










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.13 no.3 Bogotá Sep./Dec. 2008
ACTIVIDAD DIARIA DE COLONIZACIÓN DEL RECURSO ALIMENTICIO EN UN ENSAMBLAJE DE ESCARABAJOS COPRÓFAGOS (COLEOPTERA: SCARABAEIDAE) EN LA AMAZONÍA COLOMBIANA
Daily colonization of food resourses by Dung Beetles (Coleoptera: Scarabaeidae) in the Colombian Amazone Region
JORGE ARI NORIEGA A.1,2, ANA MARIA CUBILLOS3, CAMILO CASTAÑEDA3, ANA MARIA SANCHEZ3
1 Laboratorio de Entomología, Unidad de Ecología y Sistemática UNESIS, Facultad de Ciencias, Pontificia Universidad Javeriana. Carrera 7 No. 40-62. Bogotá, Colombia. jnorieg@hotmail.com.
2 Scarabaeinae Research Network - ScarabNet.
3 Estudiantes Biología, Pontificia Universidad Javeriana, Bogotá, Colombia.
Presentado 24 de enero de 2008, aceptado 11 de junio de 2008, correcciones 24 de junio de 2008.
RESUMEN
Los escarabajos coprófagos pertenecientes a la familia Scarabaeidae presentan una fuerte competencia por el recurso alimenticio, debido a la baja agregación espacial y al carácter efímero del mismo. Un mecanismo que puede contribuir a diluir la competencia es la segregación temporal, conduciendo a una especialización en las horas de actividad. El objetivo de este trabajo fue estudiar la actividad diaria en un ensamblaje de escarabajos coprófagos en un bosque húmedo tropical en la amazonía colombiana. Se establecieron dos transectos lineales de 225 m y en cada uno se montaron diez tram-pas de caída por 24 horas, cebadas con excremento humano de dos investigadores, colectando su contenido cada hora, en la época de sequía. Se registraron un total de 23 especies, contenidas en diez géneros y cinco tribus. La abundancia del gremio de los cavadores fue mayor que la de los otros grupos. Igualmente, los escarabajos diurnos fueron más abundantes que los crepusculares y los nocturnos. Se presentaron algunas especies con horas de actividad muy específicas, evidenciando el problema de tomar tiempos de captura muy amplios donde se pierde el detalle de la restricción horaria. La riqueza y abundancia de especies en los cebos de los dos investigadores mostró diferencias, insinuando que pueden existir variaciones dentro del excremento humano, como cebo. Finalmente, se propone que la restricción en la actividad diaria puede ser un mecanismo importante de dilución de la competencia inter e intraespecífica, que facilita a las especies la coexistencia y repartición del recurso en el tiempo.
Palabras clave: Actividad diaria, escarabajos coprófagos, ensamblaje, amazonía colombiana.
ABSTRACT
Dung beetles belonging to the Scarabaeidae family have strong competition for food resources given the low spatial aggregation and ephemeral character of those resources. Temporal segregation through specialization in hours of activity is a mechanism that can decrease that competition. The objective of this paper is study the daily activity of an assemblage of dung beetles in a tropical wet forest in the colombian amazon. During the dry season, two linear transects of 225 m were established, with 10 pitfall traps mounted for 24 hours, baited with human excrement of two researchers. Trap contents were collected hourly. 23 total species were recorded, within 10 genera and 5 tribes. The abundance of the diggers guild were higher then the others groups. Diurnal beetles were more abundant than nocturnal and crepuscular species. Some species with very specific hours of activity were present, evidencing the problem of allowing a long time between collections, thus losing the detail of restricted activity. Differences existed in the abundance and richness of species on the bait, implying that there can be variations between human excrement. Finally, it was shown that restriction in daily activity can be an important mechanism for diminishing inter and intra specific competition, which facilitates the coexistence of species and the division over the resource in time.
Key words: Daily activity, dung beetles, assemblage, colombian amazon.
INTRODUCCIÓN
Todos los organismos poseen un mecanismo de tipo fisiológico para medir el tiempo, conocido como "reloj biológico". La expresión más común de este mecanismo es el ritmo circadiano, o la capacidad para determinar y repetir funciones a intervalos de tiempo aproximadamente diarios, aun en ausencia total de algún referente externo de esta periodicidad como lo es la luz (Odum, 1972; Stumpf, 1986; Verdú et al., 2004). La existencia de un reloj biológico se constituye en una gran ventaja para las especies al poder integrar los ritmos ambientales y fisiológicos, permitiéndoles generar patrones de periodicidad en sus actividades (Koskela, 1979). La actividad diaria de un ensamblaje es la combinación de los ritmos (sincrónicos y/o asincrónicos) de todas las especies que lo componen, sumando las actividades de tendencia diurna, crepuscular y nocturna (Odum, 1972).
La estructura temporal se ve afectada por una serie de variables dentro de las cuales cabe destacar principalmente las restricciones térmicas, la disponibilidad y utilización del recurso alimenticio, las fluctuaciones en los niveles de humedad, las estrategias antidepredatorias y los mecanismos de repartición para evitar la competencia interespecífica (Doube, 1983; Houston y Mcintyre, 1985; Mena et al., 1989). La actividad diaria en el caso específico de organismos ectotermos como los insectos, está fuertemente relacionada con el mecanismo de termorregulación, donde los individuos presentan rangos de actividad cuando la temperatura (ambiental y corporal) sea la más adecuada (Verdú et al., 2004). Uno de los grupos de insectos donde este tipo de patrones en la actividad pueden estudiarse fácilmente, debido a lo sencillo de su muestreo y a la existencia de una estrecha relación ecológica con las variables que los condicionan, son los escarabajos coprófagos de la familia Scarabaeidae. En las regiones neotropicales esta familia juega un importante papel al interior de los ecosistemas, ya que ayuda en el proceso de reciclaje de nutrientes al descomponer el excremento de mamíferos y otros vertebrados (Halffter y Halffter, 1989; Gill, 1991). Este proceso es realizado por medio de la relocalización del excremento, para lo cual existen tres estrategias comportamentales que obedecen a la forma como los individuos toman el excremento y lo relocalizan: los cavadores (paracópridos) hacen galerías y llevan parte del excremento hacia abajo, los rodadores (telecópridos) hacen bolas con el excremento y las alejan rodándolas y los residentes (endocópridos) viven dentro del excremento o en la interfase excremento-suelo (Halffter y Matthews, 1966; Hanski, 1990).
A pesar de las facilidades que brinda este grupo son muy pocos los trabajos que han estudiado la variación de la actividad diaria, ya que la mayoría han tomado periodos de 24 o 48 horas en donde se diluyen por completo las variaciones especificas (Davis, 1993; Escobar, 1997; Davis et al., 2003; Boonrotpong et al., 2004; Andresen, 2005). Para intentar recuperar alguna fracción de esta actividad algunos muestreos se han realizado cada 12 horas, dividiendo el ensamblaje en especies diurnas y nocturnas, pero esta división es sumamente arbitraria (Halffter et al., 1992; Kirk, 1992; Montes de Oca y Halffter, 1995; Halffter y Arellano, 2002; Hernandez et al., 2003; Escobar, 2004). Otros estudios han tomado rangos de seis o de cuatro horas (Fincher et al., 1971; Beerwinkle y Fincher, 1980; Peck y Forsyth, 1982). Estudios en laboratorio han demostrando que la actividad evaluada en campo utilizando trampas de caída es solo una fracción de la misma, correspondiente al momento de la colonización del recurso alimenticio (Martin-Piera et al., 1994). Adicionalmente se ha evidenciado que existen patrones de actividad muy marcados, reconociéndose géneros y especies típicamente diurnas, crepusculares o nocturnas. De igual manera existen especies generalistas así como otras que muestran periodos de actividad muy limitados, que pueden llegar a ho-ras específicas (Estrada et al., 1993; Noriega, 2001; Noriega, 2002). Son muy pocos los estudios existentes acerca de los patrones de actividad diaria de colonización del recurso alimenticio por parte de escarabajos coprófagos (Davis, 1999; Feer y Pincebourde, 2005), y hasta este trabajo no se han realizado acercamientos en el departamento del Amazonas. A esto hay que sumar que los inventarios en la amazonia colombiana son muy pobres (Howden y Nealis, 1975; Quintero, 1998; Castro, 2001). Es por lo anterior que este trabajo no solo es un primer acercamiento al entendimiento de los patrones de actividad, sino que además hace una importante contribución al conocimiento de la entomofauna de esta región del país.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
El estudio se realizó en la Comunidad Monilla Amena (Resguardo Indígena Ticuna-Uitoto), ubicado a 71 msnm en las coordenadas geográficas 4°06'46,25" Sur -69°55'52,11" Oeste, a 9,5 km de la ciudad de Leticia, departamento del Amazonas, Colombia. La zona presenta un ciclo monomodal, la época seca es entre junio y agosto. El muestreo se desarrolló a finales de la época seca en 2005 en el hábitat de Bosque de Tierra Firme (BTF).
MUESTREO DE LOS ESCARABAJOS
Para este estudio se utilizó el tipo de trampa de caída propuesto por Noriega, 2001, el cual consiste en un vaso de 600 mL enterrado a nivel del suelo, con un embudo cortado en su extremo para facilitar la entrada de los especimenes y un alambre el cual sujeta una pequeña copa (25 mL) que se coloca 10 cm arriba de la boca del vaso y en ella se deposita el cebo. Se realizaron dos transectos, cada uno con diez trampas de caída. La distancia entre trampas fue de 25 m y entre los transectos de 100 m. En cada trampa se colocaron 25 mL de excremento humano fresco. Las trampas del transecto 1 fueron cebadas con el excremento del investigador A y las del transecto 2 con el excremento del investigador B.
Las trampas fueron abiertas a las 12:00 horas del día 5 de octubre y fueron cerradas finalmente a las 12:00 del día siguiente, transcurriendo un tiempo de 24 horas. Se realizaron dos colectas del contenido total de todas las trampas a las 15:00 y 16:00 horas del 5 de octubre (transcurrida una hora y dos horas) y otras tres adicionales a las 9:00, 10:00 y 11:00 horas del 6 de octubre (transcurridas 19, 20 y 21 horas desde su apertura). En cada hora de muestreo, el contenido de cada trampa, fue guardado independientemente, con los datos del transecto, trampa y hora de muestreo. Los ejemplares colectados fueron sacrificados en etanol al 70%.
FASE DE LABORATORIO
En laboratorio se identificaron los ejemplares colectados hasta el nivel de especie, usando claves taxonómicas para este grupo y el material de referencia de la Colección Entomológica del Museo de Historia Natural de la Pontificia Universidad Javeriana, Bogotá D.C. (MPUJ); la Colección de Insectos del Instituto de Investigaciones Alexander von Humboldt, Villa de Leyva (IAvH) y la Colección de referencia de Jorge Ari Noriega, Bogotá D.C. (CJAN). Posteriormente el material fue montado y etiquetado, para ser depositado en el MPUJ.
ANÁLISIS DE RESULTADOS
Se obtuvo la riqueza y abundancia de las especies colectadas y se realizó una curva de acumulación utilizando el programa EstimateS v.7.5 (Colwell, 1997). Se evaluó la homogeneidad y normalidad de varianza, y se realizó una prueba de Kruskall Wallis para todas las comparaciones de abundancia, mediante el Software Statisca 7.0. Por último, se comparó la diferencia en la abundancia y riqueza entre los dos transectos, a partir de una prueba de bondad de ajuste con X2.
RESULTADOS
RIQUEZA Y COMPOSICIÓN
Se colectaron un total de 244 individuos, correspondientes a 23 especies contenidas en diez géneros y cinco tribus (Tabla.1). La especie más abundante en el muestreo fue Canthon aequinoctialis (73 ind., 29,9%), seguida por Onthophagus haematopus (48 ind., 19,6%) y Canthon luteicollis (22 ind., 9,01%). Nueve especies que están representadas por un solo individuo.
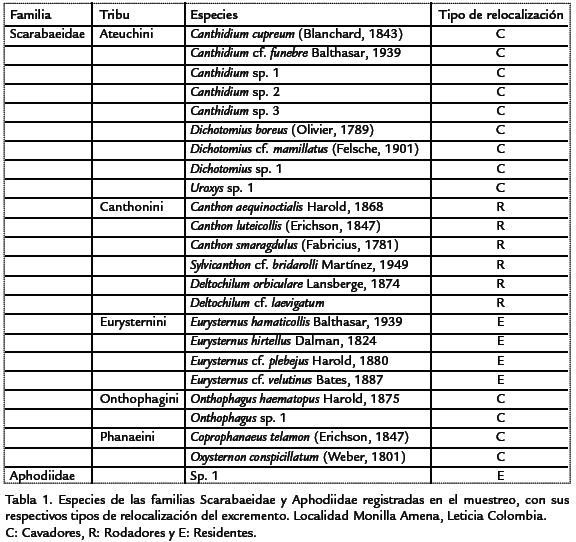
De las 43 especies registradas para el departamento del Amazonas (Medina et al., 2001), se registraron únicamente 11 (25,6%), seis adicionales están en proceso de confirmación. Las restantes especies C. aequinoctialis, Dichotomius boreus, Deltochilum orbiculare, O. haematopus y Deltochilum cf. laevigatum son registros nuevos para el departamento, así como Scybalocanthon cf. bridarolli y Uroxys sp. 1 son nuevos registros a nivel genérico para esta región, ya que no estaba registrada ninguna especie de estos géneros en esta zona del país (Medina et al., 2001). Adicionalmente se registró la presencia de cinco individuos de la familia Aphodiidae, asociados únicamente a los muestreos de la mañana, los cuales se encuentran en proceso de identificación por especialistas.
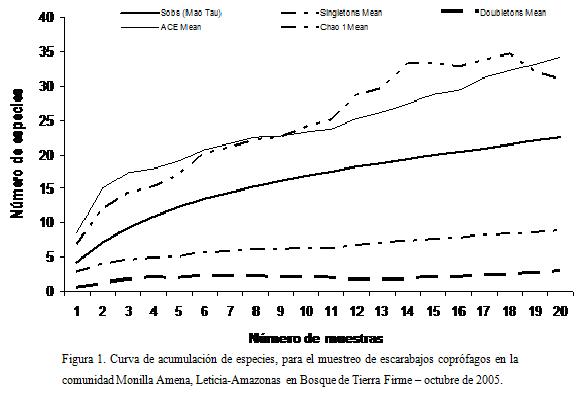
La curva de acumulación de especies, arrojó un muestreo moderadamente eficiente, con un rango entre el 64,5% - 58,8% de las especies esperadas por los diferentes estimadores (Fig. 1). La curva de las especies observadas no llega a una asintota, aunque las especies que solo aparecen con dos individuos parecieran disminuir.
Con respecto al tipo de relocalización del recurso, el ensamblaje tiene una dominancia de los cavadores, presentando 13 especies (56,5%) con este hábito, seguidos delos rodadores con seis (26,1%) y solo cuatro especies (17,4%) residentes. Se encontraron un total de 144 individuos pertenecientes al grupo de los cavadores (59,0%), 72 rodadores (29,5%) y 28 residentes (11,5%). Al comparar la abundancia de los gremios de escarabajos coprófagos no se encontraron diferencias significativas entre ellos (Kruskal-Wallis p=0,5366, α=0,05).
ACTIVIDAD DIARIA
De los 24 muestreos a lo largo de un día completo de actividad, se obtuvieron datos completos de dos muestreos de la tarde del 5 de octubre y dos muestreos de la mañana del 6 de octubre, donde se puede garantizar con certeza que los individuos presentes en las trampas se vieron atraídos a esas horas en particular, ya que sus rangos de búsqueda de alimento coincidieron con las horas de apertura y colecta. El muestreo de las 9:00 horas contiene información de un lapso de tiempo de 17 horas, incluyendo individuos con actividad crepuscular tarde, nocturna y crepuscular mañana, por lo que esta información es poco confiable y no se utilizó para el análisis estadístico.
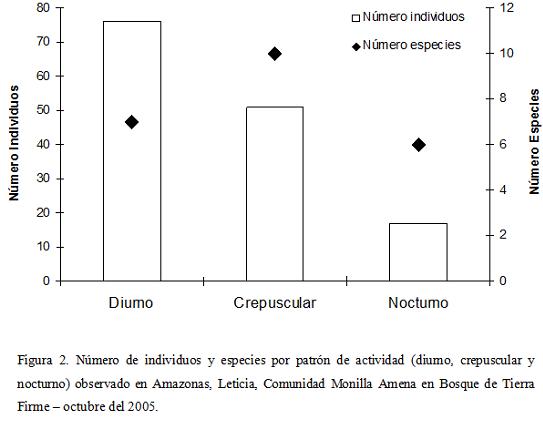
Con respecto a los diferentes patrones de actividad (diurnos, nocturnos y crepusculares) se pudo determinar que existe una predominancia en la actividad diurna con un total de 116 individuos (47,5%), seguida por los crepusculares con 81 individuos (33,2%) y los nocturnos con solo 47 individuos (19,3%). Sin embargo, al referirse al número de especies, la variación entre los patrones de actividad no es tan marcada, presentando mayor cantidad de especies el grupo de los crepusculares (diez spp., 43,5%), seguido del grupo de los diurnos (siete spp., 30,4%) y por último los nocturnos (seis spp., 26,1%). Es interesante anotar que no se presenta una estrecha relación entre el número de especies y la abundancia de las mismas, según el patrón de actividad (Fig. 2).
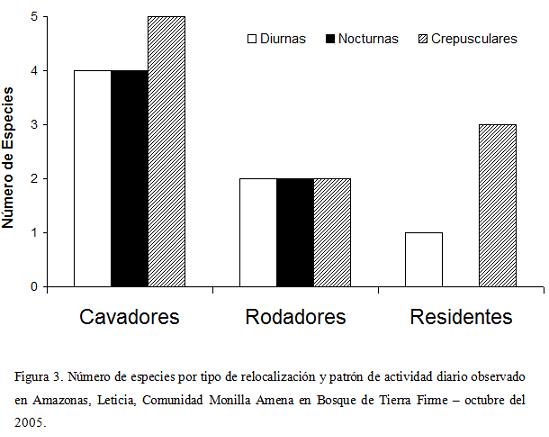
Al analizar los tipos de relocalización del recurso alimenticio y los patrones de actividad encontramos que para el grupo de los cavadores se observa el mismo número de especies de actividad diurna y nocturna, con una pequeña dominancia de los crepusculares. Algo semejante sucede para el caso de los rodadores, grupo con igual número de especies en cada uno de los tres grupos. El grupo de los endocópridos rompe con esta relación, no solo por no presentar especies de actividad nocturna, sino por evidenciar una dominancia de las especies crepusculares sobre las diurnas (Fig. 3). Al comparar las abundancias de los gremios de escarabajos coprófagos según las horas de actividad no se encontraron diferencias significativas (Kruskal-Wallis p=0,6059, α=0,05).
El patrón de actividad observado en los cuatro muestreos, evidencia que el mayor número de individuos por trampa se presentó a las 16:00 para el transecto 2. A pesar de ello el máximo número de individuos en conjunto lo presentó el muestreo de las 10:00 horas, seguido por el muestreo de las 15:00 (Fig. 4). En general se obtuvo un mayor número de individuos por la tarde que por la mañana aunque este valor no es significativamente diferente (Kruskal-Wallis p=0,4847, α=0,05).
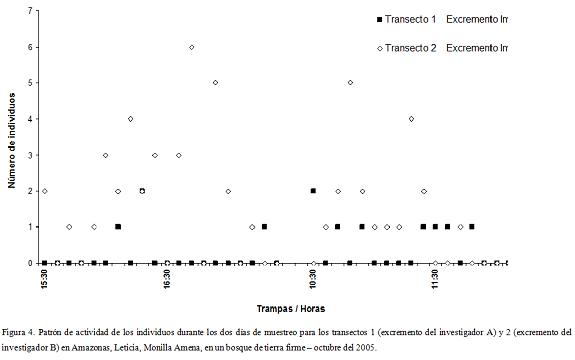
Al contrastar el número de individuos entre los dos transectos, teniendo en cuenta que se utilizaron cebos de dos investigadores diferentes, se encontraron diferencias significativas (Kruskal-Wallis p=0,0430, α=0,05) presentando el transecto 2 una mayor abundancia que el 1. La riqueza en cambio fue semejante entre los dos excrementos utilizados.
DISCUSIÓN Y CONCLUSIONES
RIQUEZA Y COMPOSICIÓN
Al comparar el número de especies encontradas con las especies registradas para el Amazonas (Medina et al., 2001), se evidencia una baja representatividad, la cual está compensada con la existencia de cinco nuevos registros específicos y dos nuevos registros a nivel genérico para el departamento. El bajo número de trampas, el corto tiempo de muestreo y la dependencia espacial de las unidades de muestreo, hacen de este estudio una excelente herramienta de análisis hacia futuros trabajos. Las curvas de acumulación advierten de la necesidad de incrementar el número de unidades de muestreo, estimando la existencia de una riqueza de más de 40 especies para la zona, aunque este número según algunos trabajos recientes puede ser aun mucho mayor (Noriega y Quintero, 2008). El haber realizado un muestreo en un solo hábitat, en una única estación climática y con un solo tipo de cebo, sin incluir variables como microhabitats, estratificación del bosque y ecotonos, son sin duda limitaciones que afectan directamente en la baja riqueza encontrada. A pesar de lo anterior es muy posible que verificando las especies cuya identificación taxonómica está en proceso, aparezcan más registros nuevos para esta localidad, demostrando lo poco muestreada que se encuentra esta zona del país.
ACTIVIDAD DIARIA
Aunque muchos autores como Feer y Pincebourde, 2005, establecen la existencia de tres patrones: diurno, crepuscular y nocturno, los datos obtenidos en este estudio ratifican otros resultados a nivel nacional (Noriega, 2001; Noriega, 2002; Cultid, 2006) donde se insinúa que estos patrones pueden ser mucho más estrechos y que al interior de cada una de estas agrupaciones artificiales, existen rangos más específicos y naturales de actividad diaria. La predominancia de especies diurnas, puede estar fuertemente relacionada con los picos de actividad de producción del excremento (Gill, 1991; Davis, 1999), aunque algunos autores han planteado que este pico coincide más con las horas crepusculares, ya que los mamíferos producen una mayor cantidad de excremento durante este periodo (Howden y Young, 1981; Julliot, 1997; Feer y Pincebourde, 2005).
Los datos encontrados en este estudio coinciden con los reportes de Feer y Pincebourde, 2005, para la Guyana Francesa, en donde las especies nocturnas son el grupo menos dominante. A pesar de esto, la existencia de otros valores muy diferentes obtenidos en otras localidades, indican que existen variaciones geográficas muy marcadas. La mayoría de los escarabajos en Australia presentan una actividad de tipo nocturna (Hill, 1996), mientras en Asia la actividad es más de tipo diurna (Hanski y Krikken, 1991). Según Hill, 1996, existen más especies con patrones de actividad crepuscular, que diurna o nocturna, lo cual coincide con los resultados encontrados para esta localidad. La mayor cantidad de especies cavadoras, puede estar relacionada con la abundancia del recurso y con características ecológicas específicas de la localidad que hacen que ser cavador sea más exitoso. Aunque algunos autores han encontrado que los telecópridos son más dominantes en el día y los paracópridos y endocópridos en la noche (Krell et al., 2003), para este estudio no se encontraron diferencias significativas. Es muy interesante el caso de los endocópridos quienes muestran una cierta afinidad a una actividad de tipo crepuscular, lo cual puede tener mucha relación con la cantidad de excremento que necesitan (Halffter y Edmonds, 1982). Al hacer sus galerías al interior del mismo, los endocópridos podrían tener más éxito cuando el recurso es más abundante, agregado y la presión de temperatura es menor, coincidiendo de nuevo con la hipótesis del pico de producción de excremento para los bosques neotropicales (Howden y Young, 1981; Gill, 1991).
A raíz de la variación encontrada entre los cebos de los dos investigadores, se propone realizar un estudio con un mayor número de replicas para comparar entre diferentes excrementos de distintas personas, para de esta manera analizar si existen diferencias importantes en cuanto a la abundancia y riqueza de especies atraídas, lo cual podría afectar radicalmente los protocolos de muestreo. Sin duda, la segregación espacial de la actividad en patrones establecidos, es un mecanismo importante de dilución de la competencia interespecífica (Tokeshi, 1999), al evitar que determinadas especies se encuentren en el recurso al mismo tiempo. Es posible que estos patrones encontrados se vean igualmente afectados por la época del año y más particularmente por la estacionalidad, como se ha demostrado para otras localidades (Doube, 1991). Finalmente, se puede concluir que existen patrones de actividad diarios muy marcados entre las especies que estructuran un ensamblaje, lo cual juega un papel fundamental a la hora de mantener una alta diversidad. Es por esto que se hace prioritario entender estos patrones de repartición temporal con miras a entender procesos ecológicos y evolutivos, que no solo nos permitan conocer nuestra biodiversidad sino poder llegar a protegerla.
AGRADECIMIENTOS
A la comunidad indígena Monilla Amena por toda su hospitalidad y a Giovanny Fagua del Laboratorio de Entomología de la Pontificia Universidad Javeriana. A Luisa Fernanda Escobar y Carlos Rivera en la accesoria estadística. A Fernando Vaz de Mello, Ricardo Botero-Trujillo y a dos revisores anónimos por los aportes al manuscrito. A ScarabNet quien a contribuido a desarrollar y mejorar los mecanismos de comunicación y ayuda entre investigadores. Este trabajo se realizó bajo el convenio No. 1662, entre CorpoAmazonía, Comunidad Monilla Amena y Pontificia Universidad Javeriana.
BIBLIOGRAFÍA
ANDRESEN E. Effects of season and vegetation type on community organization of dung beetles in a tropical dry forest. Biotropica. 2005;37(2):291-300.
[ Links ]BEERWINKLE KR, FINCHER GT. Automatic trap for determining hourly flight activity of dung beetles. Southwest Entomol. 1980;5:107-111.
[ Links ]BOONROTPONG S, SOTTHIBANDHU S, PHOLPUNTHIN C. Species composition of dung beetles in the primary and secondary forests at Ton Nga Chang wildlife sanctuary. ScienceAsia. 2004;30:59-65.
[ Links ]CASTRO DJ. Estudio del estado de la comunidad de escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae y Coprinae) en áreas de colonización de la selva amazónica, Departamento de Guaviare, Colombia [Trabajo de Grado]. Universidad Nacional de Colombia. Facultad de Ciencias. Departamento de Biología; 2001.
[ Links ]COLWELL RK. EstimateS. Statistical estimation of species richness and shared species samples. 1997. http://viceroy.eeb.uconn.edu/estimates.
[ Links ]CULTID CA. Ritmos de actividad diaria de una comunidad de coleópteros coprófagos (Scarabaeidae: Scarabaeinae) en un paisaje andino, Risaralda, Colombia [Trabajo de Grado]. Universidad del Valle, Cali, Colombia; 2006.
[ Links ]DAVIS ALV. Alpha-diversity patterns of dung beetle assemblages (Coleoptera: Scarabaeidae, Aphodiidae, Staphylinidae, Histeridae, Hydrophilidae) in the winter rainfall region of South Africa. Afr Entomol. 1993;1(1):67-80.
[ Links ]DAVIS AJ. Species packing in tropical forest: diel flight activity of rainforest dung-feeding beetles (Coleoptera: Aphodiidae, Scarabaeidae, Hybosoridae) in Borneo. Raffles Bull Zool. 1999;47:473-486
[ Links ]DAVIS AL, AARDE RJ, SCHOLTZ CH, DELPORT JH. Convergence between dung beetle assemblages of a post-mining vegetational chronosequence and unmined dune forest. Restoration Ecology. 2003;11(1):29-42.
[ Links ]DOUBE BM. The habitat preference of some bovine dung beetles (Coleoptera, Scarabaeidae) in Hluhluwe Game Reserve. South Africa Bull Entomol Res. 1983;73:357-371.
[ Links ]DOUBE BM. Dung beetles of Southern Africa. En: Hanski I., Cambefort Y., editores. Dung Beetle Ecology. Princeton University Press, Princeton; 1991. p. 133-135.
[ Links ]ESCOBAR F. Estudio de la comunidad de coleópteros coprófagos (Scarabaeidae) en un remanente de bosque seco al norte del Tolima, Colombia. Caldasia. 1997; 19(3):419-430.
[ Links ]ESCOBAR F. Diversity and composition of dung beetle (Scarabaeinae) assemblages in a heterogeneous Andean landscape. Tropical Zoology. 2004;17:123-136.
[ Links ]ESTRADA A, HALFFTER G, COATES-ESTRADA R, MERRITT AJ. Dung beetles attracted to mammalian herbivore (Alouatta palliata) and omnivore (Nasua narica) dung in the tropical rain forest of Los Tuxtlas, Mexico. J Trop Ecol. 1993;9:45-54.
[ Links ]FEER F, PINCEBOURDE S. Diel flight and ecological segregation within an assemblage of tropical forest dung and carrion beetles. J Trop Ecol. 2005;21:21-30.
[ Links ]FINCHER GT, DAVIS R, STEWART TB. Flight activity of coprophagous beetles on a swine pasture. Ann Entomol Soc Am. 1971;64(4):855-860.
[ Links ]GILL BD. Dung beetles in tropical American forest. En: Hanski I, Cambefort Y, Eds. Dung Beetle Ecology. Princeton University Press, Princeton; 1991. p. 211-229.
[ Links ]HALFFTER G, ARELLANO L. Response of dung beetle diversity to human-induced changes in a tropical landscape. Biotropica. 2002;34(1):144.154.
[ Links ]HALFFTER G, EDMONDS WD. The nesting behavior of dung beetles (Scarabaeinae): an ecological and evolutive approach. Instituto de Ecología, Publicación No. 10, México, D.F. 1982.
[ Links ]HALFFTER G, FAVILA ME, HALFFTER V. A comparative study of the structure of the scarab guild in mexican tropical rain forests and derived ecosystems. Folia Entomologica Mexicana. 1992;(84):131-156.
[ Links ]HALFFTER G, HALFFTER V. Behavioral evolution of the nonrolling roller beetles. Instituto de Ecología, México; 1989;32:1-53.
[ Links ]HALFFTER G, MATTHEWS EG. The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Folia Entomológica Mexicana. 1966;12-14:1-312.
[ Links ]HANSKI I. Dung and carrion insects. In: Shorrocks B, Swingland I. Eds. Living in a patchy environment. Oxford Press, N.Y.; 1990. p. 127-145.
[ Links ]HANSKI I, KRIKKEN J. Dung beetles in tropical forest in South East Asia. En: Hanski I, Cambefort Y, Eds. Dung Beetle Ecology. Princeton University Press, New Jersey; 1991. p. 179-197.
[ Links ]HERNANDEZ B, MAES JM, HARVEY CA, VILCHEZ S, MEDINA A, SANCHEZ D. Abundancia y diversidad de escarabajos coprófagos y mariposas diurnas en un paisaje ganadero en el departamento de Rivas, Nicaragua. Agroforesteria en las Américas. 2003;10(39-40):93-102.
[ Links ]HILL CJ. Habitat specificity and food preferences of an assemblage of tropical Australian dung beetles. J. Trop. Ecol. 1996;12:449-460
[ Links ]HOWDEN HF, NEALIS VG. Effects of clearing in the tropical rain forest on the composition of the coprophagous scarab beetle fauna (Coleoptera). Biotropica. 1975;7(2):77-83.
[ Links ]HOWDEN HF, YOUNG OP. Panamanian Scarabaeinae. Contribution to the American Entomological Instituted (Ann Arbor); 1981;18:1-204.
[ Links ]HOUSTON KW, MCINTYRE P. The daily onset of flight in the crepuscular dung beetle Onitis alexis. Entomol Exp Appl. 1985;39:223-232.
[ Links ]JULLIOT C. Impact of seed dispersal by red howler monkeys (Alouatta seniculus) on the seedling population in the understorey of tropical rainforest. J Ecol. 1997;85: 431-440.
[ Links ]KIRK AA. Dung beetles (Coleoptera: Scarabaeidae) active in patchy forest and pasture habitats in Santa Cruz Province, Bolivia, during spring. Folia Entomologica Mexicana. 1992;(84):45-54.
[ Links ]KOSKELA H. Patterns of diel flight activity in dung inhabiting beetles: an ecological analysis. Oikos. 1979;33:419-439.
[ Links ]KRELL FT, KRELL-WESTERWALBESLOH S, WEISS I, EGGLETON P, LINSENMAIR K E. Spatial separation of Afrotropical dung beetle guilds: a trade off between competitive superiority and energetic constraints (Coleoptera: Scarabaeidae). Ecography. 2003;26:210-222.
[ Links ]MARTIN-PIERA F, SANMARTIN I, LOBO JM. Observaciones sobre el ritmo de actividad diaria en escarabaeidos telecópridos (Coleoptera, Scarabaeidae). Bull Soc Entomol France. 1994;99(5):463-470.
[ Links ]MEDINA CA, LOPERA AT, VITOLO A, GILL B. Escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) de Colombia. Biota Colombiana. 2001;2(2):131-144.
[ Links ]MENA J, GALANTE E, LUMBRERAS CJ. Daily flight activity of Scarabaeidae and Geotrupidae (Col.) and analysis of the factors determining this activity. Ecología Mediterránea. 1989;XV(1/2):69-80.
[ Links ]MONTES DE OCA ET, HALFFTER G. Daily and seasonal activities of a guild of the coprophagous, burrowing beetle (Coleoptera Scarabaeidae Scarabaeinae) in tropical grassland. Tropical Zoology. 1995;8:159-180.
[ Links ]NORIEGA JA. Estudio de la actividad diaria de colonización del recurso alimenticio, en una comunidad de escarabajos coprófagos (Coleoptera: Scarabaeidae), a lo largo de un gradiente altitudinal en la Sierra Nevada de Santa Marta [Tesis de Grado]. Bogotá: Departamento de Ciencias Biológicas, Universidad de Los Andes; 2001. p. 112.
[ Links ]NORIEGA JA. Aportes a la biología del escarabajo suramericano Sulcophanaeus leander (Coleoptera: Scarabaeidae). Acta Zool Mex. 2002;87:67-82.
[ Links ]NORIEGA JA, QUINTERO I. Escarabajos Coprófagos (Coleoptera: Scarabaeidae) de la Provincia de la Amazonía. Biota Colombiana (en prensa), 2008.
[ Links ]ODUM EP. Ecología. Ed. Interamericana, 3ª edición, México, 1972. p. 639.
[ Links ]PECK SB, FORSYTH A. Composition, structure, and competitive behaviour in a guild of Ecuadorian rain forest dung beetles (Coleoptera; Scarabaeidae). Can J Zool. 1982;60:1624-1634.
[ Links ]QUINTERO SI. Composición, diversidad y preferencias por recurso alimenticio en una comunidad de escarabajos coprófagos en una área de selva amazónica en Leticia-Amazonas-Colombia (Coleoptera: Scarabaeidae: Scarabaeinae). Tesis de Grado, Universidad Pedagógica Nacional, Santafé de Bogotá; 1998.
[ Links ]STUMPF IVK. Estudo da Fauna de escarabeídos em Mandirituba, Paraná, Brasil. Acta Biologica Paranense. 1986;15:125-153.
[ Links ]TOKESHI M. Species coexistence: ecological and evolutionary perspectives. Blackwell Science, Oxford; 1991. p. 454.
[ Links ]VERDÚ JR, DIAZ A, GALANTE E. Thermoregulatory strategies in two closely related sympatric Scarabaeus species (Coleoptera: Scarabaeinae). Physiol Entomol. 2004;29:32-38.
[ Links ]
