










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.12 suppl.1 Bogotá Dec. 2007
HEMATOLOGÍA Y QUÍMICA SANGUÍNEA DE JUVENILES DE RUBIO (Salminus affinis PISCES: CHARACIDAE) DEL RÍO SINÚ
Hematology and Blood Chemistry of Juveniles Rubio (Salminus affinis Pisces: Characidae) Captured in the River Sinú
VÍCTOR ATENCIOGARCÍA1, M.Sc.; FERNANDO GENES LÓPEZ1, Profesional en Acuicultura; DIANA MADARIAGA MENDOZA1 Profesional en Acuicultura; SANDRA PARDO CARRASCO,2* Ph. D
1Centro de Investigación Piscícola, Universidad de Córdoba, CINPIC, Montería, Córdoba, Colombia.
2FCA/DPA, Universidad Nacional de Colombia, Sede Medellín.
*Autor para correspondencia: sandrapardocarrasco@hotmail.co Carrera 59A No. 3620, bloque 50, Medellín, Antioquia, Colombia.
Presentado 28 de mayo de 2007, aceptado 22 de julio de 2007, corregido 29 de agosto de 2007.
RESUMEN
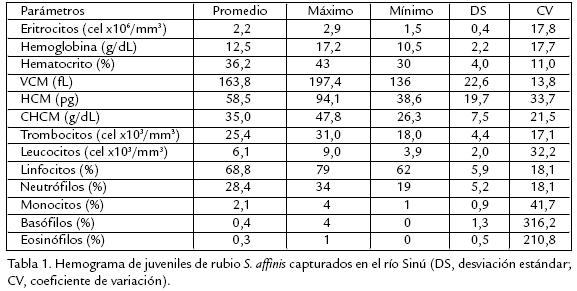
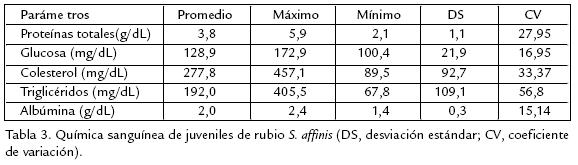
Rubio Salminus affinis es un pez reofílico, distribuido en ríos de Colombia y Ecuador. Se estudiaron características hematológicas y química sanguínea buscando generar información básica para desarrollar tecnologías de producción piscícola. Fueron examinados diez rubios juveniles aparentemente sanos (117,5±38,6 g de peso y 17,6±3,3 cm de longitud total) capturados en el río Sinú. Los peces se tranquilizaron durante cinco minutos en agua a 18 °C y se tomaron 2 mL de sangre por vacutecnia con EDTA (Vacuette®, Greiner Bioone, USA). La química se determinó utilizando reactivos para cada parámetro (ByoSystems SA, España) y el hemograma se desarrolló con técnicas tradicionales, a excepción del recuento total de leucocitos y trombocitos en el que se utilizó el reactivo de Natt y Herricks. Los frotis sanguíneos fueron teñidos con Wright y en ellos se determinaron las dimensiones celulares. El recuento total de eritrocitos fue 2,2±0,4x106/mm3, los trombocitos 25,4±4,4x103/mm3 y los leucocitos fueron estimados en 6,1±2,0x103/mm3. El conteo diferencial de leucocitos fue 68,8±5,9% linfocitos, 28,5±5,2% neutrófilos, 2,1±0,9% monocitos, 0,4±1,3% basófilos y 0,2±0,4% eosinófilos. La hemoglobina en 12,53±2,2 g/dL y el hematocrito 36,2%. Los índices eritrocitarios: volumen corpuscular medio (VCM) 163,8±22,6 fL, concentración de hemoglobina corpuscular media (CHCM) 35,0±7,5 g/dL y hemoglobina corpuscular media (HCM) de 58,5±19,7 pg. Las proteínas totales fueron 3,8±5,9 g/dL, glucosa 128,9±21,9 mg/dL, colesterol 277,8±92,7 mg/dL, triglicéridos 192,0±109,1 mg/dL y albúmina 2,0±0,3 g/dL. Los resultados sugieren que las características hematológicas de rubio están en el rango reportado para peces neotropicales aparentemente sanos.
Palabras clave: hematología, química sanguínea, Salminus affinis, peces neotropicales, leucocitos.
ABSTRACT
Rubio Salminus affinis is distributed in Colombia and Ecuador rivers. To develop new technologies of fishery production, hematological and blood chemistry of 10 juvenile Rubios captured in Sinú river, apparently healthy were examined (117.5+38.6 g of weight and 17.6+3.3 cm of total length). Fishes were calmed during five minutes in water at 18 °C and 2 mL of blood samples were taken in EDTA (Vacuette®). The chemistry was determined using ByoSystems INC (Spain), reagents and blood count were developed with traditional techniques with the exception of total leukocyte and thrombocyte for which the reagent of Natt and Herricks was used. The blood smears were dyed with Wright and cellular dimensions were determined. The total erythrocytes were of 2.2+0.4x106/mm3, thrombocytes 25.4+4.4x103/mm3 and leukocytes were estimated in 6.1+2.0x103/mm3. The differential count of leukocytes were of 68.8+5.9% Lymphocytes, 28.5+5.2% neutrophils, 2.1+0.9% monocytes, 0.4+1.3% basophils and 0.2+0.4% eosinophils. The hemoglobin in 12.53+2.2 g/dL, and the hematocrit 36.2%. The erythrocyte index: Mean corpuscular volume (MCV) 163.8+22.6 fL; mean corpuscular hemoglobin concentration (MCHC) 35.0+7.5 g/dL and mean corpuscular hemoglobin (MCH) 58.5+19.7 pg. Total proteins were 3.8+5.9 g/dL, glucose 128.9+21.9 mg/dL, cholesterol 277.8+92.7 mg/dL, triglycerides 192.0+109.1 mg/dL and albumin 2.0+0.3 g/dL. The results suggest that the hematological characteristics of Rubio are in range for neotropical apparently healthy fishes.
Key words: Salminus affinis, haematology, blood chemistry, freshwater fish; hematocrit; leucocytes.
INTRODUCCIÓN
El rubio (Salminus affinis Steindachner 1880) se distribuye en Colombia en las cuencas de los ríos Magdalena, San Jorge, Sinú, Cauca, Cesar y Ranchería y en Ecuador en el río Santiago (Lima et al., 2003). Mojica et al. (2002) lo catalogaron como una especie vulnerable a la extinción en la cuenca del río Magdalena por la fuerte presión pesquera que ha sufrido este recurso y anotaron que su situación en la cuenca del río Sinú merece especial atención en virtud de los cambios introducidos por la construcción y operación de la Hidroeléctrica Urrá (HU). Entre los principales impactos ocasionados por la HU, sobre las especies reofílicas, se destacan: interrupción de la migración reproductiva, pérdida de las áreas de maduración y desove aguas arriba, alteración de la calidad del agua en las áreas de maduración y desove aguas abajo y pérdida del potencial reproductivo (AtencioGarcía, 2000). En atención a esta situación, es de interés estudiar los aspectos básicos de su biología para avanzar en el desarrollo de tecnologías de producción en cautiverio, intentado su recuperación y cultivo (Mejía y Navarro, 2006; PardoCarrasco et al., 2007; HernándezMuñoz, 2007; LozanoPérez, 2007); aunque desde inicios de la década del 70 Dahl (1971) la señaló como una especie con potencialidad para la piscicultura continental colombiana. Conocer la hematología de los peces es de importancia para el conocimiento del estado sanitario de las poblaciones naturales de peces (Adhikari et al., 2004) pero también resulta de interés para el manejo nutricional en cautiverio; ya que puede ser evaluada la interacción entre los nutrientes buscando los mejores beneficios para la especie; así como en toxicología la presencia de tóxicos y sus efectos hematológicos (Lim et al., 2000). Affonso et al. (2002) señalan su uso en estudios ambientales para la determinación de sustancias contaminantes como agentes estresantes para peces y Martins et al. (2000) reportan que en sangre puede ser medido el estrés generado durante los procesos de confinamiento, captura y manipulación, contribuyendo a establecer protocolos menos perturbadores para los peces. Todo lo anterior se constituye en valiosa información para el desarrollo de tecnologías de producción en los campos de la nutrición, la reproducción y la sanidad. La escasez de información sobre las constantes hematológicas básicas en peces tropicales de América del Sur limita su uso en todas las áreas relacionadas anteriormente. Por tanto este estudio tiene como propósito ofrecer información sobre la hematología y algunos valores de química sanguínea de juveniles de rubio como una herramienta valiosa para el establecimiento de los controles sanitarios que favorezcan el desarrollo piscícola de esta especie, así como información básica de la especie que contribuya con futuros estudios ambientales.
METODOLOGÍA
Los peces analizados fueron capturados con atarraya en la parte baja del río Sinú entre marzo y diciembre de 2005 y trasladados en una hora en un tanque con aireación, al laboratorio de Sanidad Acuícola del Centro de Investigación Piscícola de la Universidad de Córdoba (CINPIC, Montería, Colombia) donde permanecieron en tanques de concreto de 2.000 L entre 48 y 72 horas, con agua del río Sinú, con el propósito de reducir los efectos del estrés generado por la manipulación y cambio de ambiente. Las condiciones del agua en los tanques de adaptación al cautiverio, como oxígeno disuelto (7±1 mg/L), temperatura (28±2 °C), pH (7± 0,5), dureza (50±10 mg/L), alcalinidad (80±10 mg/L) y amonio total (menor de 0,1 mg/L) se mantuvieron dentro del rango confort para las especies de la cuenca del río Sinú (AtencioGarcía, 2001). Para la extracción de la sangre los animales fueron tranquilizados en agua a 18 °C durante cinco minutos. Se utilizó vacutecnia, puncionando el paquete vascular caudal a una altura media entre la línea lateral y la base media de la aleta anal, se extrajeron 2 mL de sangre y fue recibida en tubos para vacutecnia con anticoagulante EDTA (Vacuette®, Greiner Bioone, USA). Posteriormente la sangre fue dividida en dos porciones, una de ellas se dejó completa para hacer el hemograma y la otra porción se centrifugó a 12.000 rpm durante cinco minutos para la extracción del plasma en el cual se determinó la química sanguínea.
El recuento total de eritrocitos, leucocitos y trombocitos se realizó en cámara de Neubauer de 0,0025 mm2 (Optic Labor, Alemania), para el conteo de eritrocitos se utilizó como diluyente solución salina fisiológica y para trombocitos y leucocitos se utilizó el reactivo de NattHerrick (Conroy, 1998), en ambos casos la dilución fue de 1:200. Se determinaron hematocrito (Ht) en microcentrifuga; hemoglobina (Hb) con el reactivo de Drabkin siguiendo la metodología de la cianometahemoglobina y leyendo en espectrofotómetro a 546 nm y los índices eritrocíticos volumen corpuscular medio (VCM), hemoglobina corpuscular media (HCM) y concentración de hemoglobina corpuscular media (CHCM), aplicando las fórmulas descritas por Conroy y Conroy (1987). Para realizar el recuento diferencial de leucocitos se hicieron frotis sanguíneos teñidos con coloración tipo Romanowsky (Wrigth) (Conroy y Conroy, 1987). Se tomaron fotografías de cada tipo celular con ayuda de un microscopio óptico (Carl Zeiss, Axioestar 4.3, Alemania) con cámara fotográfica digital incorporada (Canon Power Shot G5, Japón) y las mediciones celulares se realizaron con un analizador de imágenes (Carl Zeiss, AxioVisión 4.3, Alemania). Glucosa, proteína total, triglicéridos, colesterol y albúmina fueron determinados utilizando reactivos para cada parámetro del laboratorio Biosystem SA (España). Las lecturas, por separado para cada parámetro, se realizaron por medio de un analizador de química (Awareness Technology Inc. Stat Fax 3300, Alemania) a longitudes de onda de 500 nm para glucosa, colesterol y triglicéridos, 545 nm para proteína total y 630 nm para albúmina, de acuerdo con las indicaciones del fabricante para cada parámetro.
RESULTADOS
Solo fue posible examinar diez rubios juveniles debido a la dificultad para capturar ejemplares de esta especie en la cuenca del río Sinú, los cuales presentaron valores promedio de 19,63±3,3 cm de longitud total y 117,5±38,6 g de peso. El sexo no fue determinado por cuanto los ejemplares no fueron sacrificados y no se observó dimorfismo sexual en la talla analizada. Los ejemplares analizados se observaron aparentemente sanos presentando nado en línea horizontal y equilibrada, color de la piel normal, aletas caudal y anal roja y branquias de color rojo brillante. En la Tabla 1 se presentan los resultados del hemograma para rubio. Los eritrocitos son las células sanguíneas más abundantes representando el 98,8%. Estas células se caracterizaron por presentar forma ovoide y núcleo de igual apariencia; el cual se encontró en el centro de la célula formado por una masa compacta de cromatina; el citoplasma es homogéneo, acidófilo y ocupa gran parte de la célula, presentando una relación en área de 4,5 (citoplasma/núcleo). El área celular se estimó en 73,2 µm2 por lo que se puede considerar una célula de tamaño medio con respecto a las otras (fig 1A).
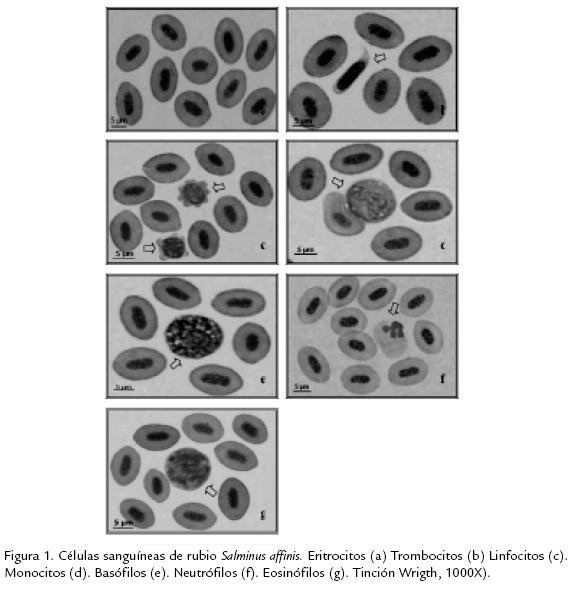
Los trombocitos fueron el 1,1% del total de las células sanguíneas. Son usualmente pequeños con formas que varían de redondas a fusiformes, pero con predominancia de estas últimas. El núcleo es frecuentemente largo siguiendo el contorno de la célula y esparcido en el citoplasma; el cual fue escaso, poco visible y sin granulaciones. La relación citoplasma/núcleo fue de 0,8 y el tamaño celular se estimó en 41,7 µm2 considerándose la célula sanguínea más pequeña (fig 1B). Los linfocitos representaron el mayor porcentaje del grupo de las células blancas (68,8 %); son células de tamaño y forma variables, observándose pequeñas, grandes, redondas y ligeramente redondas; citoplasma escaso agranular, frecuentemente irregular y basófilo. El núcleo es grande, ocupa gran parte de la célula y la relación citoplasma/núcleo fue de tres. El área celular se estimó en 50,5 µm2 (fig 1C. ). Los monocitos representaron el 2,1% de las células blancas, de mayor tamaño que los linfocitos, son células esféricas cuyo núcleo presenta forma variable entre redonda y ovalada y es frecuente observar una ligera invaginación en forma de riñón; la cromatina se encontró un poco dispersa.
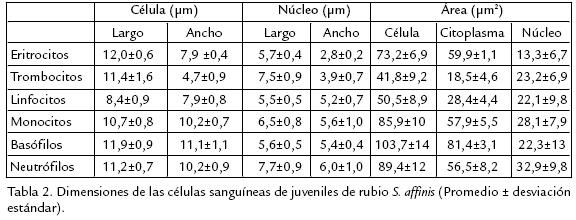
Se estimó una relación citoplasma/núcleo de dos. El citoplasma presentó aspecto basófilo y vacuolas en su interior fueron evidentes. El área celular fue de 85,9 µm2 (fig 1D). Los basófilos representaron el 0,4% de los leucocitos, de forma redonda, con citoplasma basófilo debido a la presencia de gránulos esféricos oscuros que generalmente cubren el núcleo; el cual se mostró excéntrico y presentó una relación citoplasma/núcleo de 3,7. Presentaron un área de 103,7 µm2 resultando en las células de mayor tamaño (fig 1E). Los neutrófilos son el segundo grupo de leucocitos más abundante (25,8 %), se presentaron de forma redonda y semirredonda; el núcleo se encontró excéntrico, redondo, segmentado, generalmente bilobulado, con cromatina ligeramente compactada. La relación del citoplasma/núcleo fue de 1,7. El citoplasma presentó basofilia y gránulos redondos que se tiñeron poco. El área celular se estimó en 89,4 µm2 (fig 1F). Los eosinófilos fueron las células sanguíneas de menos abundancia (0,2%), morfológicamente se observaron redondas y relativamente grandes, su núcleo es excéntrico, a menudo bilobulado. Se caracterizaron por un citoplasma con presencia de gránulos alargados numerosos y eosinofílicos (fig 1G). La Tabla 2 muestra las mediciones de largo, ancho y área de la célula del núcleo y del citoplasma de cada una de los diferentes tipos de células de S. affinis; mientras que la Tabla 3 registra los valores de los parámetros químicos en la sangre periférica del rubio en condiciones aparentemente normales y procedentes de su ambiente natural.
DISCUSIÓN
Los estudios hematológicos de las diferentes especies de peces son de interés fisiológico y ecológico, en virtud que posibilitan entender la relación entre las características sanguíneas con variables como sexo, estadio gonadal, estrés, filogenia, actividad física, hábitat y adaptabilidad al ambiente (Larson et al., 1976; Rambhaskar y Srinivasa–Rao, 1987).
Estos factores provocan que cada especie presente sus propios parámetros hematológicos; es sabido que existen diferencias sanguíneas entre especies de distintos medios osmóticos como las encontradas entre peces marinos y estuarinos, los cuales presentan una mayor cantidad de hemoglobina y eritrocitos en comparación con los de agua dulce (Romestend et al., 1982). Igualmente, la temperatura es otro parámetro que afecta los índices hematológicos; especies de un mismo medio osmótico pero en diferentes grados térmicos presentan grandes diferencias en el cuadro hemático, debido al efecto inversamente proporcional de la temperatura con la solubilidad del oxígeno disuelto en el agua.
Debido a la amplia relación de estas variables con los valores hematológicos; es importante considerar que, los resultados del presente estudio fueron comparados con especies de teleósteos que comparten preferencias térmicas y osmóticas, en particular con otros carácidos. El valor promedio de eritrocitos en juveniles de Salminus affinis fue de 2,2x106/mm3, valor similar al reportado para Salminus maxillosus (2,1x106/mm3) capturado en ambiente natural (RanzaniPaiva et al., 2001), para Prochilodus scroffa (2,1x106/mm3; RanzaniPaiva et al., 1999) y P. magdalenae (1,8 x106/mm3; ArgelBorja y QuinteroDíaz, 2007), estos dos últimos procedentes de cautiverio. También hay reportes de valores inferiores a los obtenidos en el presente trabajo, para Galaxias maculatus, especie que habita en aguas frías. JaramilloSchadebropt (2005) reportó 1,1x106/mm3, quien justificó este valor por la conducta sedentaria de la especie y consideró que los parámetros hematológicos son directamente proporcionales a la actividad del pez. De la misma manera, Martins et al. (2004a) reportaron 1,7x106/mm3 para Oreochromis niloticus. Valores superiores se reportaron para Piaractus mesopotamicus (2,9x106/mm3) por TavaresDias y Mataqueiro (2004) y para Brycon amazonicus (=B. siebenthalae; 3,8x106/mm3) por Benavides (2002). Además de tratarse de especies diferentes, estas variaciones pueden ser explicadas por diferencias en las condiciones ambientales, el origen (cautiverio versus natural) y los procedimientos a los que fueron sometidos.
En el presente estudio no fueron incluidos los trombocitos en el recuento diferencial de glóbulos blancos; a cambio, el reactivo Natt y Herrick permitió en una misma dilución diferenciar leucocitos y trombocitos permitiendo el recuento total de estas dos células. El recuento total de trombocitos encontrado en S. affinis fue de 25,4x103/mm3, similar al reportado por TavaresDias et al. (2004) en Cyprinus carpio (28,2x103/mm3). En ambos trabajos el conteo de trombocitos fue excluido del conteo diferencial de leucocitos y se utilizó la misma tinción, por lo que es posible que no se haya presentado confusión entre linfocitos pequeños y trombocitos redondos. Sin embargo, TavaresDias y Mataqueiro (2004), reportaron para P. mesopotamicus trombocitos en cantidades superiores (56,6x103/mm3) realizando el conteo de esta célula en la misma forma. Esto sugiere que es posible que estas variaciones estén asociadas a factores de tipo interespecífico. TavaresDias et al. (2002a) y TavaresDias et al. (2003) incluyendo los trombocitos en el recuento diferencial, encontraron en Piaractus mesopotamicus porcentajes de trombocitos entre 67,8 y 74,8 en cada publicación respectivamente y no reportaron valores de recuento total por lo cual se imposibilita su comparación. El número de leucocitos en teleósteos tropicales de aguas continentales se ha reportado muy variable, con valores entre 6,4 a 72,5x103/mm3 (Tandon y Joshi, 1976; RanzaniPaiva et al., 1999) presentándose incluso variaciones intraespecíficas en los teleósteos (Ueda et al., 1997; TavaresDias et al., 1999a; TavaresDias et al., 1999b; TavaresDias et al., 1999c; TavaresDias et al., 2000). En el presente estudio, el valor promedio de leucocitos para S. affinis fue de 6,1x103 /mm3, valor que se encuentra en el límite inferior del rango anotado anteriormente; sin embargo, para P. mesopotamicus, se han reportado valores mucho menores (4,6x103 /mm3; TavaresDias et al., 2002a). Los índices hematológicos calculados para S. affinis presentaron datos compatibles con los reportados en peces neotropicales.
El hematocrito para la especie en estudio fue de 36,2 %, valor superior a los reportados para C. carpio 34,3 % (TavaresDias et al., 2004) y para P. mesopotamicus 31,9 % (TavaresDias y Mataqueiro, 2004); las tres especies mencionadas son de ambientes lóticos y fueron evaluadas en condiciones aparentemente saludables. En general existe una correlación entre la hemoglobina y el hematocrito puesto que sus valores también están relacionados con la actividad y el hábitat de los peces. Se ha encontrado que los valores menores se han registrado en peces primitivos, en peces que habitan ambientes lénticos, sedentarios y bentónicos; mientras que los mayores ocurren en especies marinas, pelágicas (Larson et al., 1976) y activas (TavaresDias y Moraes, 2004). Molnár y Tamassy (1970) anotaron que los peces predadores (carnívoros), poseen mayor concentración de hemoglobina cuando se comparan con herbívoros y omnívoros. Sin embargo, la hemoglobina en S. affinis fue de 12,5 g/dL valor similar al reportado para omnívoros como B. amazonicus 13,7 g/dL (Benavides, 2002); B. hilari 12,2 g/dL, y P. mesopotamicus 12,8 g/dL (TavaresDias et al., 2003). El VCM para S. affinis fue 163,8 fL similar al encontrado para P. mesopotamicus 163,1 fL (RanzaniPaiva et al., 1999); la CHCM fue de 35,0 g/dL similar a lo reportada para B. hillari 33,9 g/dL (TavaresDias et al., 2003) y el valor de la HCM fue de 58,5±19,7 pg.
En el conteo diferencial de leucocitos fueron observados linfocitos, neutrófilos, monocitos, basófilos y eosinófilos; siendo los linfocitos y los neutrófilos las células más frecuentes en los frotis de S. affinis, situación que coincide con lo reportado en S. maxillosus (RanzaniPaiva et al., 2003); P. mesopotamicus (TavaresDias et al., 2002a; TavaresDias y Mataqueiro, 2004) y B. amazonicus (Benavides, 2002). El porcentaje de linfocitos y neutrófilos fue de 68,8±5,9 y 28,4±5,2% respectivamente. Estos resultados y los reportados en la literatura muestran que la cantidad de estos dos tipos de células es muy variable. En Leporinus macrocephalus fue establecido 29,4 % linfocitos (Martins et al., 2004b), 11,61% para Rhamdia quelen (TavaresDias et al., 2002a); para neutrófilos se encontró 92,3% para Prochilodus scrofa (RanzaniPaiva et al., 1999); y 1,1% para Piaractus mesopotamicus (TavaresDias et al., 2002b). Esta variabilidad puede atribuirse a que la presencia de las mismas está sujeta a variaciones interespecíficas de cada individuo (TavaresDias y Moraes, 2004). También es posible atribuir esta variabilidad en el conteo de leucocitos, a procesos patológicos. Se ha reportado por diversos autores que la linfopenia en los peces es consecuencia de estrés por déficit de oxígeno, septicemias bacteriales e infecciones por hongos (Conroy, 1972; Satchell, 1991; Stoskopf, 1993). La neutrofilia es habitual en diversas patologías, observándose en inflamaciones, situaciones de estrés, infecciones bacterianas y protozoarias (Stoskopf, 1993; Olabuenaga, 2000; Fernández et al., 2002). Sin embargo, los peces analizados en el presente estudio provenían de su medio natural, se mostraron aparentemente sanos, y para reducir el estrés fueron adaptados al cautiverio; para lo cual se mantuvieron en recipientes con agua con características similares a las del ambiente natural. Los monocitos fueron las células seguidas en abundancia con 2,1%, resultado algo similar al reportado para B. amazonicus (Benavides, 2002) con 1,4%; Fernández et al. (2002) señalaron que estas células se incrementan cuando existen tejidos inflamados. La abundancia de basófilos y eosinófilos puede estar relacionada con enfermedades, puesto que en condiciones normales (peces sanos), son escasas y en algunos casos ausentes. Los basófilos en los extendidos de S. affinis fueron de 0,4 %, resultados similares a los reportados por Martins et al. (2004a) para O. niloticus y Benavides (2002) para B. amazonicus, con 0,5 % para ambas especies. También se reporta la ausencia de estas células en el coespecífico S. maxillosus (Veiga et al., 2000; RanzaniPaiva et al., 2003). Los eosinófilos fueron las células menos frecuentes (0,3 %), lo cual sugiere un buen estado de salud de los animales analizados. La eosinofilia está asociada a la ocurrencia parasitaria en peces, como lo registran los reportes de Martins et al. (2004b) en Leporinus macrocefalus (13,3%) y RanzaniPaiva et al. (1999) en Colossoma macropomum y Piaractus mesopotamicus con 13,6 y 55,7% respectivamente. En S. affinis el coeficiente de variación (CV) de leucocitos, permite sugerir que el Cv de linfocitos (8,6%) mostró homogeneidad de los valores de estas células en los ejemplares analizados; mientras que los resultados de neutrófilos (18,1%) y monocitos (41,7%) se podrían considerar heterogéneos, sin que aún signifiquen enfermedad, Pero el coeficiente de variación en basófilos (316,2%) y eosinófilos (210,8 %) se consideraría extremadamente variable. Se sugiere como explicación la presencia escasa de estas células en los ejemplares analizados y cuando se observa causa una elevación considerable del coeficiente de variación. Valores elevados del CV en eosinófilos y basófilos también fueron observados por TavaresDias y Mataqueiro (2004), quienes señalaron que estos valores son habitualmente encontrados en peces.
MORFOLOGÍA CELULAR SANGUÍNEA
La morfología de los diferentes tipos celulares sanguíneas en S. affinis coincide con la reportada para otros carácidos. Las características morfológicas de los eritrocitos y trombocitos es similar a la reportada para S. maxillosus (Veiga et al., 2000; RanzaniPaiva et al., 2003) y B. amazonicus (Benavides, 2002). En los trombocitos predominaron las formas fusiformes sobre las ovaladas y redondas; aunque según Ellis (1977), estudiando las células sanguíneas en Pleuronectes platessa, encontró que las formas fusiformes en las trombocitos representaron células inmaduras y no tan abundantes como las ovaladas y redondas, en forma contraria a lo observado en este rubio. La morfología de los linfocitos y eosinófilos para S. affinis coincide con la reportada para B. amazonicus (Benavides, 2002); para S. maxillosus (Veiga et al., 2000; RanzaniPaiva et al., 2003); de igual manera las características de los basófilos coinciden con las de
B. amazonicus(Benavides, 2002). Registros morfométricos de células sanguíneas en especies neotropicales (Carácidos) son escasos en la literatura. Se destaca el reporte reciente de mediciones de largo y ancho total de la célula y del núcleo en los eritrocitos, trombocitos y leucocitos para Galaxias maculatus (JaramilloSchadebrodt, 2005). La morfometría de las células sanguíneas de rubio se podría considerar el primer registro en peces reofílicos de una cuenca colombiana. En términos generales el tamaño de las células sanguíneas de S. affinis es mayor que las reportadas para alevinos de G. maculatus (JaramilloSchadebrodt, 2005). Esta diferencia de tamaño podría estar asociada a diferencias interespecíficas, de ambiente o al estado de desarrollo biológico del pez.
QUÍMICA SANGUÍNEA
Los estudios de química sanguínea en peces tropicales suramericanos son muy escasos, a pesar de que estos permiten realizar evaluaciones diagnósticas con fines preventivos cuando se refiere al planeamiento de medidas de control de patologías. La concentración de proteínas esta altamente relacionada con el estado nutricional y la calidad del alimento. La concentración de proteína para la especie en estudio fue de 3,8 g/dL, concentración similar a la reportada para Oreochromis niloticus 3,1 g/dL (Bittencourt et al., 2003); mientras que para B. amazonicus (Benavides, 2002) se reportó una concentración de 7,2 g/dL en animales mantenidos en cautiverio y alimentados con dos raciones diarias de alimento con 24% de proteína bruta. Para S. affinis la albúmina se encontró en 2 g/dL, mientras que en juveniles en cautiverio de P. brachypomus fue de 0,9 g/dL (Sakamoto et al., 2001) y trucha arco iris presentó valores más similares de 1,65 g/dL (Miller et al., 1983). Los niveles de glucosa pueden presentar variaciones intraespecíficas, como consecuencia de talla, edad, peso, temperatura, estado nutricional, estado reproductivo y estrés. El valor de la glucosa en rubio fue de 128,9 mg/dL para esta especie; mientras que en otras especies se han reportado valores que van desde 147,6 mg/dL para B. amazonicus (Benavides, 2002); 108,23mg/dL para P. brachypomus (Eslava y Hernández, 1995); 74,8 mg/dL para P. mesopotamicus (TavaresDias et al., 2002a); hasta 60,32 mg/dL para O. niloticus (Bittencourt et al., 2003). Generalmente los niveles de lípidos decrecen en el plasma después del desove y y de períodos de ayuno; pero se incrementan cíclicamente con la alimentación en la época no reproductiva y son especialmente elevados antes del desove. Por esta razón, se presentan niveles altos de fosfolípidos asociados con gametogénesis, debido a la movilización de reservas de grasas para el desarrollo gonadal (Groff y Zinkl, 1999). El nivel de colesterol para juveniles de S. affinis fue de 277,8 mg/dL similar al reportado para juveniles B. amazonicus 299,5 mg/dL (Benavides, 2002); ambos estudios se realizaron en animales juveniles con gónadas inmaduras. Los niveles de triglicéridos para S. affinis, P. brachypomus (Eslava y Hernández, 1995) y B. amazonicus (Benavides, 2002) fueron de 192,0, 203,5 y 328,3 mg/dL respectivamente, el valor relativamente bajo de los triglicéridos en S. affinis puede atribuirse a que son animales mucho más jóvenes que las otras dos especies. Los resultados del presente estudio permiten concluir que las características hematológicas y los parámetros químicos de juveniles de S. affinis están dentro de los rangos reportados para teleósteos neotropicales evaluados en condiciones ambientales normales y aparentemente saludables.
AGRADECIMIENTOS
Los autores agradecen a la profesora Piedad Osorio de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad de Córdoba su colaboración en los análisis sanguíneos. Este estudio fue financiado en su totalidad por la Unidad de Investigación y Extensión de la Universidad de Córdoba (proyecto FMZ0204, 1120143).
BIBLIOGRAFÍA
ADHIKARI S, SARKAR B, CHATTERJEE A, MAHAPATRA CT, AYYAPPAN S. Effects of Cypermethrin and Carbofuran on Certain Hematological Parameters and Prediction of their Recovery in a Freshwater Teleost; Labeo rohita (Hamilton). Ecotoxicol Environ Saf. 2004;58:220-226. [ Links ]
AFFONSO EG, POLEZ VLP, CORREA CF, MAZON AF, ARAUJO MRR, MORAES G, RATIN FT. Blood Parameters and Metabolites in the Teleosts Fish Colossoma macropomum Exposed to Sulfide or Hypoxia. Comp Biochem Physiol. 2002;133:375-382. [ Links ]
ARGELBORJA EL , QUINTERODIAZ CP. Caracterización de los parámetros hematológicos del bocachico Prochilodus magdalenae en cultivo [trabajo de grado]. Montería (Colombia): Departamento de Ciencias Acuícolas, Facultad de Medicina Veterinaria y Zootecnia. Universidad de Córdoba; 2007. [ Links ]
ATENCIO GARCÍA VJ. Impactos de la hidroeléctrica Urrá en los peces migratorios del río Sinú. Rev. Temas Agrarios. 2000;5(9):29-40. [ Links ]
ATENCIO GARCÍA VL. Producción de alevinos de especies nativas. Rev MVZ Córdoba. 2001;6(1):9-19. [ Links ]
BENAVIDES C. Valoración hematológica y química sanguínea en el Yamú Brycon amazonicus [trabajo de grado]. Villavicencio (Colombia): Escuela de Medicina Veterinaria y Zootecnia, Universidad de los Llanos; 2002. [ Links ]
BITTENCOURT NL, MOLINARI LM, SCOARIS DO, PEDROSO RB, NAKAMURA CV, UEDANAKAMURA T, et al. Haematological and Biochemical Values for Nile Tilapia Oreochromis niloticus Cultured in SemiIntensive System. Acta Sci Biol Sci. 2003;25(2):38-59. [ Links ]
CONROY DA, ARMAS DE CONROY G. Manual de métodos de diagnóstico en ictiopatología, con especial referencia a los salmónidos. Brasilia (Bra): FAO. Documento de campo 4 (Esp); 1987. [ Links ]
CONROY DA. Manual de métodos y técnicas de laboratorio de uso común en la hematología pisciaria. Maracay (Venezuela): Pharmafish S.R.L. 1998; p.25. [ Links ]
CONROY DA. Studies on the Hematology of the Atlantic Salmon (Salmo salar). Z S L. 1972;(30):101-27. [ Links ]
DAHL G. Los peces del norte de Colombia. Bogotá (Colombia): INDERENA; 1971. [ Links ]
ELLIS AE. Leucocytes and Related cells in the Plaice Pleuronectes platessa. J Fish Biol. 1977;8:143-156. [ Links ]
ESLAVA LA, HERNÁNDEZ CP. Hematología básica de la cachama blanca Piaractus brachypomus. Rev MVZ Universidad de Los Llanos. 1995;1(1)3. [ Links ]
FERNÁNDEZ AB, RUIZ I, DE BLAS I. El sistema inmune de los teleósteos. (I): Células y órganos. Rev Aquatic 2002; (16). Disponible en: URL : http://www.revistaaquatic.com/ [ Links ]
GROFF JM, ZINKL JG. Hematology and Clinical Chemistry of Criprinid Fish: Veterinary Clinics of North America: Exotic animal practice. California 1999; 2(3):741. [ Links ]
HERNÁNDEZMUÑOZ J. Descripción morfológica del tubo digestivo de rubio (Salminus affinis) en la cuenca del río Sinú [trabajo de grado]. Montería (Colombia): Universidad de Córdoba; 2007. [ Links ]
JARAMILLOSCHADEBRODT N. Estudio hematológico del puye Galaxias maculatus en estado postlarval y adulto [trabajo de grado]. Temuco (Chile): Universidad Católica de Temuco; 2005. [ Links ]
LARSON A, JOHANSSONSJOBECK MJ, FANGE R. Comparative Study of some Haematological Blood Parameters in Fishes from the Skagerrak: J Fish Biol. 1976;9:425-40. [ Links ]
LIM C, KLESIUS PH, LI MH, ROBINSON EH. Interaction Between Dietary Levels of Iron and Vitamin C on Growth, Hematology, Immune Response and Resistance of Channel Catfish (Ictalurus punctatus) to Edwardsiella Ictalury Challenge. Aquaculture. 2000;185:313-327. [ Links ]
LIMA FC, MALABARBA LR, BUCKUP PA, DA SILVA JF, VARI RP, HAROLD A, et al. Genera Incertae Sedis in Characide. En: Reis RE, Kullander SO, Ferraris CJ, editores. Checklist of the freswater fishes of South and Central America. Porto Alegre (Brasil): Edipucrs 2003; p. 106-169. [ Links ]
LOZANOPEREZ R. Aspectos preliminares del desarrollo ovocitario en rubio (Salminus affinis) [trabajo de grado]. Montería (Col): Universidad de Córdoba; 2007. [ Links ]
MARTINS ML, MORAES FR, MORAES JRE, MALHEIROS EB. Falha na resposta do cortisol estresse por captura e por carragenina em Piaractus mesopotamicus Holmberg, 1887 (Osteichthyes: Characidae). Acta Sci Ani Sci. 2000;22:545-552. [ Links ]
MARTINS ML, TAKAHASHI D, YAMAGUCHI D, PILARSKY F, RIBEIRO K, CASTRO M, et al. Physiological and Haematological Response of Oreochromis niloticus (Osteichthyes: Cichlidae) Exposed to Single and Consecutive Stress of Capture. Acta Sci Anim Sci. 2004a;26(4):449-56. [ Links ]
MARTINS ML, TAVARESDIAS M, FUJIMOTO R, ONAKA EM, NOMURA DT. Haematological Alterations of Leporinus macrocephalus (Osteichtyes: Anostomidae) Naturally Infected by Goezia leporini (Nematoda: Anisakidae) in Fish Pond. Arq Bras Med Vet Zootec. 2004b;56(5):640-646. [ Links ]
MEJÍA KE, NAVARRO YP. Determinación de parásitos en rubio (Salminus affinis Steindachner, 1880) de las cuencas de los ríos Sinú y San jorge [trabajo de pregrado] Montería (Colombia): Universidad de Córdoba; 2006. [ Links ]
MILLER WR III, HENDRICKS AC, CAIRNS JJR. Normal Ranges for Diagnostically Important Hematological and Blood Chemistry Characteristics of Rainbow Trout (Salmo gairdneri). Can J Fish Aquat Sci. 1983;40:420-425. [ Links ]
MOJICA JI, CASTELLANOS C, USMAS J, ÁLVAREZ R. Libro rojo de peces dulce acuícolas de Colombia: La serie libros rojos de especies amenazadas de Colombia. Bogotá (Colombia): Universidad Nacional de Colombia; 2002. [ Links ]
MOLNÁR G, TAMASSY E. Study of the Haemoglobin Content of a Single Erythrocyte (M index) in Various Cultured Fish Species. J Fish Biol. 1970;2:267-171. [ Links ]
OLABUENAGA E. Sistema inmune de peces. Gayana. 2000;64(2):205-15. [ Links ]
PARDO S, MEJÍA K, NAVARRO Y, ATENCIO V. Prevalencia abundancia de Contracaecum sp. en rubio Salminus affinis en el río Sinú y San Jorge: Descripción morfológica. Rev MVZ Córdoba 2007;12(1):887-896. [ Links ]
RAMBHASKAR B, SRINIVASARAO K. Comparative Haematology of Ten Species of Marine Fish from Visakhapatnam Coast. J Fish Biol. 1987;30:59-66. [ Links ]
RANZANIPAIVA MJ, SALLES F, EIRAS J, EIRAS A, MASSATOSHI C, ALEXANDRINO A. Analises hematologicas de Curimbatá (Prochilodus scrofa), Pacu (Piaractus mesopotamicus) e Tambaqui (Colossoma macropomum) das estações de piscicultura do instituto de pesca, estado de São Paulo. Bol. Inst. de Pesca; 1998/1999, 25(unico): 77-83. [ Links ]
RANZANI PAIVA MJ, RODRIGUEZ E, VEIGA ML, EIRAS AC. Association Between the Hematological Characteristics and the Biology of the Dourado Salminus maxillosus Valenciennes, 1840 from MogiGuaçu River, state of São Paulo, Brazil. Acta Sci Anim Sci. 2001;23(2):52733. [ Links ]
RANZANI PAIVA MJ, RODRÍGUEZ EL, VEIGA ML, EIRAS AC, CAMPOS BE. Differential Leukocite Counts in Dourado Salminus maxillosus Valenciennes, 1840, from the mogi guaçu river, Pirassununga, SP. Rev Braz J Biol. 2003; 517-25. [ Links ]
ROMESTEND B, HALSBAND E, BRAGONI G, KNEZEVIC G, MARIC B, PROCHOW F. Haematological Study of Erythrocytic Constants in Some Marine and Freshwater Fishes. Rev Trav Inst Peches Marit Nante.s 1982;46(2):147-56. [ Links ]
SAKAMOTO K, LEWBART GA, SMITH II TM. Blood Chemistry Values of Juvenile Red Pacu (Piaractus brachypomus). Vet. Clin. Path. 2001;30(2). [ Links ]
SATCHELL G. Physiology and Form of Fish Circulation. Cambridge, editor. Univ. Press. Australia; 1991. [ Links ]
STOSKOPF M. Fish Medicine. Philadelphia (Usa): Saunders; 1993. [ Links ]
TANDON RS, JOSHI BD. Total Red and White Blood Cell Count of 33 Species of Fresh Water Teleosts. Z. Tierphysiol. Tierernaehrg. Futtermittelkd. 1976;37:293-97. [ Links ]
TAVARES DIAS M, SWANDRIM EF, CAMPOS FE. Características hematológicas do tambaqui Colossoma macropomum Cuvier, Osteichthyes, Characidae) em sistema de monocultivo intensivo. II. Leucocitos. Rev Braz Zool. 1999a;16(1):841-75. [ Links ]
TAVARES DIAS M, FRASCÁSCORVO CM, CAMPOS FILHO E. MORAES F. Características hematológicas de teleósteos brasileiros. IV. Parâmetros eritroleucométricos, trombométricos e glicemia do matrinxã Brycon cephalus Günther, 1869 (Osteichthyes:Characidae). Ars Veterinaria. 1999b;15:149-53. [ Links ]
TAVARES DIAS M, TENANI RA, GIOLI LD, FAUSTINO C. Características hematológicas de teleósteos brasileiros. II. Parâmetros sangüíneos do Piaractus mesopotamicus Holmberg, 1887 (Osteichthyes: Characidae) em Policultivo Intensivo. Rev Bras Zool. 1999c;16(2):423-31. [ Links ]
TAVARES DIAS M, SCHALCH HC, MORAES F, ONAKA EM, MARTINS ML. Haematological Characteristics of Brazilian Teleosts. III. Parameters of the Hybrid Tambacu (Piaractus mesopotamicus Holmberg, 1887 x Colossoma macropomum Cuvier, 1818) (Osteichthyes: Characidae). Rev Bras Zool. 2000;17(4):899-926. [ Links ]
TAVARES DIAS M, MARTINS ML, SCHALCH SHC, ONAKA EM, Moraes JRE, Quintana CIF, MORAES FR. Alterações hematológicas e histopatológica em pacus Piaractus mesopotamicus Holmberg, 1887 (Osteichthyes: Characidae) tratatos com sulfato de cobre (CuSO4). Acta Sci Anim Sci. 2002a;24(2):547-554. [ Links ]
TAVARES DIAS M, MELO F, MORAES G. MORAES F. Haematological Characteristics of Brazilian Teleosts: IV. Parameters of Jundiá Rhamdia Quelen (Pimelodidae): Cienc Rural. 2002b;32(4):693-98. [ Links ]
TAVARES DIAS M, HENRIQUE S, SCHALCH C, MORAES F. Hematological Characteristics of Brazilian Teleosts: VII. Parameters of Seven Species Collected in Guariba, São Paulo State, Brazil. B. Inst Pesca. 2003;29(2):109-15. [ Links ]
TAVARES DIAS M, MATAQUEIRO M. Características hematológicas, bioquímicas e biométricas de Piaractus mesopotamicus Holmberg, 1887 (Osteichthyes: Characidae) oriundos de cultivo intensivo: Acta Sci Biol Sci. 2004;26(2):157-62. [ Links ]
TAVARESDIAS M, MORAES F. Hematologia de Peixes Teleósteos. Ribeirão Preto (Brasil); 2004. [ Links ]
TAVARES DIAS M, RIZZI F, FERREIRA S, CAMPOSFILHO E, MORAES F. Células sangüíneas, eletrólitos séricos, relação hepato e esplenossomática de carpacomum, Cyprinus carpio (Cyprinidae) na primeira maturação gonadal: Acta Sci Anim Sci. 2004;26(1):73-80. [ Links ]
UEDA IK, MATUSHIMA E, EGAMI MI. Estudos hematológicos do sangre periférico de Oreochromis niloticus (Linnaeus, 1758) (Cichlidae, Teleostei). Braz J Vet Res Anim Sci. 1997;34(Pt1):270-75. [ Links ]
VEIGA MI, EGAMI MI, RANZANIPAIVA MJ, RODRIGUES E. Aspectos morfológicos y citoquímicos de las células sanguíneas de Salminus maxillosus Valenciennes, 1840 (Characiformes, Characidae). Rev Chil Anat. 2000;18(2):245-50. [ Links ]

