










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.13 no.3 Bogotá Sep./Dec. 2008
DESCRIPCIÓN MORFOLÓGICA DEL TUBO DIGESTIVO DE JUVENILES DE RUBIO Salminus affinis (PISCES: CHARACIDAE)
Alimentary tract of juvenile Rubio Salminus affinis (Pisces: Characidae) morphological description
ATENCIO GARCÍA VJ1, HERNÁNDEZ MUÑOZ J1, PARDO CARRASCO SC2*1 FMVZ/DCA/CINPIC, Universidad de Córdoba, Carrera 6 No. 76-103, Montería, Colombia.
2 FCA/DPA, Universidad Nacional de Colombia, Sede Medellín, Calle 59A No. 63-020, Medellín, Colombia. *Autor correspondencia: scpardoc@unal.edu.co, Calle 59 A #63-020 Bloque 50 oficina 313 Departamento de Producción Animal, Universidad Nacional de Colombia, Sede Medellín, Colombia.
Presentado 17 de mayo de 2008, aceptado 20 de junio de 2008, correcciones 14 de agosto de 2008.
RESUMEN
El objetivo del estudio fue la descripción del tubo digestivo de juveniles de Rubio Salminus affinis. Se analizaron cinco juveniles con 23,6±2,6 cm de longitud total y 142,8±62,5 g de peso. Se realizó descripción topográfica y morfológica de los órganos y cortes histológicos con tinción H-E. El esófago es un órgano tubular corto de pared gruesa que representa el 7,8% de la longitud total del tubo digestivo, presenta pliegues internos que ofrecen gran capacidad de distensión permitiendo el paso de presas de gran tamaño. El estómago es una bolsa asimétrica en forma de "Y", grande y musculosa, amplio en la porción anterior y estrecho hacia el fondo; el número de ciegos pilóricos varió entre 13 y 23 pudiendo ser monotubulares y ramificados. El intestino, con tres asas, se extiende desde el esfínter pilórico hasta el ano. Histológicamente el tubo digestivo está formado por cuatro capas: mucosa, submucosa, muscular y serosa; el epitelio de revestimiento del esófago es estratificado plano no queratinizado con células caliciformes, cambiando a simple cilíndrico mucosecretor llegando al estómago; en el estómago se encuentra epitelio simple cilíndrico mucosecretor y los ciegos pilóricos e intestino son revestidos por epitelio simple cilíndrico con células caliciformes. Tanto el valor promedio del coeficiente intestinal (0,58), como la pared distensible del estómago sugieren que el Rubio tiene un tubo digestivo propio de carnívoro y sus características histológicas presentan similitud con la mayoría de vertebrados superiores.
Palabras clave: anatomía, histología, intestino, peces, río Sinú
ABSTRACT
The aim of work was giving biological information about Salminus affinis,a morphological description of the alimentary tract was carried out. Five juvenile Rubio were analyzed (23.6±2.6 cm of total length and 142.8±62.5 g of weight). Topographical and morphological description organs and histology with H-E stain were carried out.
The esophagus is a short tubular thick-walled organ that represents 7.8% of the alimentary tract total length; it presents internal folds that offer great distension capacity allowing the step of great volume of bit meat. The stomach is an asymmetric bag in "Y" form, big and muscular, wide in anterior part and narrow toward the bottom; the number of pyloric cecum can vary between 13 and 23 and could be monotubular and branching. The intestine, with three loops, extends from the pyloric sphincter until the anus. Histologycally alimentary tract is formed by four layers: mucous, submucous, muscular and serous; the epithelium lining of the esophagus is stratified plane non keratinized with chaliced cells changing to simple cylindrical mucosecretor up to stomach; in the stomach epithelium simple cylindrical mucosecretor are found and the pyloric and intestine cecum are covered by cylindrical simple epithelium with chaliced cells. So the middle value of the intestinal coefficient (0.58), as the distensible wall of the stomach suggest that Rubio has an alimentary tract of carnivorous and its histologycal characteristics are similar with most of superior vertebrates.
Key words: anatomy, fish, histology, intestine, Sinú river.
INTRODUCCIÓN
Los peces tropicales constituyen casi el 75% del número total de especies de peces; a pesar de esta dominancia es muy poco lo conocido acerca de ellos. Exhiben una enorme diversidad a todos los niveles de organización biológica, desde morfología, comportamiento y la forma de sus patrones de coloración hasta sus habilidades fisiológicas para adaptarse a las desafiantes condiciones ambientales (Brauner y Val, 2006).
La diversidad de peces reportada en las cuencas colombianas se estimó en 838 especies (Mojica, 1999); sin embargo el conocimiento que se tiene de los aspectos básicos de su biología en la mayoría de los casos es incompleto o inexistente, generando en ocasiones equívocos cuando se trata de desarrollar tecnologías de cultivo (Mejía y Navarro, 2006). Una de estas especies es el Rubio Salminus affinis; el cual se creía endémico de Colombia, donde su distribución se ha registrado en las cuencas de los ríos Magdalena, San Jorge, Sinú, Cauca, Cesar y Ranchería (Miles, 1971; Dahl, 1971; Mojica et al., 2002); pero recientemente fue registrado en Ecuador en el río Santiago (Lima et al., 2003). Mojica et al., 2002, lo declararon como vulnerable a la extinción en la cuenca del Magdalena por la fuerte presión pesquera que ha sufrido y alertaron sobre su situación en la cuenca del río Sinú debido a los cambios introducidos por la construcción y operación de la hidroeléctrica Urrá; además recomendaron con urgencia investigar la biología básica de esta especie. Esta situación ha generado interés para estudiar este pez reofílico, con importancia en la pesca deportiva (Dahl, 1971; Martínez, 1981) y que podría tener interés para la piscicultura. Se destacan estudios sobre los parásitos que lo infectan en los ríos Sinú y San Jorge (Pardo et al., 2007) y hematología y química sanguínea (Atencio García et al., 2007); pero se desconocen aspectos morfológicos (macroscópica y microscópica) de su tubo digestivo, lo cual es fundamental para construir el conocimiento alrededor de la nutrición en cautiverio (Seixas-Filho et al., 2001).
El interés por estudiar el aparato digestivo de los peces suramericanos de interés comercial es reciente; destacándose los estudios en Piaractus brachypomus (Eslava-Mocha et al., 2000), Brycon orbignyanus (Seixas-Filho et al., 2000), Pseudoplatystoma coruscans (Seixas-Filho et al., 2001) y Brycon amazonicus (Eslava-Mocha et al., 2001), que permiten ver diferencias anatómicas y funcionales asociadas con los hábitos alimentarios (Sánchez et al., 2003). El objetivo del estudio fue describir macroscópica y microscópicamente el tubo digestivo de Rubio, lo cual aportará información básica para el desarrollo de tecnologías de producción en cautiverio.
MÉTODOS
La descripción morfológica del tubo digestivo se realizó en cinco ejemplares juveniles de Rubio de 23,6±2,6 cm de longitud total y 142,8±62,5 g de peso, capturados por pescadores en la parte baja del río Sinú entre Lorica y San Bernardo del Viento (Córdoba, Colombia). Los animales fueron fijados inmediatamente en formol al 3,7% (neutralizado con fosfato monohidratado de sodio 4 g/L y fosfato dibásico de sodio 6,5 g/L).
Además, se realizaron perfusiones con el fijador vía oral y anal en el sitio de captura para mayor fijación de la mucosa del tubo digestivo y mejor conservación de los órganos internos (Seixas Filho et al., 2000); luego los ejemplares fueron trasladados al Centro de Investigación Piscícola de la Universidad de Córdoba (CINPIC) para su posterior análisis.
A cada ejemplar se le realizó una incisión longitudinal que inició en el ano y concluyó debajo de la mandíbula inferior, se procedió a levantar la pared lateral izquierda y luego la pared lateral derecha de la cavidad abdominal dejando al descubierto los órganos internos (esófago, estómago, ciegos pilóricos e intestino) para analizar su situación topográfica. Posteriormente fueron extraídos para medirlos con la ayuda de un analizador de imágenes (Carl Zeiss AxioVisión 4.3, Germany). Con el valor de la longitud intestinal se calculó el coeficiente intestinal (CI), mediante la fórmula CI = Longitud intestinal⁄ Longitud estándar (Bértin, 1958).
DESCRIPCIÓN ESPACIAL Y TOPOGRÁFICA DE LOS ÓRGANOS
La ubicación topográfica de los órganos del tubo digestivo en la cavidad peritoneal se realizó considerando los planos anatómicos de delimitación craneal, media y caudal y para cada uno de estos segmentos las orientaciones dorsal, ventral, lado izquierdo y derecho. Los ciegos pilóricos se describieron con base en sus formas (monotubulares o ramificados); la longitud total de los ciegos pilóricos ramificados se midió desde la base del pedúnculo común de cada ramificación hasta el ápice. El diámetro de los ciegos ramificados se midió en la base y se dividió entre el número de ramificaciones; también, se midió el diámetro en la parte media y en el ápice del ciego, con estos valores se calculó el diámetro promedio de los ciegos (Seixas Filho, 2000).
DESCRIPCIÓN MORFOLÓGICA DE LOS ÓRGANOS
La descripción de la morfología macroscópica de los órganos se realizó con ayuda de un estereoscopio (Carl Zeiss Stemi 2000-C, Germany) con sistema de luz incidente de fibra óptica KL 1500 (Carl Zeiss, Germany) y se tomaron fotografías con cámara digital incorporada (Canon Power Shot G5, Japan).
Para la descripción microscópica se utilizaron técnicas histológicas de rutina, fijación, deshidratación en alcohol, aclaración en xilol, inclusión en parafina y cortes de 3 a 5 µm con la ayuda de un micrótomo manual y finalmente tinción con hematoxilinaeosina (H-E). Las placas se observaron al microscopio de luz (Carl Zeiss Axioestar, Germany) con cámara fotográfica digital incorporada (Canon Power Shot G5, Japon). Para describir los tipos de tejidos y sus células se siguió la terminología propuesta por Junqueira y Carneiro, 1974, Leeson et al., 1990, Takashima e Hibiya, 1995 y Fawcett, 1995. Para describir la anatomía funcional y morfométrica del tubo digestivo se utilizó la terminología propuesta por Seixas Filho, 2000, Sánchez et al., 2003 y Rodríguez et al., 2004.
RESULTADOS
DESCRIPCIÓN ESPACIAL Y TOPOGRÁFICA
La cavidad abdominal de Rubio es alargada y amplia, ovalada en la sección craneal y comprimida en sentido caudal, la disposición de los órganos que conforman el tubo digestivo está directamente relacionada con la forma de la cavidad abdominal y la del cuerpo. La mayoría de los órganos se encuentran en el primer tercio o porción craneal como esófago, primera porción del estómago, la primera asa intestinal y los ciegos pilóricos. En vista lateral izquierda se observan los 2/3 finales del estómago (región fúndica), las regiones cárdica y pilórica del estómago están cubiertas por ciegos pilóricos y parte de hígado; también, se observa la segunda y tercera asa intestinal y la trayectoria del intestino posterior hacia el ano (Fig. 1A). En el lado derecho se observa el estómago cubierto por parte de hígado, el cual llega hasta el final de la porción media de la cavidad abdominal y 2/3 de la totalidad de los ciegos pilóricos entre la porción craneal y media. La mayoría de los ciegos están en el lado derecho; también se observa la región fúndica del estómago y la segunda asa intestinal en la región caudal (Fig. 1B).
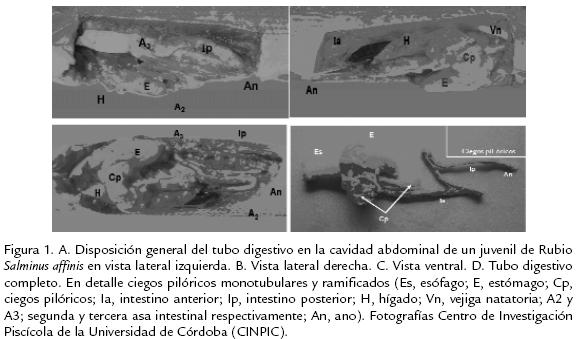
El Rubio presenta tres asas o vueltas intestinales; la primera se encuentra ubicada inmediatamente después del esfínter pilórico (duodeno) justo en los ciegos pilóricos, del lado derecho de la cavidad abdominal; la segunda asa está ubicada en el lado derecho de la región caudal, flexionándose en U, en sentido craneal diagonal hacia el lado izquierdo donde se forma la tercera asa, la cual define la trayectoria final del intestino posterior hacia el ano. Esta última porción del intestino corre paralela a la pared lateral izquierda de la cavidad abdominal y se dispone en sentido latero caudal dirigiéndose hacia el centro hasta llegar al orificio anal (Fig. 1C; Fig. 1D).
DESCRIPCIÓN MICRO Y MACROSCÓPICA DEL TUBO DIGESTIVO
A nivel histológico, en general, el tubo digestivo está conformado por cuatro capas: mucosa, submucosa, muscular y serosa. La capa mucosa está constituida por el epitelio de la mucosa, el cual reviste la parte interna del tubo digestivo y está en contacto con la luz del órgano; en ella se ve la forma y la ubicación del núcleo de la célula, lo cual permite la identificación del epitelio de revestimiento. La lámina propia, conformada por tejido conjuntivo de sostén, forma pliegues longitudinales hacia la luz del órgano. La subcapa muscular, la más externa de la mucosa, presenta dos tipos de epitelios: el de revestimiento y el glandular. El epitelio de revestimiento es la capa de células más superficial, se encuentra recubriendo la parte interna del tubo digestivo, está en contacto con la luz del órgano y con el alimento. La capa submucosa conecta la mucosa con la capa muscular, está ubicada inmediatamente debajo de la capa mucosa y está formada por tejido conectivo laxo; en esta capa el estrato compacto no se diferencia del granuloso. La capa muscular está formada por músculo liso con orientación circular y, seguidamente se encuentra la capa serosa que conforma la superficie externa del tubo digestivo.
Esófago. Es un órgano tubular corto, de pared gruesa que inicia en la parte final de la cavidad bucofaríngea, sin una delimitación externa visible en la superficie del órgano que se asemeje a un esfínter. Tiene una longitud de 10,3±1,8 mm que representa el 7,8% de la longitud total de tubo digestivo y un diámetro 5,8±0,8 mm que equivale a 46,7% del diámetro promedio máximo del estómago; lo cual permite ver una diferenciación entre estos dos órganos. El esófago sirve como conductor del alimento de la boca hacia el estómago, presenta una luz amplia, pliegues internos distensibles y secreciones mucosas que permiten el paso de alimento de gran tamaño con facilidad hacia el estómago. Histológicamente el esófago presenta las cuatro capas básicas para todo el tubo digestivo (mucosa, submucosa, muscular y serosa). Se encuentran pliegues longitudinales que se proyectan hacia la luz del órgano (crestas) muy cortos y de gran diámetro con bordes redondeados, separados unos de otros por surcos poco profundos.
La capa muscular presenta una subcapa de músculo circular que le permitiría al animal reducir la luz del órgano cuando el estómago está lleno. El epitelio de revestimiento que protege el esófago es de tipo estratificado plano no queratinizado con abundantes células caliciformes que interrumpen la regularidad de las células epiteliales tanto en el ápice como en los lados de los pliegues, las secreciones mucosas de las células caliciformes permiten el paso del alimento con mayor facilidad (Fig. 2).

Estómago. Es una bolsa asimétrica en forma de "Y", grande y musculoso, amplio en la porción anterior y estrecho hacia el fondo del órgano. El estómago de Rubio es de fácil identificación con relación a los demás órganos del tubo digestivo. La longitud es de 31,0±6,7 mm representando el 23,5% de la longitud total del tubo digestivo. Se reconocen tres regiones: cárdica, fúndica y pilórica; la cárdica inicia en la unión entre el esófago y el estómago, llamada así por su cercanía con el corazón, se encuentra en el primer tercio del estómago (en sentido cefálico-caudal), presenta gran capacidad de distensión cuando el animal se encuentra en estado de repleción (Fig. 3A); el diámetro promedio de esta región es de 8,4 mm. La región fúndica o región del fondo, tiene forma de saco ciego con diámetro promedio de 6,3 mm; la cual es menor que la cárdica pero mayor que el diámetro del intestino; esta región presenta una notable disminución en el tamaño de las circunvoluciones internas, las cuales se extienden a lo largo del estómago como canales formados por la capa mucosa. La región pilórica es la parte más próxima al intestino con una disminución notable de su diámetro (4,1 mm) pero no del tamaño de las circunvoluciones internas; esta región cumple la misma función de digestión del alimento. Al final de esta región en sentido caudal se encuentra un esfínter pilórico el cual regula el paso del alimento hacia el intestino. Entre estas tres regiones se forma el cuerpo del estómago con pliegues longitudinales y en forma de "V", estos últimos formados por los pliegues de las regiones cárdica y pilórica (Fig. 3B).
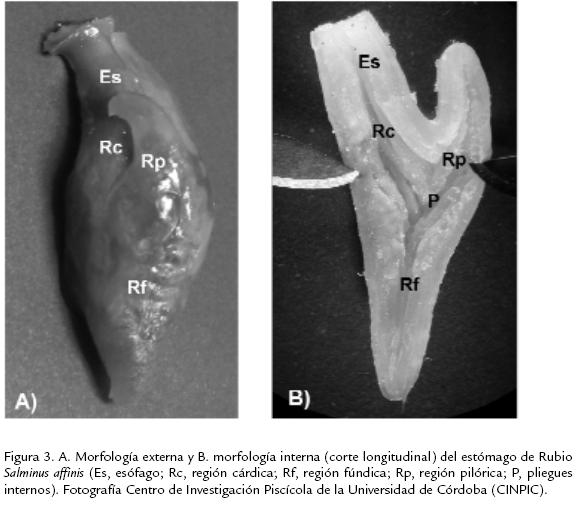
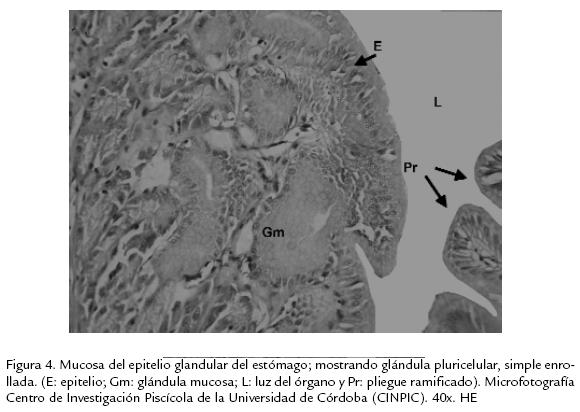
En el estómago el epitelio que recubre la mucosa es de tipo simple cilíndrico con células mucosecretoras, el producto de secreción de estas células se caracteriza por ser acidófilo diferente al de las células caliciformes (Ludovico et al., 1998). En la región fúndica los pliegues longitudinales suelen ser más cortos y, en el lado donde se forma la "Y" entre las regiones cárdica y pilórica más largos. Presenta el mismo tipo de epitelio de revestimiento que la región cárdica, simple cilíndrico con células mucosecretoras con núcleo basal, característica de estas células (Fig. 4). En la región pilórica la luz del órgano se reduce notablemente con relación a la luz que presenta el estómago en la región cárdica; el epitelio que reviste ésta región es el mismo del resto del estómago.
Ciegos pilóricos. Son órganos en forma de sacos ciegos tubulares, generalmente de mayor diámetro en la base, aguzados y redondeados hacia el ápice, evaginados de la porción del intestino inmediatamente después de la región pilórica del estómago, que se conoce como duodeno; una dilatación de la porción inicial del intestino anterior (primera asa) con aberturas que comunican al intestino con los ciegos pilóricos y cuya función es la absorción de nutrientes. El número de ciegos pilóricos puede variar entre 13 y 23, siendo monotubulares y ramificados; estos últimos pueden ser bifurcados, trifurcados y tetrafurcados (Fig. 1D, en detalle) iniciando estas divisiones en la base del pedúnculo común. La longitud promedio de los ciegos monotubulares es de 9,8±3,7 mm y su diámetro promedio es de 1,2±0,2 mm. Los ciegos ramificados presentaron longitud y diámetro menores que los monotubulares; siendo estos valores de 6,5±0,9 mm y 1,1±0,2 mm de longitud y diámetro, respectivamente. El tamaño de los ciegos no tiene relación con la trayectoria del intestino hacia la parte caudal, es decir, se pueden encontrar ciegos largos o cortos a lo largo de la ubicación de estos en el duodeno. La suma de las longitudes promedio de los ciegos monotubulares y ramificados es de 150 mm, representando un incremento del 19% sobre la longitud del intestino. En los ciegos se encuentran pliegues longitudinales internos con distancias muy reducidas entre cada pliegue.
Los ciegos presentan un epitelio de revestimiento simple cilíndrico con escasas células caliciformes, las capas que forman los ciegos son de un diámetro reducido con relación a las capas de los demás órganos debido al tamaño menor de los ciegos.
Intestino. Es un tubo corto que inicia en el esfínter pilórico del estómago y termina en el ano; con una longitud 121,5 mm, representando el 92,3% sobre la longitud total del tubo digestivo y un diámetro promedio de 2,2 mm, presenta tres curvas que se conocen como asas intestinales. Estas asas tienen un diámetro promedio de 3,0 mm, representando un incremento del 27,3% sobre el diámetro intestinal. En el intestino no hay ninguna manifestación externa que permita observar alguna diferenciación morfológica entre la región anterior y la posterior; denominándose intestino posterior a la última porción inmediatamente anterior al ano; el cual concluye abriéndose antes de la aleta anal. El coeficiente intestinal (CI) promedio se estimó en 0,58.
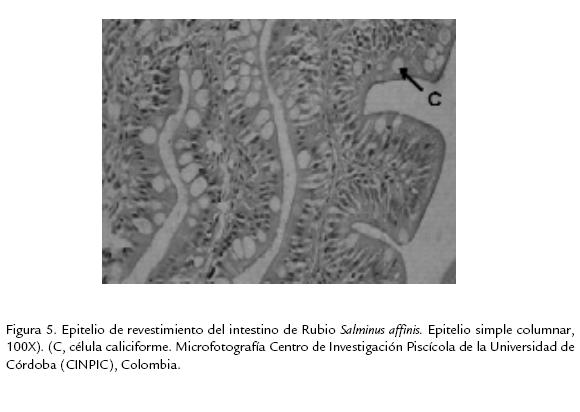
Histológicamente el intestino es un tubo con numerosos pliegues oblicuos que retardan el paso del alimento y aumentan el área de absorción de nutrientes. El epitelio que recubre la capa mucosa es simple cilíndrico con abundantes células caliciformes (Fig. 5). Hacia la parte posterior hay un incremento moderado en la cantidad de células caliciformes permitiendo diferenciar las dos porciones del intestino, la anterior de la posterior.
DISCUSIÓN
En general la estructura del tubo digestivo de los peces es similar a la de la mayoría de los vertebrados superiores. Sin embargo, morfológica y fisiológicamente presentan sus particularidades; ya que viven en un medio acuático que determina ciertas especializaciones que faltan en los animales terrestres (Roberts, 1989). La disposición de los órganos dentro de la cavidad abdominal está directamente relacionada con la forma de la misma y a su vez con la forma del pez, disposición que es generalmente encontrada en la mayoría de los teleósteos (Menin, 1988; Menin y Mimura, 1993; Logato, 1995; Gomide, 1996). La disposición del tubo digestivo de Rubio coincide con lo descrito para carní-voros neotropicales como Pseudoplatystoma coruscans (Seixas-Filho et al., 2001).
Se han realizado numerosos estudios sobre las características histológicas de muchas especies de peces (Yasutaque y Wales, 1983); para la cual se han empleando diferentes técnicas para el análisis del tubo digestivo a través de microscopía de luz (Rodríguez et al., 2004) también se ha empleado la microscopía electrónica de barrido (Sis et al., 1979; Ezeasor y Stokoe, 1980; Caceci, 1984; Humbert et al., 1984; Grau et al., 1992; Jenkins et al., 1992; Nachi et al., 1998). Histológicamente las cuatro capas que conforman el tubo digestivo de Rubio fueron descritas para otros peces por Takashima e Hibiya, 1995, sin embargo en Girardinus metallicus se observaron tres capas: mucosa, submucosa y muscular (Rodríguez et al., 2004). Estos estudios demuestran la conformación variable del tubo digestivo de los peces para atender las particularidades del régimen alimentario de cada especie. Takashima e Hibiya, 1995, reportan que la estructura del tubo digestivo cambia considerablemente a lo largo del mismo y que grandes diferencias son encontradas entre especies.
El esófago de Rubio, en general corresponde con la estructura descrita en Bagre de canal Ictalurus punctatus (Grizzle y Rogers, 1976), en Salmónidos (Yasutake y Wales, 1983), Sábalo (Domitrovic, 1983; Domitrovic y Moreira, 1985) y a la descrita por Menin, 1988, para seis teleósteos de diferentes hábitos alimenticios. Presenta una descripción similar a las realizadas en Characidos iliófagos como Semaprochilodus insignis (Chávez y Vazzoler, 1984) y para detritívoros como Plecostomus sp. (Zamith, 1952). Por otra parte, Rodríguez et al., 2004, describieron en el esófago del carnívoro Gambusia punticulata un epitelio pseudoestratificado plano con células caliciformes y continúa con epitelio pseudoestratificado cilíndrico con células caliciformes hacia el estómago. Para Girardinus metallicus la mucosa que protege el esófago está constituida por epitelio pseudoestratificado cilíndrico con células caliciformes.
El estómago de Rubio es un órgano grande a diferencia de los peces omnívoros y herbívoros en los cuales el estómago es de baja capacidad volumétrica y por tanto deben realizar varias ingestiones en el día (Vásquez-Torres, 2004). La estructura general no difiere mucho de la observada en otros teleósteos (Grizzle y Rogers, 1976; Yasutake y Wales, 1983; Domitrovic, 1983; Domitrovic y Moreira, 1985; Roberts, 1989). Está constituido por cuatro capas: mucosa, submucosa, muscular y serosa al igual que en Cachama blanca Piaractus brachypomus (Vásquez-Torres, 2004), mientras que para Prochilodus marggravii y Leporinus reinhardtii el estómago presenta tres capas: mucosa, muscular y serosa, con epitelio de revestimiento simple cilíndrico mucosecretor (Castro, 2002). Entre las regiones cárdica, pilórica y del fondo está formado el cuerpo del estómago y es hacia la porción central del mismo donde se encuentra la mayor cantidad de glándulas gástricas las cuales estarían presentes en otros peces hacia la región cárdica (Rotta, 2003).
La presencia de ciegos pilóricos en peces no está relacionada con el hábito alimentario y su número no es constante para una misma especie. En Rubio se encontraron entre 13 y 23; mientras que en Pseudoplatystoma coruscans especie con el mismo hábito alimentario hay ausencia total de estos apéndices (Seixas Filho et al., 2001); en Piracanjuba (omnívoro) oscilan entre 42 y 93 (Seixas Filho et al., 2000).
El intestino de Rubio es un tubo corto si se compara con omnívoros como Piracanjuba Brycon orbygnianus que presenta nueve asas intestinales (Seixas Filho et al., 2000). La mucosa del intestino y de los ciegos pilóricos presenta pliegues internos oblicuos, siendo estos los más complejos de todo el tubo digestivo ya que ayudan a retardar el paso del alimento y aumentan el tiempo de absorción de los nutrientes ayudados por las asas intestinales las cuales aseguran el transporte lento del material en procesamiento. Esto está directamente relacionado con los procesos fisiológicos de digestión y absorción de nutrientes; mientras que el epitelio de la mucosa del esófago y el estómago presentan pliegues menos complejos (Angelescu y Gneri, 1949). En el Rubio no hay delimitación morfológica externa que permita diferenciar el intestino anterior del posterior, la misma situación fue descrita por Seixas Filho, 2000, en Piracanjuba. Sin embargo, a nivel histológico se logra ver un pequeño incremento en la cantidad de glándulas unicelulares (células caliciformes) en el intestino posterior con relación al anterior, situación que debe ser estudiada más a fondo utilizando coloraciones diferenciales para células caliciformes (Castro, 2002). El epitelio de revestimiento del intestino de Rubio es de tipo simple cilíndrico con células caliciformes, similar a lo encontrado en Salminus brasiliensis, Prochilodus marggravii y Leporinus reinhardtii (Castro, 2002), lo contrario ocurre en el intestino de Gambusia punticulata y Girardinus metallicus en el que el epitelio es de tipo pseudoestratificado cilíndrico (Rodríguez et al., 2004).
Todo el tubo digestivo de Rubio está formado por cuatro capas en cada uno de los órganos que lo conforman, en algunos peces como Salminus brasiliensis, Prochilodus marggravii y Leporinus reinhardtii a nivel de estómago se encuentran tres capas: mucosa, muscular y serosa mientras que para el intestino se observan cuatro capas: mucosa, submucosa, muscular y serosa (Castro, 2002).
Atendiendo los valores del coeficiente intestinal (CI) propuesto por Bértin, 1958, el del Rubio (0,58) corresponde a un carnívoro; en otro carnívoro Pseudoplatystoma coruscans este coeficiente fue reportado en 0,48 (Seixas Filho et al., 2001). Como es de esperarse en omnívoros como Brycon orbignyanus este coeficiente fue estimado en 1,17 (Seixas Filho et al., 2000).
CONCLUSIONES
Los resultados obtenidos en este estudio permiten concluir que Rubio Salminus affinis presenta una morfología macro y microscópica del tubo digestivo similar a la de los teleósteos con régimen alimenticio carnívoro. Por esta razón el manejo alimentario que tendría que darse a la especie en cautiverio debe considerar el suministro de niveles altos de proteína, probablemente de origen animal, situación que disminuiría la posibilidad de ofrecer la especie como una opción sostenible para la piscicultura.
AGRADECIMIENTOS
Los autores agradecen al profesor Juan Carlos Carrascal de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad de Córdoba, Colombia su colaboración en la interpretación de los cortes histológicos. Este estudio fue financiado en su totalidad por la Oficina de Investigación y Extensión de la Universidad de Córdoba (proyecto FMZ-02-04, 1120143).
BIBLIOGRAFÍA
ANGELESCU V, GNERI FS. Adaptaciones del aparato digestivo al régimen alimenticio en algunos peces del río Uruguay y del río de La Plata. Rev Inst Invest Mus Argent Cienc Nat. 1949;1:161-272.
[ Links ]ATENCIO GARCÍA V, GENES LÓPEZ F, MADARIAGA MENDOZA D, PARDO CARRASCO S. Hematología y química sanguínea de juveniles de rubio (Salminus affinis Pisces: Characidae) del río Sinú. Acta biol Colom. 2007;12S:27-40.
[ Links ]BÉRTIN L. Appareil digestif. En: Grassé PP (Ed). Traité de zoologie. Paris: Manson; 1958;13:1248-301.
[ Links ]BRAUNER CJ, VAL AL. Oxygen transfer. En: Val AL, Almeida-Val VMF, Randall D. (Ed). The physiology of tropical fishes. FISH PHYSIOLOGY: VOLUME 21 Academic Press; 2006. CACECI T. Scanning electron microscopy of goldfish, Carassius auratus, intestinal mucosa. J Fish Biol. 1984;25:1-12.
[ Links ] [ Links ]CASTRO E. Estudos histológico, histoquímico e histoquantitativo de células endócrinas do estômago e intestino médio de peixes (teleostei) de água doce, com diferentes hábitos alimentares [Dissertação de Mestrado]. Viçosa (Bras): Universidade Federal de Viçosa; 2002.
[ Links ]CHAVEZ PT, VAZZOLER G. Aspectos biológicos dos peixes amazônicos. III. Anatomia microscópica do esôfago, estômago e cecos pilóricos de Semaprochilodus insignis (Characiformes: Prochilodontidae). Acta Amazonica. 1984;14:343-53.
[ Links ]DAHL G. Los peces del norte de Colombia. Bogotá (Col): Ministerio de Agricultura, INDERENA; 1971.
[ Links ]DOMITROVIC HA. Histología del tracto digestivo del Sábalo (Prochilodus platenses, Holmberg 1880, pisces Prochilodontidae). Rev Ictiología. 1983;1(1):35-44.
[ Links ]DOMITROVIC HA, MOREIRA JE. Microscopia eletrônica de varredura do trato digestivo do Sábalo (Prochilodus platensis, Holmberg 1880, Pisces Prochilodontidae). Rev Bras Cien Morfol. 1985;1(1):21-30.
[ Links ]ESLAVA-MOCHA PR, CRUZ CASALLAS PR, VÁSQUEZ TORRES W. Morfología del aparato digestivo de la Cachama blanca Piaractus brachypomus. Rev MVZ Unillanos. 2000;2(2):22-31.
[ Links ]ESLAVA-MOCHA PR, SUÁREZ MAHECHA H, PARDO CARRASCO SC, ARIAS CASTELLANOS JA, CRUZ CASALLAS PE. Morfología macro y microscópica del esófago, estómago y ciegos pilóricos del yamú Brycon siebenthalae. Rev Orinoquia. 2001;5(1):111-28.
[ Links ]EZEASOR DN, STOKOE WM. Scanning electron microscopic study of the gut mucosa of the rainbow trout Salmo gairdneri Richardson. J. Fish Biol. 1980;7:529-539.
[ Links ]FAWCETT DW. Tratado de histología. 12° ed. Madrid (Esp): Interamericana-Mc Graw-Hill 1995.
[ Links ]GOMIDE AT. Anatomia funcional e morfometría comparativas do tubo digestorio de Trairão (Hoplias cf. Lacerdae Ribeiro, 1908) (Characiformes, Erithrinidae), em diferentes classes de tamanho [Tesis doctoral] Viçosa (Bra); Universidade Federal de Viçosa; 1996.
[ Links ]GRAU A, CRESPO S, SARASQUETE MC, GONZALEZ ML. The digestive tract or the amberjack, Seriola dumerili, Risso: a light and scanning electron microscope study. Rev. J. Fish Biol. 1992;41:287-303.
[ Links ]GRIZZLE JM, ROGERS WA. Anatomy and histology of the channel Catfish. Auburn Pinting Inc. Auburn University, Agr. Exptl. Station; 1976. HUMBERT W, KIRSCH R, MEISTER MF. Scanning electron microscopic study of the oesophageal mucous layer in the eel, Anguilla anguilla L. J Fish Biol. 1984;25:117-22.
JENKINS PG, PULSFORD AL, HARRIS JE. Microscopy of the absortive cells and gut associated lymphoid tissue of the flounder Platichthys flesus. J Mar Biol. 1992;72:553-67. [ Links ] [ Links ] [ Links ]JUNQUEIRA LC, CARNEIRO J. Histología básica. Barcelona (Esp): Salvat Editores; 1974.
[ Links ]LEESON TS, LEESON CR, PAPARO AA. Texto/Atlas de histología. México: Interamericana Mc Graw-Hill; 1990.
[ Links ]LIMA CT, MALABARBA LR, BUCKUP PA, DA SILVA JF, VARI RP, HAROLD A. Genera incertae sedis in Characide. En: Reis RE, Kullander SO, Ferraris CJ. Checklist of the freswater fishes of south and Central America. Porto Alegre (Bra): Edipucrs; 2003:106-69.
[ Links ]LOGATO PV. Anatomo-histología funcional do aparelho digestório do Pacú, Piaractus mesopotamicus Holmberg, 1988 (Characiformes, Characidae, Myelinae) [Tesis doctoral] Viçosa; Universidade Federal de Viçosa; 1995.
[ Links ]LUDOVICO L, RODRIGUES C, LESQUEVES DE CASTRO R. Histologia comparada. Revisada e ampliada. São Paulo (Bra); 1998.
[ Links ]MARTÍNEZ A. Peces deportivos de Colombia. Bogotá: Fondo Cultural Cafetero; 1981.
[ Links ]MEJÍA KE, NAVARRO YP. Determinación de parásitos en Rubio (Salminus affinis Steindachner, 1880) de las cuencas de los ríos Sinú y San jorge. [Trabajo de pregrado] Montería: Universidad de Córdoba; 2006.
[ Links ]MENIN E. Anátomo-histologia funcional comparativa do aparelho digestivo de seis Teleostei (Pisces) de água doce. [Tese Doutorado] São Paulo: Instituto de Biociências Universidade de São Paulo; 1988.
[ Links ]MENIN E, MIMURA OM. Anatomia comparativa do esôfago de seis peixes Teleostei de distintos hábitos alimentares. Rev Ceres. 1993;40:334-69
[ Links ]MILES C. Los peces del río Magdalena. 2 ed. Bogotá (Col): Ministerio de Economía Nacional, Pesca y Caza; 1971.
[ Links ]MOJICA JI, CASTELLANOS C, USMAS J, ÁLVAREZ R. Libro rojo de peces dulce acuícolas de Colombia. La serie libros rojos de especies amenazadas de Colombia. Bogotá (Col): Instituto de Ciencias Naturales, Universidad Nacional de Colombia. 2002:547-566.
[ Links ]MOJICA JI. Lista preliminar de las especies de peces dulceacuícolas de Colombia. Rev Acad Col Cienc. 1999;23(S):547-66.
[ Links ]NACHI AM, HERNANDEZ-BLAZQUEZ FJ, BARBIER RL, LEITE RG, FERR S, PHAN MT. Intestinal histology of a detritivorous (iliophagous) fish Prochilodus scrofa (Characiformes, Prochilodontidae) Annales des Sciences Naturelles -Zoologie et Biologie Animale. 1998;19(2):81-88
[ Links ]PARDO S, MEJÍA K, NAVARRO Y, ATENCIO V. Prevalencia y abundancia de Contracaecum sp. en Rubio Salminus affinis en el río Sinú y San Jorge: Descripción morfológica. Rev MVZ Córdoba. 2007;12(1):887-96.
[ Links ]ROBERTS RJ. Fish pathology. London: Bailliene Tindall; 1989.
[ Links ]RODRÍGUEZ J, GONZÁLEZ E, HERNÁNDEZ N, CAPÓ V, GARCÍA I. Comparación morfológica e histológica del tubo digestivo de Gambusia punticulata y Girardinus metallicus, peces utilizados en el control biológico de mosquitos. Rev. Cubana Med Trop. Instituto de Medicina Tropical Pedro Kourí. 2004;56(1):73-76.
[ Links ]ROTTA MA. Aspectos gerais da fisiologia e estrutura do sistema digestivo dos peixes relacionados à piscicultura. Corumbá, MS (Bra). Empresa Brasileira de Pesquisa Agropecuaria; 2003.
[ Links ]SÁNCHEZ RM, GALVIS G, VICTORIANO P. Relación entre características del tracto digestivo y los hábitos alimentarios de los peces del río Yucao, sistema del río Meta (Col); Gayana. 2003;65(1):75-86.
[ Links ]SEIXAS FILHO JT, BRAS JM, GOMIDE AT, OLIVEIRA MG, DONZELE JL, MENIN E. Anatomia funcional e morfometría dos intestinos e dos cecos pilóricos do teleostei (Pisces) de água doce Brycon orbignyanus (Valenciennes,1849). Rev Bras Zootec. 2000;29(2):313-324.
[ Links ]SEIXAS FILHO JT, BRAS JDM, GOMIDE ATM, OLIVERA MGA, DONZELE JL, MENIN E. Anatomia funcional e morfometría do intestino no Teleostei (Pisces) de água doce Surubim (Pseudoplatystoma coruscans Agassiz, 1829). Rev Bras Zootec. 2001;30(6):1670-80.
[ Links ]SIS RF, IVES PJ, JONES DM, LEWIS DH, HAENSLEY WE. The microscopic anatomy of the oesophagus, stomach and intestine of the channel catfish Ictalurus punctatus. J Fish Biol. 1979;4:179-86.
[ Links ]TAKASHIMA F, HIBIYA T. An atlas of fish histology normal and pathological features. 2 ed. Tokio (Jap): Kondansha; 1995.
[ Links ]VÁSQUEZ-TORRES W. Principios de nutrición aplicada al cultivo de peces. Villavicencio: Universidad de los Llanos; 2004.
[ Links ]YASUTAKE WT, WALES JH. Microscopic anatomy of salmonids: an atlas. U.S. Fish Wildlife Serv. Res. Publ. 1983.
[ Links ]ZAMITH AP. Contribuição para o conhecimento da estrutura da mucosa do esôfago dos vertebrados. Ann Esc Sup Agric Luis de Queirós. 1952;9:359-434.
[ Links ]
