANSON PE. The Bees (Apidae). En: Hanson PE, Gauld ID, Editors. The Hymenoptera of Costa Rica. Oxford University Press, Oxford, UK. 1995. p. 650691.
[ Links ]HUFFAKER CB, RABB RL. Ecological Entomology. John Wiley and Sons, Nueva York, Nueva York; 1984.
[ Links ]KROMBEIN KV. TrapNesting Wasps and Bees: Life Histories, Nests, and Associates. Smithsonian Press, Washington; 1967.
[ Links ]LEONG JM, THORP RW. ColourCoded Sampling: The Pan Trap Colour Preferentes of Oligolectic and Nonoligolectic Bees Associated with a Vernal Pool Plant Ecol Entomol. 1999;24:329335.
[ Links ]MAGURRAN AR. Ecological Diversity and Its Measurement. New Jersey, Princenton University Press; 1988.
[ Links ]MICHENER CD. Biogeography of the Bees. Ann Mo Bot Gard. 1979;66:277347.
[ Links ]MICHENER CD. The Bees of the World. Johns Hopkins University Press; Baltimore, MD; 2000.
[ Links ]MICHENER CD. The Bee Genus Chilicola in the Tropical Andes, with Observations on Nesting Biology and a Phylogenetic Analysis of the Subgenera (Hymenoptera, Colletidae, Xeromelissinae). Nat Hist Mus Univ Kansas. 2002;26:147.
[ Links ]MURDOCH WW, EVANS FC, PETERSON CH. Diversity and Pattern in Plants and Insects. Ecology. 1972;53: 819828.
[ Links ]NATESPARRA G, GONZALEZ VH. Las abejas silvestres de Colombia: por qué y cómo conservarlas. Acta biol Colomb. 2000;5(1):537.
[ Links ]NEFF JL, SIMPSON BB. Bees, Pollination Systems and Plant Diversity. En: LaSalle, J, Gauld ID, Editors. Hymenoptera and Biodiversity. C.A.B International, Oxon, UK. 1993. p. 143168.
[ Links ]PÉREZ CM, GUTIÉRREZ V. Estudio de la vegetación en cuatro estados sucesionales en las zonas aledañas a la hidroeléctrica Porce II [tesis de pregrado]. Medellín: Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia; 1996.
[ Links ]PEINADO JE, TARAZONA A. Reconocimiento preliminar de la flora apícola y su interacción con la fauna apoidea en la región de Nuevo Colón (Boyacá) [tesis de pregrado]. Tunja: Universidad Pedagógica y Tecnológica de Colombia; 1982.
[ Links ]PINHEIROMACHADO C, SANTOS I, IMPERATRIZFONSECA VL, PEIXOTO AM, SILVEIRA FM. Brazilian Bee Surveys: State of Knowledge, Conservation and Sustainable Use. En: Kevan P, ImperatrizFonseca VL, Editors. Pollinating Bees. The Conservation Link Between Agriculture and Nature. Ministry of Environment, Brasilia, Brazil. 2002. p. 115129.
[ Links ]ROUBIK DW. Ecology and Natural History of Tropical Bees. University Press, Cambridge, Nueva York. 1989.
[ Links ]ROUBIK DW, HANSON PE. Abejas de orquídeas de la América tropical. Biología y guía de campo. Instituto Nacional de Biodiversidad, Santo Domingo de Heredia, Costa Rica. 2004.
[ Links ]SMITHPARDO AH. Abejas (Hymenoptera: Apoidea) de la zona de influencia del embalse Porce II (Antioquia, Colombia) [tesis de Maestría]. Medellín: Facultad de Ciencias Agropecuarias, Postgrado en Entomología, Universidad Nacional de Colombia; 1999a.
[ Links ]SMITHPARDO AH. Evaluación de cinco métodos de muestreo para abejas en dos estados sucesionales del área de influencia del embalse Porce II (Antioquia). Rev Fac Nal Agr. 1999b;52(1):435450.
[ Links ]SMITHPARDO AH. A Preliminary Account of the Bees of Colombia (Hymenoptera: Apoidea): Present Knowledge and Future Directions. J Kansas Entomol Soc. 2003;76(2):335341.
[ Links ]SOUTHWOOD TRE, BROWN VK, READER PM. The Relationships of Plant and Insect Diversities in Succession. Biol J Linn Soc. 1979;12:327348.
[ Links ]THIELE R. Nesting Biology and Seasonality of Duckeanthidium thielei Michener (Hymenoptera: Megachilidae), an Oligolectic Rainforest bee. J Kansas Entomol Soc. 2002;75(4):274282.
[ Links ]VÁSQUEZ AJ, CORREA A. Estudio sobre la fauna Apoidea y sus relaciones con la flora y el medio ambiente en la región de Llano Grande (Río Negro, Antioquia) [tesis de pregrado]. Medellín: Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia; 1976.
[ Links ]
