










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.13 no.2 Bogotá May./Aug. 2008
CAMBIOS EN LA ESTRUCTURA DE LA POBLACIÓN DE Boeckella gracilis (CRUSTACEA, CENTROPAGIDAE) EN EL PLANCTON DEL LAGO DE TOTA, BOYACÁ - COLOMBIA
Changes Of The Population Structure Of Boeckella gracilis (Crustacea, Centropagidae) In The Plankton From Lago de Tota, Boyaca - Colombia
ADRIANA DEL PILAR GONZÁLEZ ANGARITA1, Bióloga; NELSON JAVIER ARANGUREN RIAÑO1, M.Sc.; SANTIAGO GAVIRIA MELO1,2, Ph. D.
1 Unidad de Ecología en Ecosistemas Acuáticos - UDESA. Escuela de Ciencias Biológicas. Universidad Pedagógica y Tecnológica de Colombia. Carretera Central del Norte, UPTC, Tunja, Boyacá, Colombia. agonzalez236@yahoo.com nelson.aranguren@uptc.edu.co
2 University of Vienna, Department of Freswater Ecology, and Technisches Büro für Biologie, Fred-Raymondgasse 19/2/4, A-1220 Vienna, Austria. santiago.gaviria@gmx.at
Presentado 3 de octubre de 2007, aceptado 12 de diciembre de 2007, correcciones 11 de abril de 2008.
RESUMEN
Se realizó un seguimiento a la población de Boeckella gracilis Daday, 1902 en el lago de Tota, Boyacá, Colombia, durante seis meses con periodicidad quincenal a partir de mayo de 2004. La densidad poblacional fue de 11.968 Ind.m-3 en promedio y expresó un incremento progresivo hasta el final del estudio que corresponde a época de fuertes lluvias en la zona y aguas altas en el lago. Los machos adultos dominaron en densidad y biomasa, constituyendo el 32,5% y el 35,8% de la población, respectivamente. La fecundidad promedio se estimó en 0,32 huevos por hembra. En la época de aguas altas se presentó un aumento de la actividad reproductiva evidenciado por el incremento de la fecundidad y el mayor número de copepoditos. Con base en el análisis de componentes principales, la transparencia, el pH y el oxígeno disuelto fueron las variables determinantes de la expresión temporal del hábitat. El comportamiento de la biomasa de la población se asoció positivamente con la variación de la densidad de los adultos y copepoditos y negativamente con la concentración de oxígeno disuelto en el lago.
Palabras clave: Boeckella gracilis, Copepoda Calanoida, lago andino, zooplancton.
ABSTRACT
The population of Boeckella gracilis Daday, 1902 of the lago de Tota, Boyacá, Colombia, was studied during a period of six months forthnightly starting in May 2004. The population density with a mean of 11.968 nd.m-3 showed a progressive increase to the end of the period of study corresponding to high values of rain intensity in the region and high water level of the lake. Mature males dominated in density and biomass, achieving 32.5% and 35.8% respectively. Fecundity indicated a mean of 0.32 eggs for female. In the period of high water level an increase of the reproductive activity occured and was evidenced by the increment on fecundity and the highest copepodite density. The analysis of main components showed that transparency, pH and dissolved oxygen were the decisive variables for the temporal expression of the habitat. Biomass variation was positively associated to adult and copepodit density variation and negatively related to oxygen dissolved concentration in the lake.
Key words: Boeckella gracilis, Copepoda Calanoida, Andean lake, zooplankton.
INTRODUCCIÓN
El lago de Tota, Boyacá, Colombia, se considera un ecosistema estratégico para la región (Corpoboyacá, 1998) por su capacidad de almacenamiento y los servicios ambientales que brinda (Márquez, 1996). En este importante sistema lacustre habita Boeckella gracilis (Copepoda: Centropagidae), y es la especie dominante tanto en abundancia como en biomasa dentro de la comunidad zooplanctónica del lago (Monroy, 2002). Por esta razón, el estudio de esta población en el lago de Tota, puede constituir un aporte importante para comprender el funcionamiento del lago y, sus atributos podrían ser utilizados como indicadores de las características generales del ecosistema, como se ha evidenciado en estudios de otros copépodos planctónicos (López, 1994; Margalef, 1983). El género Boeckella es de gran interés biogeográfico ya que tiene una distribución restringida a ambientes continentales del hemisferio sur (Menu-Marque y Locascio, 1998). Al norte del Ecuador solo se conoce en Colombia hasta los 6° N y en China y Mongolia entre 41° y 50° N (Bayly, 1992; Guo, 1999). En Sur América, el género se distribuye a lo largo de los Andes hasta Colombia donde está limitado a lagos y lagunas altoandinas. La especie Boeckella gracilis habita en la cordillera Oriental en los páramos de Sumapaz, Chisacá, Pisba, Guantivá y La Rusia y fuera de ella solo se ha reportado en la laguna del Otún (Gaviria, 1989). En Colombia forma parte del plancton pero también habita charcas de páramo (Gaviria, 1989, Gaviria y Aranguren, 2007). Si bien existen algunos estudios morfológicos, taxonómicos y de distribución de otras especies del género (Menu-Marque y Balseiro, 2000; Bayly et al., 2003) y uno sobre la dinámica poblacional de B. gracilipes (Balseiro, 1991), no hay referencias sobre la biología y ecología de B. gracilis.
MÉTODOS
ÁREA DE ESTUDIO
El lago de Tota es un sistema de origen tectónico (Rangel y Aguirre, 1983) con influencia glacial (Gaviria, 1992) localizado a 3.015 msnm entre los 5º28’13’’ -5º38’14’’ N y los 72º50’38’’ -73º00’00’’ W en la cordillera Oriental hacia el suroriente del departamento de Boyacá en los municipios de Tota, Cuítiva y Aquitania (Corpoboyacá, 1998). Tiene una profundidad media de 30 m y máxima de 62 m, un perímetro de 47 Km, un espejo de agua de 60 Km², un volumen máximo de 1.920x106 3 y se divide en dos sectores: lago Chico y lago Grande (Armada Nacional, 1998). El lago de Tota es un sistema polimíctico cálido, posee aguas poco mineralizadas y pH con una ligera tendencia a la basicidad. La relación C:N es baja, lo cual es típico en lagos tropicales altoandinos y se considera como un sistema oligotrófico (Bermúdez y Moreno, 2002; Cardozo y Pita, 2004). El lago está afectado por dos regímenes pluviales: el efecto interandino bimodal en la cuenca occidental (730 mm.año-1) y el monomodal predominante en la región nororiental (945 mm.año-1) influenciado por los llanos orientales, donde se presenta una estación lluviosa desde abril hasta noviembre y una seca de diciembre a marzo. El área de estudio dentro del lago parece estar más afectada por el régimen monomodal. El comportamiento del nivel del agua en el año 2004 se caracterizó por presentar valores bajos durante los periodos iniciales (mayo), incrementándose gradualmente hasta alcanzar niveles máximos durante agosto (3015,36 msnm) y septiembre (3015, 48 msnm) según González, 2005. Así mismo, los valores de precipitación en la región registrados por Cordero et al., 2005 fueron altos en los meses de septiembre (98 mm) y octubre (110 mm) del año 2003.
TRABAJO DE CAMPO
Se realizaron muestreos quincenales durante seis meses entre mayo y octubre de 2004. Los dos sitios de muestreo se ubicaron en la zona limnética central de cada uno de los sectores del lago: lago Chico y lago Grande. Se registraron valores de transparencia empleando un disco Secchi; en la superficie del lago se tomaron datos de temperatura, conductividad eléctrica (equipo portátil Schott Geräte CG858), de pH (potenciómetro Orion 210) y oxígeno disuelto (Winkler) del agua según las recomendaciones de Ruiz, 2002. La clorofila a se midió con periodicidad mensual en muestras integradas tomadas de la zona eufótica, transportadas de forma refrigerada para su posterior análisis en laboratorio por el método espectrofotométrico. Las muestras biológicas se obtuvieron con una trampa tipo Schindler-Patalas de 10 L de capacidad (Wetzel y Likens, 1991) en tres profundidades: 5 m, 15 m y 25 m (lago Chico) y 5 m, 15 m y 30 m (lago Grande). Las muestras de cada sitio (30 L) fueron integradas y filtradas a través de una red con diámetro de poro de 45 µm. La preservación se hizo en formalina al 4% (Boltovskoy, 1995).
TRABAJO DE LABORATORIO
Los individuos de B. gracilis fueron agrupados según el estadio de desarrollo (seis fases naupliares, cinco de copepodito y fase adulta), según las indicaciones de Hutchinson, 1967, y Einsle, 1993. Se realizaron conteos totales de los individuos por estadio, discriminando en el último (adulto) machos y hembras; en éstas últimas se consideró la presencia de saco ovígero y número de huevos. Con la ayuda de un ocular micrométrico se realizaron mediciones de la longitud corporal (sin cerdas furcales) de cada individuo. El cálculo de la biomasa se adelantó mediante el uso de la ecuación y =0,5593e 0,0024x, la cual establece la relación entre longitud corporal y peso seco para B. gracilis en el lago de Tota (Monroy, 2002).
TRATAMIENTO DE DATOS
La densidad y demás atributos poblacionales considerados para cada periodo deestudio se determinaron como el promedio de las expresiones parciales de cadavariable en los dos puntos de muestreo. Los recuentos discriminaron los individuospor estadio de desarrollo para así establecer la distribución por edades. La propor-ción entre sexos fue calculada mediante una proporción estimativa que indica ladiferencia entre machos y hembras. La fecundidad se valoró con base al número dehuevos y hembras adultas encontradas (Krebs, 1985).La tendencia del comportamiento de los parámetros ambientales considerados se es-tableció mediante estadísticos de resumen (promedio aritmético) y mediante el coefi-ciente de variación, el cual permite comparar variables que no se expresan en lasmismas unidades (Daniel, 1991). El grado de asociación entre dichas variables se esti-mó por el análisis multivariado de ordenación ACP (Análisis de Componentes Princi-pales) el cual permite evaluar matrices de registros físicos y químicos y, conjuga enpocas variables, alta cantidad de información con base en las correlaciones encon-tradas, empleando el paquete estadístico Statgraphics Plus versión 2.0. La relaciónentre los parámetros ambientales y aspectos poblacionales se estableció medianteanálisis de correlación simple, procedimiento que determina la dependencia esta-dística entre dos variables (Gutiérrez, 1995). En los resultados, las épocas de estudiose presentan con abreviaturas del mes correspondiente y números romanos de acuer-do al orden consecutivo.
RESULTADOS
VARIABLES AMBIENTALES
El promedio de las variables ambientales y su respectivo coeficiente de variación sepresentan en la Tabla.1. Se encontró que la clorofila a,el oxígeno disuelto y laconductividad fueron los parámetros más estables durante el periodo estudiado.
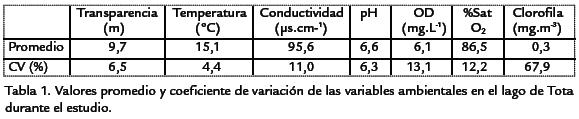
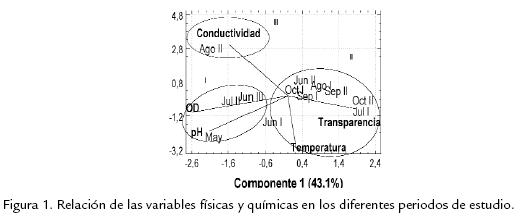
El análisis de componentes principales (Fig. 1 ) permite observar la relación de las varia-bles físicas y químicas en función de los periodos de muestreo; los componentes 1 y 2explican el 73,5% de la información. Respecto al primer componente se conforma el gru-po I determinado por valores altos de oxígeno disuelto y pH, correspondiente a las pri-meras etapas del muestreo (nivel del agua bajo en el lago), así como el grupo II determi-nado por valores altos de transparencia, comportamiento que se dio en las etapasmedia y final del muestreo (nivel del agua alto en el lago). Respecto del componente 2,se disocia el grupo III conformado por el muestreo de agosto II y determinado por losvalores altos de la conductividad eléctrica estimados en esta época.
VARIABLES POBLACIONALES
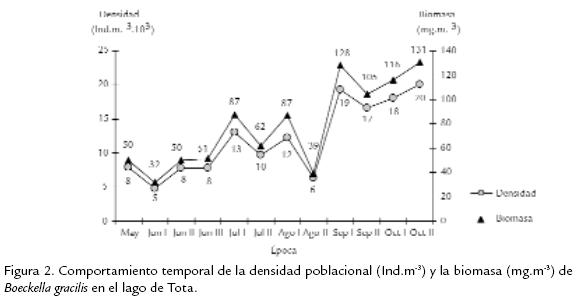
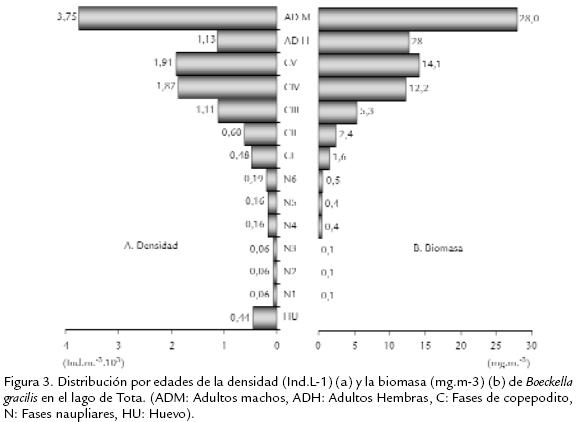
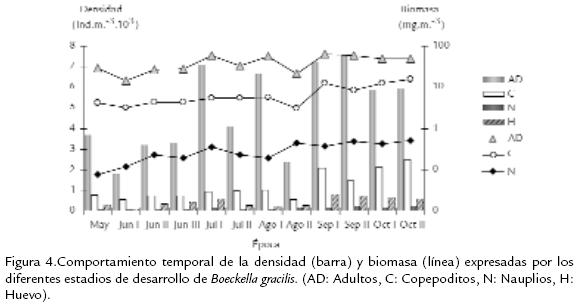
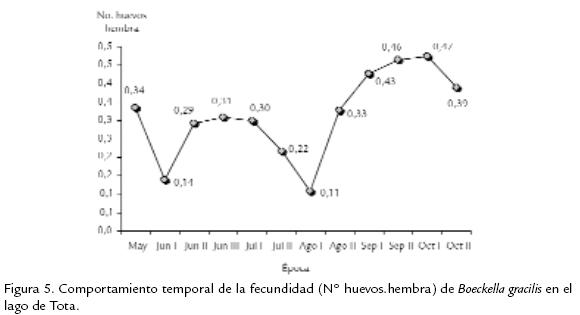
La población de B. gracilis en el lago de Tota, expresó una densidad promedio de 11.968 Ind.m-3 y su variabilidad durante el estudio se presenta en la Fig. 2 . Se destaca un incremento temporal progresivo de la densidad, con un máximo para el periodo octubre II. La distribución por edades de la densidad y la biomasa de B. gracilis se ilustra en la Fig. 3 . Los machos contribuyen con una proporción significativa de la densidad (32,5 %) y de la biomasa (35,8 %) poblacional, seguidos por las fases de copepodito V (densidad 16,6 %, biomasa 18,1 %); el aporte de las fases naupliares es menor del 6 % de la densidad total y menor del 2 % de la biomasa total. La variación temporal en densidad y biomasa considerando los estadios de desarrollo del copépodo se presentan en la Fig. 4 . Los adultos y los copepoditos dominan de forma constante tanto en densidad como en biomasa. Por otra parte, la proporción de sexos en B. gracilis presenta un promedio de 3 machos por hembra y su variación temporal se resume en la Tabla.2. La fecundidad de la especie en el lago de Tota se estimó en un promedio de 0,32 huevos por hembra. En la Fig. 5 se registra un comportamiento fluctuante de esta variable, con una reducción importante en junio I y agosto I en contraste con lo observado en septiembre II y octubre I.

DISCUSIÓN
VARIABLES AMBIENTALES
Los valores para las variables ambientales encontradas son comparables a los reportes de Bermúdez y Moreno, 2002, Cardozo y Pita, 2004 adelantados en el mismo lago. Con base en la información física y química observada y considerando los coeficientes de variación, el lago de Tota expresó bajo grado de fluctuación durante el periodo de estudio. Solo se notó un coeficiente de variación elevado en el caso de la clorofila a (67%, promedio 0,3 mg.m-3), lo que puede deberse a un efecto del pastoreo temporal heterogéneo por parte de organismos del zooplancton. Además de B. gracilis los otros crustáceos filtradores presentes en el lago de Tota son Ceriodaphnia pulchella (Sars, 1862), Daphnia pulex (Leydig, 1860), Bosmina longirostris (O.F.Müller, 1785), Diaphanosoma birgei (Korinek, 1981), Metacyclops leptopus totensis (Reid, Molina Arévalo y Fukushima, 1990) y Tropocyclops prasinus altoandinus (Gaviria, 1994) según Monroy et al., 2004, aunque no son dominantes. La relativa estabilidad física del lago de Tota en la zona limnética puede asociarse con el efecto de la gran superficie y volumen del lago. La extensión de la zona eufótica (profundidad Disco Secchi: 9,7 m) indica condiciones de ultraoligotrofía (Henao, 1987).
VARIABLES POBLACIONALES
La densidad poblacional de B. gracilis en el lago de Tota es similar a la reportada para otras especies de calanoideos en la zona altoandina de Colombia como Colombodiaptomus brandorffi Gaviria, 1989 en la laguna de Chingaza, con densidades entre 3.100 Ind.m-3 y 28.000 Ind.m-3 (Gaviria, 1993). Los incrementos de la densidad no se asociaron con eventos reproductivos durante el estudio. A pesar de que se observa tendencia al incremento de la fecundidad con el aumento en la densidad de la población (Fig. 2 y Fig. 5 ), la relación no fue significativa (r=0,55 r2=30,3% p=0,06). Los procesos de redistribución y agregación espacial de los individuos por efecto de vientos y corrientes internas pueden afectar la expresión de la densidad de las poblaciones en el zooplancton (Rinke et al., 2007). La variación temporal de la biomasa (Fig. 2 ) se asoció directamente con los cambios de la densidad de los adultos (r= 0,97 r2=95,55% p < 0,001) y los copepoditos (r=0,79 r2=62,82% p=0,002). A su vez, el oxígeno disuelto presentó relación significativa (de tipo negativo) con la expresión de la biomasa (r=-0,7 r2=49,4% p=0,01), indicando que una mayor biomasa requiere de una mayor demanda por este recurso (Blazka et al., 1982). Este tipo de relación también se observó con el pH, aunque en este caso no fue significativa (r=-0,5 r2=31,1% p=0,06). La reducción del pH sugiere que su valor está afectado por el aumento de la concentración de CO2, que es producto del incremento de la actividad metabólica generado por una mayor biomasa. Considerando el comportamiento temporal de la biomasa de B. gracilis con respecto al tamaño de los individuos, se encontró que los mayores índices están relacionados significativamente con las fases adultas (r=0,9 r2=95,5% p < 0,001) y los estadios de copepodito (r=0,7 r2=62,8% p < 0,01), mientras que con la fase naupliar no se presentó un nivel de significancia. En general, la proporcionalidad en la estructura de edades se mantiene estable durante todo el estudio (Fig. 4 ). De acuerdo con Lampert y Sommer, 1997, es posible que una población sea muy dinámica pero aparece estática debido a que el proceso de incremento y decremento está en balance; posiblemente este es el caso de la población de B. gracilis en el lago de Tota. La baja proporción de estadios naupliares coinciden con los datos obtenidos en el estudio realizado por Monroy, 2002, en este sistema. El menor número de nauplios con respecto a los demás estadios de desarrollo se debe probablemente a que en esta fase se presenta una mayor mortalidad (Balseiro, 1991; Infante, 1981; Lewis, 1979; Saunders y Lewis, 1988). Con respecto a la distribución de la biomasa por edades, la proporción de los aportes se relaciona directamente con el tamaño y la densidad de los individuos en cada fase de desarrollo (Gallo et al., 2004); tal es el caso en B. gracilis donde los machos, al ser más numerosos, aportan mayor cantidad de biomasa al sistema con respecto a los demás estadios; ellos superan inclusive los aportes de las hembras adultas aunque estas sean de mayor tamaño. Por su parte, los nauplios, al presentar menor talla y densidad explican los valores más bajos de biomasa observados en el lago a través de la época de estudio. Considerando ahora la proporción de sexos, la dominancia de machos respecto de las hembras podría fundamentarse en lo que afirma Dajoz, 1974; este autor asegura que la proporción de sexos es raramente igual a la unidad. En copépodos planctónicos del orden Calanoida es más común que los machos dominen en número sobre las hembras. Tal es el caso en Heterocope saliens (Lilljeborg, 1863) y Acanthodiaptomus denticornis (Wierzejski, 1887) en el lago di Monscera en Italia, mientras que en Arctodiaptomus bacillifer (Koelbel, 1885) del mismo lago el dominio de los sexos se presenta de forma alterna (revisado por Hutchinson, 1967) durante el desarrollo de la población. La dominancia de machos sobre hembras se explica porque el desarrollo de los machos en la etapa subadulta es más rápido en comparación al de las hembras (Margalef, 1983; Maier, 1989). Por otra parte, las hembras (especialmente las ovígeras) al ser de mayor tamaño y movimientos más lentos que los machos, están sometidas a mayor depredación visual por parte de peces (Balseiro, 1991; Gallo et al., 2004), aves e invertebrados. Se ha evidenciado depredación por peces en el lago de Tota por datos de la dieta alimenticia de la trucha arcoiris Oncorhynchus mykiss (Walbaum, 1792), la cual es el principal recurso íctico del sistema (Mora et al., 1992). Observando el comportamiento de la fecundidad de B. gracilis durante el estudio encontramos que este tiende a aumentar hacia el final del periodo (Fig. 5 ) debido a un incremento (cercano al 8%) en el número de hembras ovígeras. Por otra parte, a pesar de que la concentración del alimento es un factor de control del crecimiento y reproducción de las especies zooplanctónicas (Fernandes y Pinto-Coelho, 2000), la fecundidad de B. gracilis no se relacionó con la clorofila a durante el estudio (r=0,5 r2=21,2% p=0,1). El bajo número de huevos encontrados se explica por la baja productividad del lago en la zona limnética. En lagos de diferente productividad en Polonia se encontró que el menor número de huevos en Thermocyclops oithonoides (Sars, 1863) ocurre en los lagos más oligotróficos (revisado por Hutchinson, 1967). De acuerdo a lo observado la población de B. gracilis en el lago de Tota se reproduce constantemente, lo que se evidencia con la presencia de hembras con saco ovígero y/o espermatóforos en todas las épocas. Según Hutchinson, 1981, la reproducción continua es conveniente para que la población se mantenga, especialmente en este caso, donde las hembras pro-ducen solamente de uno a dos huevos (ocasionalmente tres o cuatro) por saco ovígero. Dicho comportamiento reproductivo constante es además típico de poblaciones con reclutamiento continuo que originan generaciones que se traslapan (Lampert y Sommer, 1997; Lomniki, 1988). A partir de este estudio no resulta evidencia de que el bajo número de huevos se relacione con la temperatura. No se notó producción de huevos de resistencia durante este estudio, pero es poco probable en el lago de Tota por sus condiciones limnéticas. Se han evidenciado presencia de huevos de resistencia de Boeckella hamata (Brehm, 1928) en el sedimento del lago Waihola en Nueva Zelanda, donde se producen como estrategia para recolonizar el lago después de períodos de alta salinidad (Hall y Burns, 2001). El desarrollo de este tipo de huevos podría ocurrir en charcos de páramo habitados por B. gracilis como en aquel donde se le encontró en 1983 en Chisacá, Colombia (Gaviria, 1989). En este tipo de cuerpos de agua se presentan condiciones variables y extremas de temperatura con periodos de sequedad, que pueden inducir a su producción, como ocurre con algunas especies de diaptómidos (Hutchinson, 1967), pero todavía no hay evidencia de su existencia.
AGRADECIMIENTOS
Los autores expresan su agradecimiento a la Dirección de Investigaciones de la Universidad Pedagógica y Tecnológica de Colombia por el financiamiento para la realización del proyecto con código DIN No. 9902.073, del cual se produce la presente publicación. A la Corporación Autónoma Regional de Boyacá por el apoyo logístico durante la fase de campo de la investigación y a dos revisores anónimos que evaluaron el manuscrito.
BIBLIOGRAFÍA
ARMADA NACIONAL. Informe final levantamiento topográfico, batimétrico y sedimentológico laguna de Tota. Centro de Investigaciones Oceanográfico. Cartagena de Indias, Colombia. 1998.
[ Links ]BALSEIRO E. Dinámica poblacional de Boeckella gracilipes (Copepoda, Calanoida) en los lagos Andinos. Centro Regional Universitario Bariloche. UNC. Notas científicas RAL. Biol. Acuat. 1991;15:136-137.
[ Links ]BAYLY I. The non-marine Centropagidae (Copepoda: Calanoida) of the world.-Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 2, SPB Academic Publishing bv, The Hague; 1992.
[ Links ]BAYLY I, GIBSON J, WAGNER B, SWADLING K. Taxonomy, ecology and zoogeography of two East Antarctic freshwater calanoid copepod species: Boeckella poppei and Gladioferens antarcticus. Antarct. Sci. 2003;15:439-448.
[ Links ]BERMÚDEZ A, MORENO M. Estudio de la comunidad fitoplanctónica en el lago de Tota (Boyacá-Colombia) [tesis de pregrado]. Tunja: Departamento de Biología, Facultad de Ciencias Básicas, Universidad Pedagógica y Tecnológica de Colombia; 2002.
[ Links ]BLAZKA P, BRANDL Z, PROCHAZCOVA L. Oxygen consumption and ammonia and phosphate excretion in pond zooplankton. Limnol. Oceanogr. 1982;27(2):294-303.
[ Links ]BOLTOSKOY D. Colección de plancton. En: Lopretto E y Tell G, Editores. Ecosistemas de aguas continentales. Metodología para su estudio. Argentina: Ediciones Sur; 1995.
[ Links ]CARDOZO A, PITA S. Estudio de la estructura cualitativa y cuantitativa de la comunidad fitoplanctónica del lago de Tota. Sector Lago Chico (Aquitania-Boyacá) [tesis de pregrado]. Tunja: Departamento de Biología, Facultad de Ciencias Básicas, Universidad Pedagógica y Tecnológica de Colombia; 2004.
[ Links ]CORDERO RD, RUIZ JE, VARGAS EF. Determinación espacio-temporal de la concentración de fósforo en el lago de Tota. Rev. Colombiana de Química. 2005; 34(2):211-218.
[ Links ]CORPOBOYACÁ. Estudio para la identificación, delimitación, diagnóstico y priorización de ecosistemas estratégicos en jurisdicción de Corpoboyacá. Informe Final, Planificación Ambiental y Cuencas Hidrográficas, Tunja, Colombia; 1998.
[ Links ]DANIEL W. Bioestadística: base para el análisis de las ciencias de la salud. México: Editorial Limusa S.A.; 1991.
[ Links ]DAJOZ R. Tratado de ecología. Madrid: Mundi-Prensa; 1974.
[ Links ]EINSLE U. Crustacea, Copepoda, Calanoida und Cyclopoida. Süsswasser Fauna von Mitteleuropa. Stuttgart: Gustav Fischer Verlag; 1993.
[ Links ]FERNANDES C, PINTO-COELHO R. Efeito das algas Ankistrodesmus gracilis y Scenedesmus quadricauda e no indice lipidico de Daphnia laevis e Moina micrura. Acta Sci. 2000;22(2):397-401.
[ Links ]GALLO L, GAVIRIA S, RAMIREZ J. Dinámica de la comunidad zooplanctónica (excepto Protozoa) en la laguna del Parque Norte, Medellín (Antioquia, Colombia). Actual Biol. 2004;26(81):231-241.
[ Links ]GAVIRIA S. The calanoid fauna (Crustacea, Copepoda) of the Cordillera Oriental of the Colombian Andes. Hydrobiologia. 1989;178:113-134.
[ Links ]GAVIRIA S. Situación de la limnología en Colombia y sus perspectivas. Memorias Primer Encuentro de Limnólogos Iberoamericanos-Sevilla, España. 1992:61-68.
[ Links ]GAVIRIA S. Crustacean plankton of a high altitude tropical lake: Laguna de Chingaza, Colombia. Verh. Internat. Verein Limnol. 1993;25:906-911.
[ Links ]GAVIRIA S, ARANGUREN N. Especies de vida libre de la subclase Copepoda (Arthropoda, Crustacea) en aguas continentales de Colombia. Biota Colombiana. 2007;8 (1):53-68.
[ Links ]GONZÁLEZ A. Estructura poblacional de Boeckella gracilis (Daday, 1902) (Copepoda: Calanoida) en el lago de Tota, Boyacá - Colombia [tesis de pregrado]. Tunja: Departamento de Biología, Facultad de Ciencias Básicas, Universidad Pedagógica y Tecnológica de Colombia; 2005.
[ Links ]GUO X. Fauna of freshwater Copepoda in China [tesis doctoral]. Vienna: Facultad de Ciencias Formales y Naturales, Universidad de Viena; 1999.
[ Links ]GUTIÉRREZ E. Métodos estadísticos para las ciencias biológicas. Costa Rica: Editorial de la Universidad Nacional; 1995.
[ Links ]HALL CJ, BURNS CW. Hatching of Boeckella hamata (Copepoda: Calanoida) resting eggs from sediments of a tidally influenced lake. N. Z. J. Mar. Freshwater Res 2001;35:235-238.
[ Links ]HENAO A. El disco Secchi y el estado trófico. Asociación de Ingenieros Sanitarios de Antioquia. 1987;72:67-79.
[ Links ]HUTCHINSON G. A treatise on limnology. Vol. II Introduction to lake biology and the limnoplankton. New York: Editorial John Wiley & Sons Inc.; 1967.
[ Links ]HUTCHINSON G. Introducción a la ecología de poblaciones. 1ª Edición. Barcelona: Editorial Blume; 1981.
[ Links ]INFANTE A. Lakes of South America. Natural food of copepod larvae from Lake Valencia, Venezuela. Verh. Internat. Verein. Limnol. 1981;21:709-714.
[ Links ]KREBS CH. Ecología. Estudio de la distribución y la abundancia. 2ª Edición. México: Editorial Harla; 1985
[ Links ]LAMPERT W, SOMMER U. Limnoecology. The ecology of lakes and streams. New York: Oxford University Press; 1997.
[ Links ]LEWIS W. Zooplankton community analysis: studies on a tropical system. New York: Springer-Verlag; 1979.
[ Links ]LOMNIKI A. Population ecology of individuals. New Jersey: Princeton University Press. Princeton; 1988.
[ Links ]LÓPEZ C. Egg-stock and Clutch Size of Two Species of Thermocyclops Kiefer (Crustacea-Copepoda) in a Tropical Reservoir. Verh Internat Verein Limnol. 1994;25:1287-1290.
[ Links ]MAIER G. The seasonal cycle of Thermocyclops crassus (Fischer, 1853) (Copepoda: Cyclopoida) in a shallow, eutrophic lake. Hydrobiologia. 1989;178:43-58.
[ Links ]MARGALEF R. Limnología. Barcelona: Ediciones Omega S.A.; 1983.
[ Links ]MÁRQUEZ G. Ecosistemas estratégicos y otros estudios de ecología ambiental. Bogotá: Fondo FEN Colombia; 1996.
[ Links ]MENU-MARQUE S, BALSEIRO E. Boeckella antiqua (Copepoda, Calanoida, Centropagidae) from Patagonia. Hydrobiologia. 2000;429:1-7.
[ Links ]MENU-MARQUE S, LOCASCIO C. Distribución de las especies del género Boeckella (Copepoda: Calanoida: Centropagidae) en la Argentina. Physis. 1998;SB(130-131):1-10.
[ Links ]MONROY J. Variación espaciotemporal de la biomasa de los crustáceos planctónicos del lago de Tota (Boyacá-Colombia) [tesis de pregrado]. Tunja: Departamento de Biología, Facultad de Ciencias Básicas, Universidad Pedagógica y Tecnológica de Colombia; 2002.
[ Links ]MONROY J, ARANGUREN N, GAVIRIA S. Los crustáceos planctónicos del lago de Tota (Boyacá, Colombia). Ciencia en desarrollo. 2004;1(2): 64-81.
[ Links ]MORA G, TÉLLEZ L, CALA P, GUILLOT G. Estudio biológico de la ictiofauna del lago de Tota (Boyacá-Colombia), con énfasis en la Trucha Arcoiris (Oncorhynchus mykiss). Revista Academia Colombiana de Ciencias. 1992;18(70):409 - 422.
[ Links ]RINKE K, HÜBNER I, PETZOLDT T, ROLINSKI S, KÖNIG-RINKE M, POST J, LORKE A, BENNDORF J. How internal waves influence the vertical distribution of zooplankton. Freshw Biol. 2007;52: 137-144.
[ Links ]RANGEL O, AGUIRRE J. Comunidades acuáticas altoandinas -I. Vegetación sumergida y de ribera en el lago de Tota, Boyacá, Colombia. Caldasia. 1983; 65:719-742.
[ Links ]RUIZ E. Métodos para el estudio de las características físico-químicas del agua, En: Rueda G, editor. Manual de Métodos en Limnología. Bogotá: 1a edición. Asociación Colombiana de Limnología, Pen Clips Publicidad y Diseño. 2002. p. 9-14.
[ Links ]SAUNDERS J, LEWIS W. Dynamics and control mechanisms in a tropical zooplankton community (Lake Valencia, Venezuela). Ecol Monogr. 1988;58(4):337-353.
[ Links ]WETZEL R, LIKENS G. Limnological analyses. 2a Edición. New York: Ediciones Springer-Verlag; 1991.
[ Links ]
