










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.12 no.2 Bogotá Jul./Dec. 2007
COMUNIDAD DE MARIPOSAS DIURNAS (LEPIDOPTERA:RHOPALOCERA) EN UN GRADIENTE ALTITUDINAL DEL CAÑON DEL RÍO COMBEIMATOLIMA, COLOMBIA
Butterflies Community (Lepidoptera:Rhopalocera) Along an Altitudinal Gradient in Combeima River Canyon Tolima, Colombia
ÉDGAR CAMERO1, Biólogo, M.Sc.; ANDERSON M. CALDERÓN C.2, Biólogo. 1Autor para correspondencia. Departamento de Biología, Universidad Nacional de Colombia. Sede Bogotá, ciudad universitaria, Carrera 30 No. 4503. AA. 14490. Bogotá, Colombia. Teléfono: 571 316 50 00, ext. 11322. Fax: 571 316 53 10. eecameror@unal.edu.co 2Unidad de Conservación y Biodiversidad Biológica, Ministerio de Ambiente y Recursos Naturales de Venezuela. Av. Marginal del Torbes San Cristóbal, Venezuela.
Presentado 2 de noviembre de 2006, aceptado 26 de abril 2007, correcciones 28 de mayo de 2007.
RESUMEN
Se estudió la variación altitudinal de la composición, riqueza, diversidad y estructura de la comunidad de mariposas diurnas en un gradiente altitudinal en la cuenca del río Combeima (JuntasTolima, Colombia), desde los 1.800 m hasta los 3.000 m a lo largo del año en tres periódos de muestreo, mediante el uso de redes entomológicas y trampas VanSomerenRydon. Se encontraron 101 especies de mariposas distribuidas en 59 géneros y cinco familias cuyos parámetros poblacionales variaron espacial y temporalmente, mostrando marcada zonificación altitudinal en los 1.900 m la cual es una franja transicional de la composición faunística y en la que a partir de ella, existe un decremento en el número de especies exclusivas para cada estación de muestreo, las cuales podrían considerarse como especies indicadoras de disturbio ecosistémico.
Palabras clave: ecología, bioindicadores, insectos, Lepidoptera, diversidad, gradientes.
ABSTRACT
Altitudinal variations in composition, richness, diversity and structure were studied in the butterflies’ communities along an altitudinal gradient in Combeima River basin (JuntasTolima, Colombia) from 1800 m until 3000 m during one year through three sampling times, using insect nets and VanSomerenRydon traps. 101 butterflies species were collected that were classified into 59 genera and five families. Butterflies population values changed through altitude and year season, and showed important ecological changes in 1900 m where we found a transition zone in fauna composition and where exclusive fauna decrease in each sampling station. This exclusive fauna could be named as indicator of ecological disturbances.
Key words: Ecology, bioindicators, insects, Lepidoptera, diversity, gradients.
INTRODUCCIÓN
GENERALIDADES
La biodiversidad es uno de los temas centrales de la biología (MartínPiera, 1998); esta es el resultado de los diferentes procesos evolutivos y ecológicos y abarca toda la escala de organización de los seres vivos (Escurra, 1990; Halfter, 1998). La diversidad biológica representa más que el número de especies y su abundancia en un tiempo y un lugar; también posee una dimensión ecológica funcional que se refiere a los procesos e interrelaciones al interior de las poblaciones, así como de éstas en la comunidad y el ecosistema (MartínPiera, 1998). La valoración de la biodiversidad requiere de cualquier forma y en determinado grado, un conocimiento de estos componentes. Las comunidades biológicas se definen como un conjunto de poblaciones de diferentes especies que coexisten e interactúan en el tiempo y en el espacio (Magurran, 1989), y funcionan en virtud de una compleja red de interacciones. Estas comunidades se pueden estudiar y describir a partir de tres componentes: la composición, la distribución espaciotemporal o estructura y la función; la evaluación de estos componentes para una comunidad determinada no resulta redundante sino complementaria. La descripción de las comunidades en términos de composición se realiza a través del número de especies en un lugar determinado es decir, el inventario. Por otro lado, la estructura, la organización física o los patrones de un sistema se deben estudiar teniendo en cuenta que las comunidades biológicas poseen un conjunto de atributos que no residen en cada una de las poblaciones que las componen, sino que se manifiestan en la comunidad (Krebs, 1985) y que corresponden a las propiedades colectivas y emergentes de las poblaciones. El estudio de éstas propiedades es básico para el estudio de la biodiversidad de cualquier taxón, pero de manera especial para los insectos, ya que estas poblaciones varían considerablemente en sus atributos espaciotemporales, debido principalmente a su corto ciclo generacional y la gran adaptación que presentan en los diferentes ecosistemas (Fuentes, 2004). Los estudios de diversidad de insectos estiman que la riqueza del grupo se encuentra entre los 5 y 30 millones de especies en todo el mundo (Erwin, 1982; Gaston, 1991; Hodkinson, 1992; Stork, 1993), y a pesar de no existir una aproximación más concreta de la cifra total de especies del grupo, se estima que cerca de la tercera parte de los insectos del mundo se encuentra en el neotrópico, especialmente en Perú, Colombia y Brasil, razón atribuida a que en esta zona del planeta existe gran variedad de ecosistemas gracias a diferencias topográficas que favorecen el aislamiento de las poblaciones y aumentan el grado de endemismo de las especies (Brown, 1991; Camero, 1999). En el territorio colombiano se conocen cerca de 3.019 especies de mariposas que representan el 61,9% de las mariposas conocidas para el neotrópico. Los estudios acerca de la conservación de sus comunidades en las distintas zonas altitudinales de Colombia son importantes especialmente en zonas por encima de los 2.200 m de altitud, las cuales están desapareciendo debido especialmente al aumento de las áreas agrícolas y la introducción de especies vegetales; por lo cual, la conservación de las especies de mariposas, a menudo depende de la protección de áreas adyacentes más bajas, donde en ocasiones se presenta la mayor abundancia de sus poblaciones (Andrade, 2002).
ECOLOGÍA DE GRADIENTES
La composición y estructura de las comunidades en un ecosistema presenta variaciones a lo largo de gradientes altitudinales. El aumento de la altitud en ecosistemas naturales se refleja en cambios en la composición natural de las especies de artrópodos (Janzen, 1993). En cuanto a la estructura, existen modelos con capacidad de predicción de los patrones de distribución de las especies a lo largo de gradientes altitudinales (Terborgh, 1971; Camero, 2003), los cuales están basados principalmente en la interacción de las especies con variables físicas como temperatura y humedad, así como con variables biológicas como disponibilidad de recursos, coexistencia o discontinuidades en el hábitat. Las mariposas presentan una alta especificidad hacia las plantas de las cuales se alimentan y están estratificadas en cuanto a gradientes de luz, viento, humedad y temperatura, razón por la que constituyen un grupo de trabajo muy sensible a las variaciones climáticas y ecológicas que se presentan en un gradiente altitudinal. Los estudios en gradientes con comunidades de mariposas, muestran que la diversidad disminuye hacia las zonas de mayor altitud a la vez que aumenta el porcentaje de exclusividad y endemismo; además, la tendencia general de la riqueza es a disminuir con el aumento altitudinal del gradiente, aunque cada familia muestra una tasa independiente de disminución de la riqueza (Fagua, 1999; Andrade, 2002). La distribución de las especies es mayor entre los 600 y los 1.400 m, con una fuerte reducción de la riqueza entre los 1500 y los 1.800 m; franja que representa un área de transición entre las comunidades de tierras bajas y comunidades típicas de montaña.
INSECTOS COMO BIOINDICADORES
En los ecosistemas tropicales donde las poblaciones animales se ven más seriamente amenazadas a causa del vertiginoso avance de la deforestación y de la fragmentación, se genera la extinción de gran cantidad de recursos naturales regionales incluyendo especies de insectos aún no descritas y posiblemente de gran importancia ecológica y económica (Gordon, 1985). Los estudios en fauna de insectos aportan información sobre el estado de los ecosistemas, de su productividad y de los niveles de contaminación acuática y atmosférica, ya que interacciones como intercambios genéticos, biomasa y energía transferidas en los ecosistemas se encuentran directamente relacionadas con las poblaciones de insectos (Brown, 1991). De otro lado, los insectos tienen un papel preponderante en la conservación debido a que se desempeñan como factores formadores y reguladores de los ecosistemas (Camero, 1999), establecen relaciones que se caracterizan por ser cercanas y a menudo muy precisas con la biota en general y poseen suficientes atributos que permiten considerarlos indicadores adecuados de la calidad de hábitats (Brown, 1991; Sutton y Collins, 1991; Pearson y Cassola, 1992; Andrade, 1998; Keer et al., 2000), y son a la vez, herramientas clave en monitoreos que midan el impacto de los disturbios y de la fragmentación (Didham et al., 1996; Didham et al., 1998; Dewenter y Teja, 2000; Hoyle y Harbone, 2005), lo cual permite identificar áreas importantes en políticas de conservación y de manejo (Kremen, 1992; Fattorini, 2006). El presente estudio busca conocer la estructura espacial y temporal de la comunidad de mariposas diurnas en un gradiente altitudinal del cañón del río Combeima (Tolima) a partir de parámetros poblacionales y de diversidad, a fin de establecer herramientas biológicas que puedan utilizarse como indicadoras de perturbación en este tipo de ecosistemas y permitan ser utilizadas en posteriores estudios de conservación.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
La zona de estudio comprende un gradiente altitudinal entre los 1.800 y los 3.000 m pertenecientes a la Reserva Natural Ibanasca, en la cuenca del río Combeima, departamento del Tolima, Colombia. Esta cuenca se localiza en el costado oriental de la Cordillera Central (Hernández, 1991) y corresponde a una zona de amortiguación del Parque Nacional Natural de los Nevados (Fig. 1).
Esta cuenca está caracterizada por tres grandes formaciones geológicas correspondientes a depósitos volcánicos, esquistos metamórficos y cristalinos (Ospina et al., 1979). De acuerdo con la clasificación de la cuenca realizada por Leonel y Páez (2000), el área de estudio corresponde a una unidad climática fríohúmeda, con relieve montañoso de pendientes que oscilan entre el 30 y el 70%, y que en general está conformada en un 95% por suelos del tipo Typic Melanudans. De acuerdo con el sistema de Holdridge (1996), se trata de un Bosque Andino Montano Bajo correspondiente a una Región de Vida Andina y Subandina (Rangel y Garzón, 1987; Rangel et al., 1997), con temperaturas que pueden oscilar entre los 12 y 19 ºC, y precipitación que varía entre los 1.500 a 4.000 mm anuales. En las zonas más altas, la composición florística está dominada por Chusquea scandens, Passiflora capsularis, Passiflora mixta, Bomarea caldasii, Clusia lineata, Clusia grandiflora, Bocconia frutescens y más de 100 especies de orquídeas encontradas especialmente en las zonas boscosas (Esquivel y Nieto, 2003). En la zona, las plantaciones de especies foráneas como pinos y eucaliptos, junto con la ganadería bovina, han dado paso a ecosistemas de reemplazo caracterizados por matrices de Pennisetum clandestinum y Poa annua, que han desplazado y fragmentado gran parte del bosque nativo generando un mosaico del paisaje en el cual los relictos de bosque se encuentran principalmente en las partes más altas y de mayor pendiente.
COLECCIÓN Y DETERMINACIÓN TAXONÓMICA
La metodología empleada para la captura de las especies de lepidópteros corresponde a una adaptación de las aplicadas por Sparrow y Ehrlich (1993), De Vries et al. (1997), Andrade (1998), Fagua (1999) y De Vries y Walla (2001). Se efectuaron tres salidas de campo de 12 días entre junio de 2005 y marzo de 2006: la primera, se realizó en mitad de la época seca, la segunda a finales de la época húmeda y comienzos de la época seca y la tercera correspondió a finales de la época seca y comienzo de la época húmeda. Se determinaron cuatro sitios de muestreo a diferentes altitudes: Sitio 1 a 1.800 m, Sitio 2 a 2.200 m, Sitio 3 a 2.700 m y Sitio 4 a 3.000 m. En cada sitio se dispusieron cinco trampas VanSomerenRydon a una distancia entre los 50 y 75 m una de otra entre las 8 y las 17 horas, en las que se emplearon como cebos una mezcla de frutos en estado de fermentación (banano y papaya con extracto de panela), carroña (pescado en descomposición), y humanaza (De Vries, 1987; De Vries, 1997). Las trampas se revisaron hacia el medio día y hacia el final de la tarde. Adicionalmente, se realizaron capturas con redes entomológicas y censo de individuos observados en transectos de 500 a 600 m entre las 9 y las 17 horas en lugares donde se manifiesta actividad de estos insectos por búsqueda de alimento, de agua, pareja, por termorregulación, búsqueda de plantas hospederas, percheo y actividades conductuales como territorialidad (Andrade, 1998). Los individuos capturados tanto en las redes entomológicas como en las trampas de cebo se sacrificaron mediante presión digital en el tórax, y fueron depositados en sobres de papel Milano con códigos únicos de registro y almacenados en recipientes herméticos con naftalina. En el laboratorio, los especímenes fueron puestos en cámara húmeda entre 24 y 48 horas para su posterior montaje y determinación hasta especie utilizando las claves de De Vries (1987), De Vries (1997), Andrade (1990), Andrade (1995), Borror et al. (1995) y GarcíaRobledo et al. (2002), y por comparación directa con los ejemplares de las colecciones de referencia del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICNMHN); para la nomenclatura, se utilizaron las bases de datos del Archivo Taxonómico de las Mariposas de Colombia (Andrade, 2005), Savela (1997) y del Instituto de Investigación y Recursos Biológicos Alexander Von Humboldt (2007). Luego de ser etiquetado, cada ejemplar fue llevado a colección dejando copia en la colección de referencia de la Fundación IbanascaTolima y en la Colección de Entomología del Departamento de Biología de la Universidad Nacional de Colombia.
ANÁLISIS ESTADÍSTICO
Con la información de las especies recolectadas se elaboraron matrices de presenciaausencia de especies por sitio de muestreo. Para calcular el número total de especies esperado en cada nivel altitudinal se utilizó el método de Binomiales negativas, del cual se puede inferir que tan alejados se encuentran los resultados obtenidos de los esperados (Magurran, 1989). El grado de asociación entre el nivel altitudinal y el número de especies total se estableció mediante el coeficiente de correlación no paramétrico de Sperman (Rho de Sperman; rs). De las tablas de abundancia relativa por especie y mediante el uso del programa Biodiversity Pro versión 2, se obtuvieron los índices de ShannonWeaver como indicativo de la heterogeneidad de la muestra y los índices de Simpson y de Margalef que proporcionan un valor comparativo entre eventos muestrales de la dominancia por especie.
RESULTADOS Y DISCUSIÓN
Se encontraron 94 especies de mariposas en la zona de estudio que pertenecen a 59 géneros y cinco familias. 73 de las especies encontradas corresponden a la familia Nymphalidae dentro de las cuales, 34% pertenecen a la subfamilia Satyrinae, 13,7% a la subfamilia Nymphalinae y 13,7% a la subfamilia Biblidinae; el restante 38,6% está representado por especies de las subfamilias Ithominae, Heliconinae, Acrainae, Danainae, Brassolinae, Morphinae y Melitaeinae con porcentajes entre el 1 y 10%. Otras familias encontradas en la zona fueron Pieridae con 14 especies incluidas en tres subfamilias: Coliadinae con cinco especies, Dismorphinae con tres especies y Pierinae con seis. Se encontraron seis especies de la familia Hesperiidae incluidas en dos subfamilias: Pyrginae con cinco y Hesperiinae con una. Dentro de la familia Riodinidae se encontraron dos especies y dentro de la familia Lycaenidae una sola especie ( Tabla 1). Los géneros con mayor número de especies encontradas fueron Actinote (Acrainae) con siete especies, Pedaliodes (Satyrinae) con siete, y Adelpha (Nymphalinae), Heliconius (Heliconinae), Leptophobia, Phoebis (Pieridae) y Urbanus (Hesperiidae) con tres especies. La mayor riqueza se observó a los 1.900 m y ésta disminuye gradualmente a medida que se asciende altitudinalmente hasta los 3.000 m. En total se colectaron 70 especies de mariposas a los 1.900 m, 52 a los 2.200 m, 32 a los 2.700 m y 18 a los 3.000 m. No obstante la mayor riqueza de especies se encontró a 1.900 m, los resultados obtenidos para esta estación por evento muestral presentan mayor dispersión que para las demás estaciones (Fig. 2), lo cual incide en que para esta estación, el número total de especies colectadas se aleje bastante del número total de especies esperadas mediante el empleo de binomiales negativas. Para las demás estaciones, los resultados presentan mayor correspondencia entre el número de especies recolectadas y el número total de especies esperadas, lo cual se refleja en la menor dispersión de las medidas de riqueza a través de los eventos de muestreo.
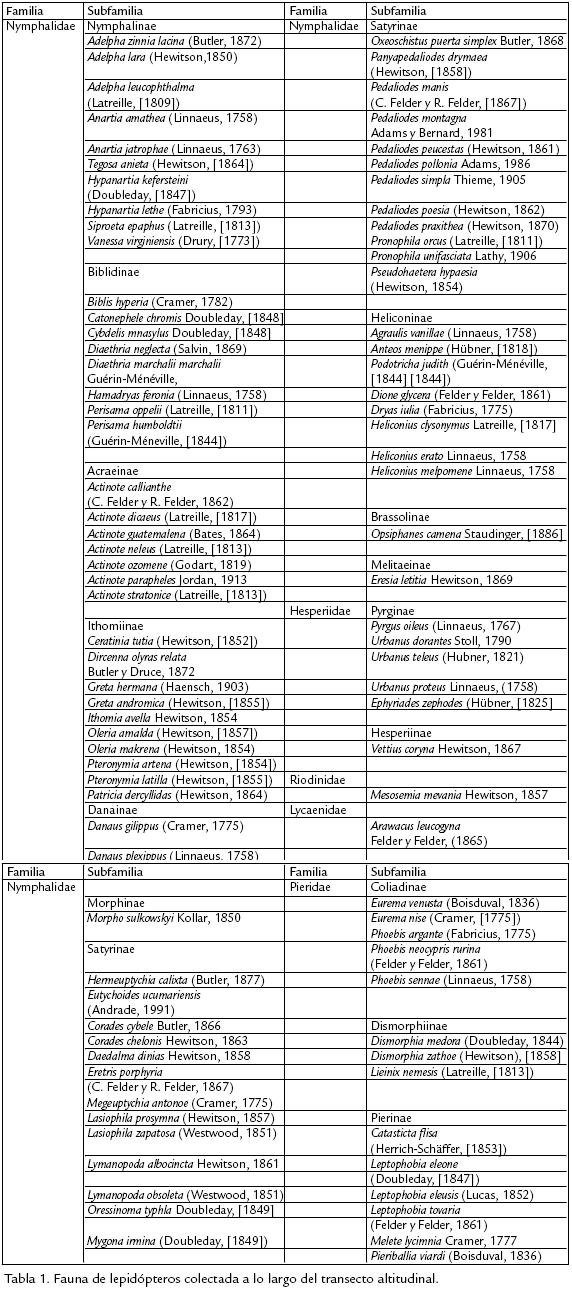
Igualmente, la riqueza disminuyó temporalmente según los resultados obtenidos por medio del Rho de Spearman (Fig. 2) con p=0,01: rs1=1,000 para el muestreo 1, rs2=0,900 para el muestreo 2,0 y rs3=0,750 para el muestreo 3, lo cual denota fuerte disminución de la riqueza hacia la época más lluviosa.
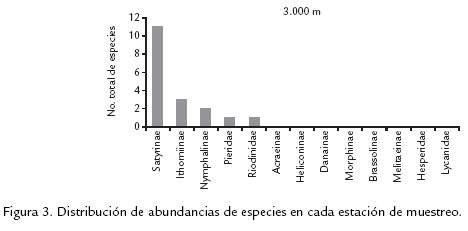
Las especies de los grupos Nymphalinae e Ithominae presentaron la mayor dominancia para los 1.900 m en todos los periodos de muestreo (Fig. 3); dicha dominancia de especies decrece a medida que se aumenta en altitud, mientras que las especies de la subfamilia Satyrinae incrementan su dominancia a medida que se asciende altitudinalmente. La diversidad, medida como heterogeneidad de la muestra según el índice de ShannonWeaver presenta una marcada disminución a medida que se asciende en el gradiente; por lo cual, existe disminución de la diversidad a mayor altitud, y este menor número de especies presenta mayor equidad poblacional la cual es más notoria en épocas de mayor humedad (Tabla 2). Los resultados de cada muestreo y su análisis en conjunto, mostraron una correlación inversa entre la riqueza, la diversidad y la altitud, lo que coincide con lo registrado en la literatura para los lepidópteros y otros grupos (Terborgh, 1971; Janzen, 1993). El mayor número de especies de mariposas registrado a los 1.900 m, puede estar relacionado con la mayor diversidad florística que se encuentra a esta altitud en comparación a las demás estaciones de muestreo (Cuatrecasas, 1989; Rangel et al., 1997); al ser la mayoría de las mariposas fitófagos especialistas, es de esperar que a mayor diversidad de recursos para las larvas, mayor la riqueza en las comunidades de mariposas, lo cual se confirma con la disminución gradual de esta de las especies de las subfamilias Heliconinae e Ithominae, las cuales son huéspedes de especies de plantas muy diversificadas solo a bajas altitudes, en contraste con las especies de la subfamilia Satyrinae, las cuales son dominantes a mayor altitud en donde la cobertura vegetal es dominada por Chusquea sp, Selaginellacea sp y briófitos de los cuales son los hospederos más frecuentes.
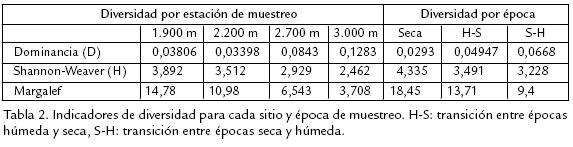
Estos resultados coinciden con los obtenidos por SánchezRodríguez (1995), quien encontró mayor proporción de mariposas generalistas a grandes niveles altitudinales lo cual relacionó con el tipo de estrategia defensiva de las plantas de media y alta montaña, las cuales presentan tasas de producción de biomasa comparativamente menores que las plantas de zonas más bajas pero con gran acumulación de reductores de digestibilidad como fenoles, taninos, cristales entre otros, factor que fomenta la diversificación de grupos generalistas más aptos al consumo de tales plantas; en nuestro caso, la gran abundancia de especies de géneros como Pedaliodes, Limanopoda,
Leptophobia y Catasticta encontradas entre los 2.200 y 3.000 m y que son hospederos frecuentes de plantas con estas características como Poaceas, Ciperaceas, Selaginellaceas y Loranthaceas (De Vries, 1987), pueden ser reflejo de esta relación plantahospedero. El análisis de clúster de correlación de diversidades muestra mayor relación entre las estaciones por encima de los 1.900 m cercana al 99%, no obstante la estación localizada a 1.900 m presenta una similitud menor a las demás y cercana al 95% (Fig. 4).
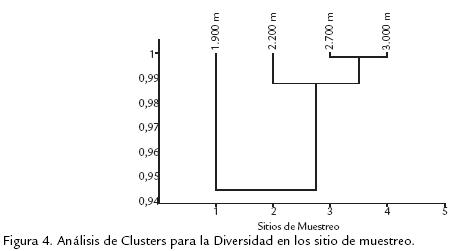
La separación de los valores de diversidad obtenidos a 1.900 m respecto al resto del gradiente permite considerar a este nivel altitudinal como un probable punto de rompimiento abrupto de las condiciones, las cuales producen reducción gradual de la riqueza y de la composición de la comunidad, y que además puede ser considerado como un nivel de transición entre las comunidades de especies de zonas bajas y altas, lo cual denomina Fagua (1999) como zona de alternancia de biotas. A partir de las matrices de abundancias relativas de especies encontradas, se ajustaron los datos a un modelo de distribución utilizando el programa estadístico PAST. En la figura 5, se muestra como los cuatro puntos de muestreo presentan distribución geométrica, en la cual pocas especies son dominantes y las demás son prácticamente raras. La dominancia aumenta al incrementarse la altitud. La mayoría de las especies fue registrada en varios sitios de muestreo y las especies Lasiophila zapatosa, Pedaliodes Manis y Leptophobia tovaria fueron observadas en todas ellas; aunque la mayor exclusividad de especies se presentó a los 1.900 m, a medida que se asciende entre los 2.200 y 3.000 m aumenta progresivamente el número de especies exclusivas para esta franja altitudinal (rsexc=0,900; p=0,05; Tabla 3). Las comunidades de altitudes bajas deben caracterizarse por tener riquezas e índices de diversidad comparativamente más altos y una baja dominancia, mientras que las comunidades de zonas más altas presentan mayor dominancia, menor riqueza, menor diversidad y una frecuencia de individuos más alta. Los resultados del actual trabajo muestran dos niveles altos en el porcentaje de exclusividad a los 1.900 m y a los 3.000 m con tendencia marcada de aumento a partir de los 2.200 m hasta los 3.000 m que concuerda con lo enunciado anteriormente, y que solo se observa al trabajar en gradientes amplios con rangos de muestreo cortos (Fagua, 1999).
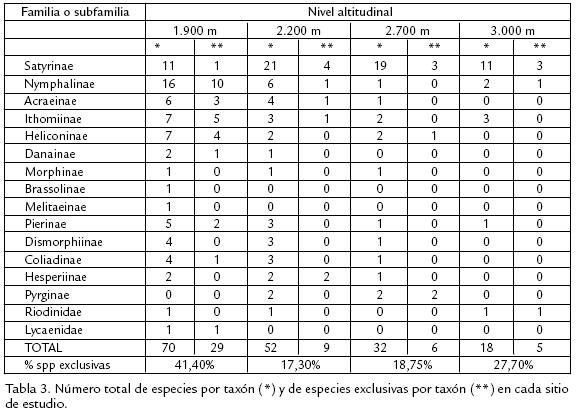
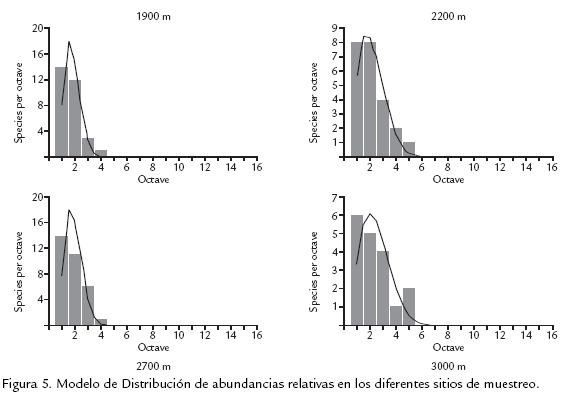
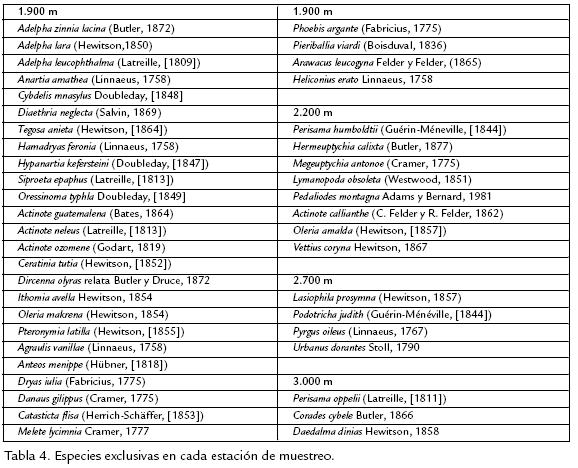
La distribución de abundancias de especies para los diferentes sitios de captura, muestra un modelo de distribución de serie geométrica en la que pocas especies son dominantes, haciendo ver a las especies restantes como prácticamente raras, e indicando una repartición diferencial de los recursos a unas pocas especies, muy seguramente debido a una preferencia de nicho por parte de las especies dominantes, lo cual disminuye la heterogeneidad de las comunidades y las medidas de Diversidad (Fuentes, 2004). La exclusividad de especies, además de permitirnos realizar afirmaciones acerca de la estructura de las comunidades con relación al recurso alimenticio disponible en los diferentes puntos del gradiente, permite proponer especies que puedan ser potencialmente utilizables como bioindicadoras de cambios en los ecosistemas (Brown, 1991). La tabla 4, muestra las especies exclusivas para cada sitio de muestreo, las cuales podrían tener un buen carácter indicador por su característica de distribución restringida, y de igual forma que la riqueza disminuye a medida que se asciende en el transecto, también el número de especies exclusivas es menor a mayor altitud, lo cual podría enmarcarse dentro del modelo de discontinuidad ecológica propuesto por Terborhg (1971) con respecto a la restricción en la distribución de especies.
AGRADECIMIENTOS
Los autores agradecen a la División de Investigaciones Bogotá DIB de la Universidad Nacional de Colombia por la financiación de esta investigación dentro del marco del proyecto “Fauna de insectos en ecosistemas naturales del departamento de TolimaColombia”, al igual que al Instituto de Ciencias Naturales de la Universidad Nacional de Colombia y a la Reserva Natural Ibanasca en el departamento del Tolima.
BIBLIOGRAFÍA
ANDRADE G. Clave para las familias y subfamilias de Lepidoptera: Rhopalocera de Colombia. Caldasia. 1990;16(77):197-200.
[ Links ]ANDRADE G. Actinote (Nymphalidae I: Acraeinae). En: Ruiz P, Pinto P, editores. Monografías de Fauna de Colombia. NymphalidaeAcraeinaeActinote. Instituto de Ciencias NaturalesUniversidad Nacional de Colombia; 1995.
[ Links ]ANDRADE G. Utilización de las mariposas como bioindicadoras del tipo de hábitat y su biodiversidad en Colombia. Rev Acad Colomb Cienc. 1998;22(84):407-421.
[ Links ]ANDRADE G. Biodiversidad de las mariposas (Lepidoptera: Rhopalocera) de Colombia. En C. Costa, S.A. Vanin y A. Melic (Eds), Proyecto de Red Iberoamericana de Biogeografía y Entomología Sistemática 2; 2002.
[ Links ]ANDRADE G. Archivo de autoridad taxonómica de las mariposas de Colombia: las especies y subespecies de las familias Papilionidae y Pieridae Instituto Alexander von Humboldt 1.0 2005. Sistema de Información sobre Biodiversidad. Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt". Bogotá, Colombia. Base de datos en línea. [Citado May 2007]. Disponible en URL: http://www.siac.net.co/sib/aat
[ Links ]BORROR D, TRIPLEHORN C, JOHNSON N. An Introduction to the Study of Insects. Philadelphia Saunders Collage; 1995.
[ Links ]BROWN K. Conservation of Neotropical Environments: Insects as Indicators. En: Collins N, Thomas J, editors. The Conservation of Insects and Their Habitats; 1991. p. 350-423.
[ Links ]CAMERO E. Estudio comparativo de la fauna de coleópteros (Insecta: Coleoptera) en dos ambientes de bosque húmedo tropical colombiano. Rev Colomb Entomol. 1999;25(34):131-136.
[ Links ]CAMERO E. Caracterización de la fauna de carábidos (Coleoptera: Carabidae) en un perfil altitudinal de la Sierra Nevada de Santa MartaColombia. Rev Acad Colomb Cienc. 2003;27(105):491-516.
[ Links ]CUATRECASAS J. Aspectos de la vegetación natural en Colombia. PerezArbelaezia. 1989;2(8):155-283.
[ Links ]De VRIES PJ. The Butterflies of Costa Rica, and their Natural History. Papilionidae, Pieridae, Nymphalidae. Princeton University Press; 1987.
[ Links ]De VRIES PJ. The Butterflies of Costa Rica, and their Natural History. Riodinidae. Princeton University Press; 1997.
[ Links ]De VRIES PJ, MURRAY D, LANDE R. Species Diversity in Vertical, Horizontal, and Temporal Dimensions of a FruitFeeding Butterfly Community in a Ecuadorian Rainforest. Biol J Linn Soc Lond. 1997;62:343-364.
[ Links ]De VRIES PJ, WALLA T. Species Diversity and Community Structure in Neotropical FruitFeeding Butterflies. Biol J Linn Soc Lond. 2001;74:1-15.
[ Links ]DEWENTER SI, TEJA T. Butterfly Community Structure in Fragmented Habitats. Ecol Lett. 2000;3:449-456.
[ Links ]DIDHAM RK, GHAZOUL J, STORCK NE, DAVIS AJ. Insect in Fragmented Forest: A Functional Approach. Elsevier Science. 1996;11(6):255-260.
[ Links ]DIDHAM RK, HAMMOND PM, STORK E, LAWTON Y. Beetle Species Responses to Tropical Forest Fragmentation. Ecol Monogr. 1998;68(3):295-323.
[ Links ]ERWIN TL. Tropical Forest: Their Richness in Coleoptera and other Arthropod Species. The Colepterist Bulletin. 1982;36(1):74-75.
[ Links ]ESCURRA E. ¿Por qué hay tantas especies raras?, la riqueza y la rareza biológicas en las comunidades naturales. Ciencias Especial. 1990;4:82-88.
[ Links ]ESQUIVEL HE, NIETO A. Diversidad florística de la cuenca alta del río Combeima. Programa PEI del Cañón del Combeima; 2003.
[ Links ]FAGUA G. Variación de las mariposas y hormigas de un gradiente altitudinal en la Cordillera oriental (Colombia). Rev Acad Colomb Cienc. 1999;2:75-90.
[ Links ]FATTORINI S. A New Method to Identify Important Conservation Areas Applied to the Butterflies of the Aegean Islands (Greece). Anim Conserv. 2006;9:75-83.
[ Links ]FUENTES PV. Composición y distribución espaciotemporal de escarabajos coprófagos (Coleoptera:Scarabeidae:Scarabaeinae) en el bosque municipal de MariquitaTolima [trabajo de grado]. Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2004.
[ Links ]GARCÍAROBLEDO CL, CONSTANTINO M, HEREDIA M, KATAN G. Mariposas comunes de la cordillera Central de Colombia. Wildlife Conservation Society Programa Colombia. Cali, Colombia; 2002.
[ Links ]GASTON KJ. The Magnitude of Global Insect Species Richness. Conserv Biol. 1991;5(3):283-296.
[ Links ]GORDON R. La taxonomía de insectos: su importancia y perspectivas. Memorias XII Congreso Sociedad Colombiana de EntomologíaSOCOLEN. Medellín, Colombia; 1985.
[ Links ]HALFTER G. A Strategy for Measuring Landscape Biodiversity. Biology International. 1998;36:3-17.
[ Links ]HERNÁNDEZ FS. Diagnóstico climático de la cuenca del río Combeima. En: Plan de Manejo de la Cuenca del Río Combeima. Universidad del Tolima CEDAR; 1991.
[ Links ]HODKINSON ID. Global Insect Diversity Revisited. J Trop Ecol. 1992;8:505-508.
[ Links ]HOLDRIDGE L. Ecología basada en zonas de vida. Instituto Interamericano de Cooperación para la Agricultura. San José, Costa Rica; 1996.
[ Links ]HOYLE M, HARBONE AR. Mixed Effects of Habitat Fragmentation on Species Richness and Community Structure in a Microarthropod Microecosystem. Ecol Entomol. 2005;30:684-691.
[ Links ]INSTITUTO DE INVESTIGACIÓN Y RECURSOS BIOLÓGICOS ALEXANDER VON HUMBOLDT. 2007 Colecciones Biológicas [Citado May 2007]; Disponible en URL: http://www.humboldt.org.co/chmcolombia/servicios/jsp/galeria/schmidt.jsp
[ Links ]JANZEN D. Sweep Samples of Tropical Foliage Insects: Effects of Seasons, Vegetation Types, Elevation, Time of Day and Insularity. Ecology. 1993;54(3):119-130.
[ Links ]KEER JT, SUGAR A, PACKER L. Indicator Taxa, Rapid Biodiversity Assessment, and Nestedness in an Endangered Ecosystem. Conserv Biol. 2000;14(6):1726-1734.
[ Links ]KREBS CJ. Ecología: estudio de la distribución y la abundancia. Segunda edición. Editorial Harla. México; 1985.
[ Links ]KREMEN C. Assessing the Indicator Properties of Species Assemblages for Natural Areas Monitoring. Ecol Appl. 1992;2(2):203-217.
[ Links ]LEONEL HF, PAEZ Y. Aproximación ecosistémica de la cuenca del río Combeima, departamento del Tolima. Revista de Ciencias Agrarias de la Universidad de Nariño. 2000;17(2):126-135.
[ Links ]MAGURRAN E. Diversidad ecológica y su medición. Barcelona, España: Ediciones Vedra; 1989.
[ Links ]MARTÍNPIERA F. Apuntes sobre al diversidad y conservación de insectos: dilemas, ficciones y ¿soluciones? Bol SEA. 1998;20:25-55.
[ Links ]OSPINA C, PEÑA E, SÁNCHEZ Y. Levantamiento hidrográfico de un tramo del río Combeima. En: Plan de Manejo de la Cuenca del Río Combeima. Universidad del Tolima CEDAR; 1979. p. 913.
[ Links ]PEARSON DL. CASSOLA F. WorldWide Species Richness Patterns of Tiger Beetles (Coleoptera: Cicindelidae): Indicator Taxon for Biodiversity and Conservation Studies. Conserv Biol. 1992;6(3):376-390.
[ Links ]RANGEL O, GARZÓN A. Parque Nacional Natural Los Nevados con referencia especial al transecto ECOANDES 1980. En: Rangel O. Colombia Diversidad Biótica 1. Bogotá, Colombia: Editorial Guadalupe; 1987. p. 184-204.
[ Links ]RANGEL O, LOWLY C, AGUILAR P. La distribución de los tipos de vegetación en las regiones naturales de Colombia. En: Rangel O. editor. Colombia Diversidad Biótica 2. Bogotá, Colombia: Editorial Guadalupe; 1997. p. 383-436.
[ Links ]SÁNCHEZ-RODRIGUEZ JF. The Efffects of Elevation on the Butterfly Communities of a Mediterranean Mountain, Sierra de Javalambre, Central Spain Lepid Soc. 1995;49(3):192-207
[ Links ]SAVELA M. 1997. Lepidoptera and Some Other Life Forms [Citado May 2007]; Disponible en URL: http://www.nic.funet.fi/pub/sci/bio/life/intro.html
[ Links ]SPARROW S, EHRLICH P. Techniques and Guidelines for Monitoring Neotropical Butterflies. Conserv Biol. 1993;8(3):800-809.
[ Links ]STORK NE. How Many Species are There? Biodivers Conserv. 1993;2:215-232.
[ Links ]SUTTON SL, COLLINS M. Insects and Tropical Forest Conservation. En: Collins N, Thomas J, editors. The Conservation of Insects and Their Habitat; 1991. p. 405-422.
[ Links ]TERBOURGH J. Distribution on Environmental Gradients: Theory and Preliminary Interpretation of Distributional Patterns in the Avifauna of the Cordillera Vilcabamba, Perú. Ecology. 1971;52(1):23-40.
[ Links ]
