










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.14 no.1 Bogotá Jan./Apr. 2009
ABEJAS VISITANTES DE Mimosa pigra L. (MIMOSACEAE):COMPORTAMIENTO DE PECOREO Y CARGAS POLÍNICAS
Bees visiting Mimosa pigra L. (Mimosaceae):foraging behavior and pollen loads
CLARA ISABEL AGUILAR SIERRA1, ALLAN HENRY SMITH PARDO2, Ph.D.
1Profesora Asociada, Facultad de Ciencias, Universidad Nacional de Colombia, AA 3840, Medellín. ciaguila@unalmed.edu.co
2Posgrado en Entomología, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín. ahsmithp@unal.edu.co
Presentado 2 de mayo de 2008, aceptado 11 de junio de 2008, correcciones 19 de octubre de 2008.
RESUMEN
Se estudiaron las cargas polínicas de 13 taxones de abejas capturadas visitando a Mimosa pigra en la zona de influencia del Embalse Porce II (Antioquia, Colombia). De los 21 tipos polínicos encontrados, M. pigra presenta el mayor porcentaje acumulado de colecta; Mimosa pudica, Piper aduncum, Solanum diversifolium, Warszewiczia coccinea y Psidium guajava, en su orden, se pueden considerar fuentes alternativas de polen, para varias de las especies de abejas cuyas cargas polínicas fueron analizadas. Dentro de las especies de abejas capturadas visitando a M. pigra se pueden diferenciar varios grupos según el tipo y abundancia relativa de los tipos polínicos encontrados en sus cargas. Uno de ellos, incluye a siete especies de abejas con más del 85% de granos de polen de M. pigra; otro, con cuatro especies de abejas que colectaron más del 94,5% del polen en M. pigra y M. pudica. Adicionalmente, se encontraron especies como Trigona dorsalis con cargas de M. pigra (59,4%), de S. diversifolium (37,8%) y especies de abejas como Lasioglossum sp. 113, en cuyas cargas polínicas predominan los granos de polen de P. aduncum (61,8%) y de W.coccinea (36,4%), en contraste con los de M. pigra (1,3%). En cuanto a la riqueza de tipos polínicos colectados por las abejas sobresalen Trigona muzoensis (12 tipos polínicos) y T. dorsalis (10 tipos), lo cual refleja nichos tróficos más amplios para estas especies y deja duda sobre su constancia floral o hábitos de limpieza.
Palabras clave: Mimosa pigra, cargas polínicas, tipos polínicos, abejas, polinización.
ABSTRACT
We studied the pollen loads of 13 taxa of wild bees visiting the flowers of Mimosa pigra on the influence zone of the Dam Project Porce II (Antioquia, Colombia). Out of 21 different pollen types, M. pigra represents the higuest percentage; Mimosa pudica, Piper aduncum, Solanum diversifolium, Warszewiczia coccinea and Psidium guajava, in that order, were also abundant, and are alternative sources of pollen for the different kinds of bees recorded. Among the species of bees collected, we differentiate several groups: one of them, includes seven species of bees whose pollen loads are in a high porcentaje of M. pigra (86% of pollen grains or above); another group of bees with high percentage (more than 94%) of pollen grains of Mimosa species (M. pigra and M. pudica); and the other two groups that include the bee species: Trigona dorsalis (who collected more than half of their pollen resources from M. pigra (59.4%) in addition to a high percentage of S. diversifolium (37.8%); the last kind of foragers includes Lasioglossum sp.113 who collected mostly pollen from species different to Mimosa (including P. aduncum with 61.8% of pollen grains, and W. coccinea with 36,4%) and only a small percentage of M. pigra (1,3%). In terms of the species of bees that collected a highest diversity of pollen grains, two species: Trigona muzoensis (12 pollen types) and T. dorsalis (10 pollen types) outstand other species of bees on on feeding niche and question either the bees capacity of cleaning or their floral constancy.
Key words: Mimosa pigra, Pollen loads, Pollen types, Bees, Pollination.
INTRODUCCIÓN
El estudio palinológico de las cargas polínicas colectadas por las abejas en las plantas con flores, permite un mejor conocimiento de la relación abeja-planta; así mismo, sirve para determinar la flora que sostiene a sus comunidades en una región particular. A pesar de la utilidad del análisis palinológico, es necesario efectuar observaciones complementarias en el campo sobre el comportamiento de las abejas y la oferta de recursos vegetales (Nates et al., 2002). Las abejas dependen de una buena floración y de condiciones climáticas favorables para sus labores de pecoreo; en épocas de escasez necesitan de reservas que les permitan mantener a los adultos y a sus crías (Girón 1995; Smith-Pardo, 1999).
En los ecosistemas tropicales húmedos es muy frecuente encontrar una amplia gama de mutualismos entre los insectos y las flores, desde ladrones de polen o néctar y visitantes accidentales, hasta polinizadores altamente específicos que establecen una simbiosis obligada, en la cual la planta y el insecto que la poliniza son completamente dependientes; por ejemplo, las orquídeas de las subfamilias Stanhopeinae y Catasetinae polinizadas exclusivamente por machos de abejas euglossinas (Williams y Whitten, 1983; Vogel y Westerkamp, 1991; Mills et al., 1993; Endress, 1994). La alteración o completa disrupción de estos mutualismos puede tener efectos en cascada, que afectan a otros organismos aunque no participen directamente de dichas relaciones (Aizen et al., 2002). En muchos casos, los mutualismos de polinización y dispersión de semillas son necesarios para garantizar la supervivencia de las poblaciones y comunidades vegetales. La polinización es el medio por el cual una planta mantiene contacto con otros miembros de la población y corresponde a la etapa en que los genes se pueden mezclar para una nueva generación; esto permite la variabilidad genética que es el punto de partida para los cambios evolutivos.
Los diferentes métodos de polinización y dispersión de semillas tienen un gran impacto sobre la amplitud de la distribución de los genes, es decir, sobre el flujo de genes en las mismas con las comunidades vegetales; igualmente, juegan un papel muy importante en la cohesión de poblaciones y comunidades (Proctor et al., 1996). De esta manera se establecen interacciones tales que, para preservar la diversidad, es necesario conservar a los polinizadores, los cuales a su vez dependen del equilibrio del ecosistema (Smith-Pardo, 1999; Michener, 2000). A pesar de que existen numerosos estudios palinológicos de las cargas polínicas, la miel y las reservas de polen de Apis mellifera (Girón, 1995; Andrada et al., 1998; Terraba et al., 2003; Sabugosa-Madeira et al., 2007), los análisis de diversidad polínica en las cargas de polen de abejas silvestres, son relativamente escasos y dispersos (Jonggjitvimol y Wattanachaiyingcharoen, 2006; Goldblatt et al., 2000).
Jonggjitvimol y Wattanachaiyingcharoen, 2006, estudiaron en Tailandia la composición polínica en las canastas de polen de tres especies de Trigona. Encontraron 29 tipos polínicos pertenecientes a 13 familias; clasificaron a las abejas así: T. collina, con todos los tipos polínicos en sus cargas, la especie más generalista, seguida de T. apicalis con 20 tipos polínicos y T. fimbriata con 16 tipos, la especie menos generalista. Goldblatt et al., 2000, en los Estados Unidos, verificaron que plantas de Sparaxis (Iridaceae: Ixioideae) son polinizadas principalmente por abejas silvestres y melíferas, como lo revela su comportamiento de pecoreo y la especificidad de sus cargas de polen.
Dada la relevancia de las abejas, no solo en los procesos reproductivos de las plantas con flores, sino también en los procesos de regeneración vegetal y, en general, en la conservación del equilibrio ecológico de los hábitat en los cuales se encuentran, se llevó a cabo el análisis palinológico de las cargas polínicas colectadas por abejas visitantes de Mimosa pigra L. (Mimosaceae); una planta pionera, abundante en áreas degradadas cercanas a orillas de ríos y quebradas de la zona de influencia del Embalse Porce II (Antioquia, Colombia). El objetivo de este trabajo fue estudiar el comportamiento de pecoreo y las cargas polínicas de las abejas silvestres visitantes de flores de M. pigra y, de acuerdo con ello, determinar si presentaban patrones de comportamiento y porcentajes de granos de polen en sus cargas, acordes con la estrategia reproductiva, tipo r, de esta planta (oferta permanente de recursos florales). Es decir, establecer si las especies de abejas que visitan a M. pigra, poseen hábitos alimenticios generalistas (cargas polínicas con diferentes tipos polínicos) o si, por el contrario, colectan altos porcentajes de granos de polen de esta especie, dada su constancia floral. Estos resultados pueden contribuir al conocimiento de la dieta de las abejas de la zona de estudio y ser útiles para recomendar el uso de plantas apícolas pioneras, en programas de recuperación de áreas degradadas.
MATERIAL Y MÉTODOS
Esta investigación se realizó en el área de influencia del proyecto hidroeléctrico Porce II, situado al noroeste del departamento de Antioquia, Colombia, a 120 km de la ciudad de Medellín (Smith-Pardo, 1999). Según el sistema de clasificación propuesto por Holdridge, en el área del proyecto se encuentran tres zonas de vida: el bosque húmedo tropical (bh-T), el cual ocupa gran parte del proyecto, principalmente circundando al embalse; la transición del bosque húmedo tropical al bosque muy húmedo premontano (bh-T -bmh-PM); y el bosque muy húmedo premontano (bmh-PM), localizado en las partes más altas de las vertientes del río Porce (Cano, 1995). La temperatura promedio anual es de 22,5 °C, la humedad relativa de 83% mensual y la pluviosidad media anual de 3.050 mm. El régimen de lluvias es bimodal: un período seco desde noviembre hasta marzo y uno lluvioso de abril a octubre con una época seca entre julio y agosto (Smith-Pardo, 1999).
La cobertura vegetal de la zona está compuesta principalmente por pastos que empiezan a ser reemplazados por rastrojos bajos; así mismo, se observan algunas áreas con rastrojos más altos y fragmentos de bosque localizados en la parte superior de colinas y vertientes y en las riberas del río Porce y algunos de sus afluentes (Smith-Pardo, 1999). Se establecieron parcelas de 50 m x 100 m, en fase sucesional de pastizal a rastrojo bajo, en cuatro localidades: Santa Lucía (1.100 msnm, N 06º 46' 34,2'' W 75º 06' 18,5''), Picardía (1.037 msnm, N 06º 54' 01,1'' W 75° 03' 11,1”), Fosforito (945 msnm, 06º 45' 37, 8'' W 75º 06' 28, 1'') y Cancana (941 msnm, N 06º 46' 47,6'' W 75° 05' 36,9”). En cada una de ellas se escogieron 24 individuos de M. pigra. Esta planta, conocida como “Uña de gato”, es muy común en la región, en suelos degradados por la actividad minera, así como en valles aluviales. Sus inflorescencias estaminadas, con flores de color lila pálido, abren temprano en la mañana y ofrecen polen y néctar como recompensa a sus visitantes.
El comportamiento de pecoreo de las abejas se observó desde las 8 h hasta las 16 h, por espacio de quince minutos, en cada una de las plantas de las parcelas estudiadas, durante cuatro días consecutivos. Se registraron las especies de abejas visitantes, el número de individuos, el tiempo total de visita y el número de inflorescencias visitadas y se midieron la temperatura y la humedad relativa, cada dos horas. Estas variables se sometieron a análisis de covarianza factoriales para establecer si existían relaciones estadísticamente significativas entre ellas mediante el programa Statgraphics® v.3.0 para Windows.
De cada una de las especies de abejas visitantes se capturaron entre dos y cuatro especímenes, los cuales se colocaron en viales de vidrio con ácido acético glacial, debidamente rotulados. Los ejemplares colectados se lavaron con el mismo ácido utilizando una jeringa y el material polínico obtenido, se sometió al proceso de Acetólisis de Erdtman, 1960, con las modificaciones descritas por Fonnegra, 1989. Se prepararon cuatro placas permanentes con gelatina glicerinada, selladas con parafina y se incluyeron en la palinoteca del Laboratorio de Ecología y Conservación Ambiental (LECA) del Departamento de Ciencias Forestales, Facultad de Ciencias Agropecuarias de la Universidad Nacional de Colombia, Sede Medellín.
El análisis palinológico comprendió la identificación de los tipos polínicos mediante la comparación con la colección de referencia del LECA y la consulta de literatura especializada. Las preparaciones se observaron bajo un microscopio óptico OLYMPUS CH-30, con un aumento de 400X. Sobre las cuatro placas preparadas se contabilizaron entre 1.000 y 1.500 granos de polen, según las recomendaciones de Vergeron, 1964, para análisis palinológicos en miel. Los resultados se expresaron como porcentajes para estimar la abundancia relativa de cada tipo polínico.
El paquete estadístico MVSP v.3 se utilizó para para obtener un análisis de agrupamiento de los diferentes tipos polínicos encontrados en las cargas de las distintas especies de abejas, de las abejas mismas y de la riqueza de tipos polínicos colectados por ellas.
RESULTADOS
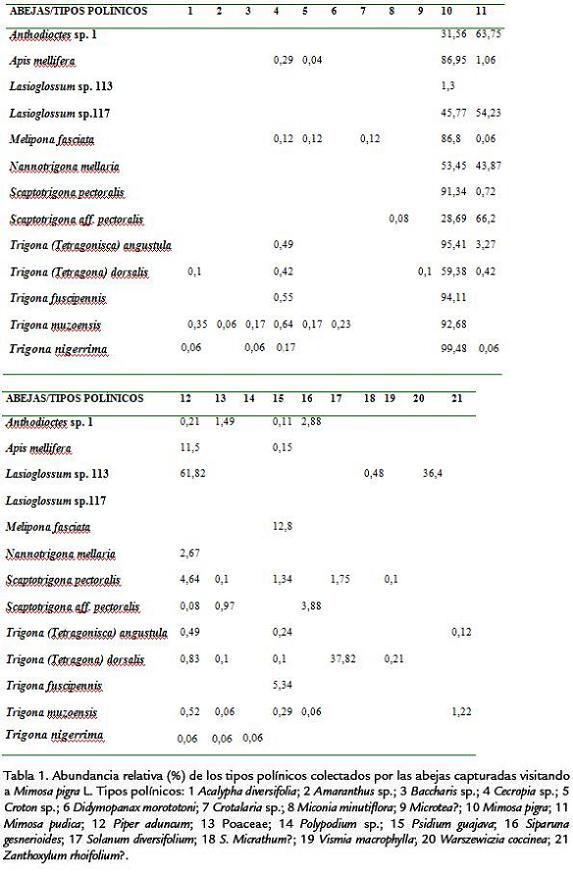
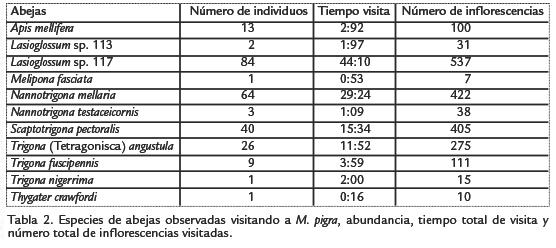
En la Tabla 1, se consigna la abundancia relativa de los tipos polínicos colectados por las especies de abejas capturadas (13) que visitaron las flores de Mimosa pigra L., así como la riqueza de tipos polínicos presentes en sus cargas. Trigona nigerrima Cresson 1878, colectó el 99,5% de sus cargas polínicas en M. pigra, seguida de Trigona fuscipennis Friese 1908 (94,1%) y Trigona muzoensis Schwarz 1948(92,7%); a diferencia de estas especies, Lasioglossum sp.113 sólo colectó el 1,3% de granos de polen de la planta estudiada. Lassioglosum sp.117 se destaca por ser la abeja con los valores más altos para el número total de individuos, el tiempo total de visita y el número total de inflorescencias visitadas; la siguen dos especies de meliponinos, Nannotrigona mellaria Smith 1862 y Scaptotrigona pectoralis Dalla Torre 1896 (Tabla 2).
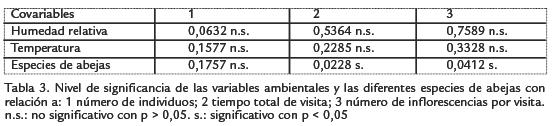
En las Tablas 3, 4 y 5 se registran los resultados de los análisis de covarianza factoriales y de las pruebas de rangos múltiples; es importante señalar que Melipona fasciata Latreille 1811, T. nigerrima y Thygater crawfordi Urban 1967, representadas por un solo individuo (Tabla 2), se excluyeron de estos análisis.
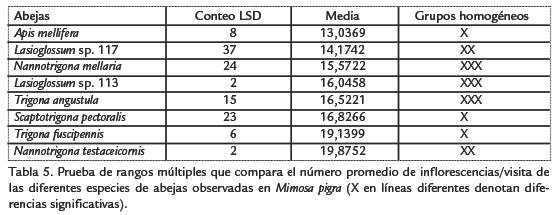
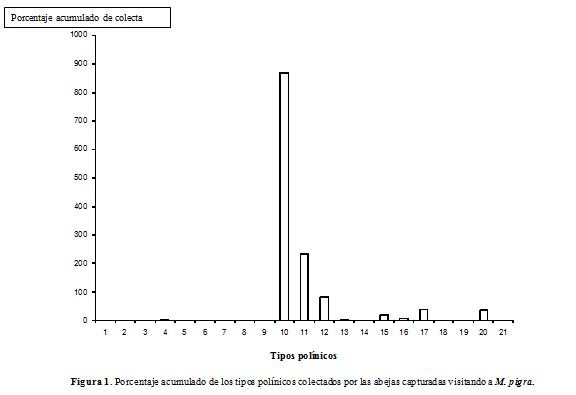
En el histograma del porcentaje acumulado de los tipos polínicos colectados (Fig. 1), M. pigra alcanza el mayor valor (866,9%), seguida de Mimosa pudica L. (233,6%). En el análisis de agrupamiento (Fig. 2), M. pigra forma un grupo separado con relación a los otros 20 tipos polínicos encontrados, como quiera que en la mayoría de las cargas polínicas analizadas su abundancia relativa fue mayor y es la colectada por todas las abejas capturadas visitándola.
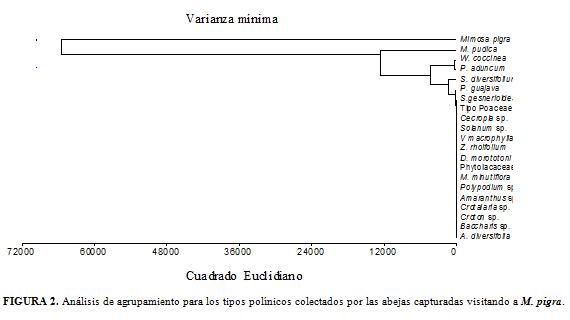
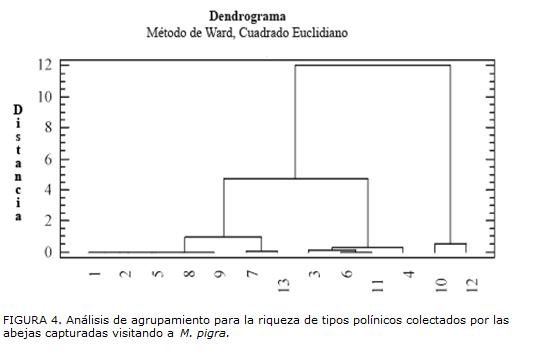
El análisis de agrupamiento para las abejas capturadas en M. pigra (Fig. 3), permite observar que éstas se distribuyen en cuatro grupos, según la abundancia relativa de los tipos polínicos colectados, como se explicará más adelante. En cuanto al análisis de agrupamiento para la riqueza de tipos polínicos presentes en las cargas estudiadas (Fig. 4) se forman tres grupos.
DISCUSIÓN
El mayor porcentaje acumulado de colecta del tipo polínico de M. pigra (Fig. 1), unido a una frecuencia superior a 85%, en el análisis palinológico de las cargas polínicas de siete especies de abejas (Tabla 1), permiten señalar la importancia de esta planta como fuente de polen para las abejas. Esto también se corrobora en el análisis de agrupamiento para los tipos polínicos colectados (Fig. 2), en el cual M. pigra forma un grupo aparte. Sin embargo, M. pudica, Piper aduncum L, Solanum diversifolium Schltdl., Warszewiczia coccinea (Vahl) Klotzsch y Psidium guajava L, son especies vegetales valiosas en la dieta de varias abejas, cuyos nichos tróficos les permiten explotar estas plantas en floración.
En la Tabla 2 se observa que Lasioglossum sp.117 tuvo los mayores valores de individuos visitantes, tiempo acumulado de visita y número total de inflorescencias visitadas, seguida de N. mellaria y S. pectoralis. Sin embargo, de acuerdo con los resultados de la Tabla 3, no hay una relación estadísticamente significativa entre el número total de individuos y las diferentes especies de abejas que fueron observadas visitando a M. pigra (p = 0,1757), pero sí entre la duración total de las visitas (p = 0,0228) y el número total de las inflorescencias visitadas por las distintas especies de abejas (p = 0,0412); en ninguno de los análisis se observa covariación significativa con las variables ambientales, lo cual era de esperarse por cuanto el ambiente es multivariado y en él ocurre sinergismo entre variables.
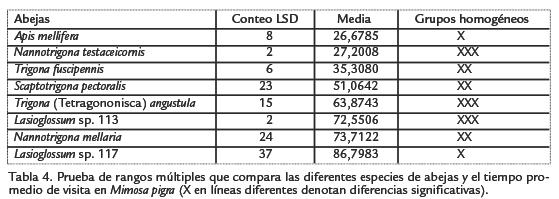
La prueba de rangos múltiples de la Tabla 4 muestra que Lasioglossum sp.117 estuvo, en promedio, visitando durante más tiempo a las inflorescencias de M. pigra (86,7983), pero la duración media de visitas no difirió significativamente de las de N. mellaria, Lasioglossum sp.113, Trigona angustula Illiger 1900 y Nannotrigona testaceicornis Lepeletier 1836. Sí existe diferencia estadísticamente significativa con el tiempo promedio de visita de Apis mellifera L., T. fuscipennis y S. pectoralis.
Aunque Lasioglossum sp.117 visitó el mayor número total de inflorescencias de M. pigra (Tabla 2), los resultados de la prueba de rangos múltiples (Tabla 5) muestran que N. testaceicornis registra el mayor número promedio de inflorescencias/visita (19,8752), pero no presenta diferencias significativas con el número promedio de inflorescencias visitadas por las demás especies de abejas, excepto con A. mellifera, la cual presenta el menor número promedio de inflorescencias/visita.
En la Figura 3 se observa que las abejas visitantes de M. pigra, se distribuyen en cuatro grupos. Un primer grupo está conformado por siete especies que colectaron un alto porcentaje de sus cargas polínicas en M. pigra, como puede constatarse en la Tabla 1: T. nigerrima (99,5%), T. angustula (95,4%), T. fuscipennis (94,1%), T. muzoensis (92,7%), S. pectoralis (91,3%), A. mellifera (87,0%) y M. fasciata (86,8%); esto indica su alta constancia floral ya que colectan el polen ofrecido por la planta, hasta casi completar sus cargas polínicas. Sin embargo, llama la atención que la variedad africana de A. mellifera esté en desventaja competitiva (reflejada en un porcentaje de colecta más bajo) con las especies nativas antes citadas, excepto con M. fasciata. Para A. mellifera existen reportes en la zona templada de una alta constancia floral. Slaa et al., 1998, encontraron que la constancia floral de tres especies de abejas sin aguijón del género Trigona fue menor a la registrada para A. mellifera, pero comparable con la de Apis cerana Fabricius 1793; esos autores proponen que estas diferencias deben obedecer a sus hábitat, dado que las estrategias de pecoreo óptimas cambian de los climas tropicales a los templados y consideran que, en un ambiente altamente diverso como la mayoría de los ecosistemas húmedos tropicales, debe ser más favorable aprovechar la disponibilidad de otros tipos de flores.
Un segundo grupo, incluye a N. mellaria, Lasioglossum sp.117 Scaptotrigona aff. pectoralis y Anthodioctes sp.1. Corresponde a las abejas que dividieron más del 94,5% de sus cargas polínicas entre M. pigra y M. pudica; todas recolectaron porcentajes mayores de granos de polen de M. pudica, excepto N. mellaria, la cual presenta un porcentaje un poco más alto del tipo polínico de M. pigra (Tabla 1). Esta diferencia es importante desde el punto de vista trófico; es decir, el hecho de abastecerse en mayor proporción con polen de M. pudica disminuye la competencia interespecífica por el recurso polínico de M. pigra. Vale la pena destacar a Trigona dorsalis Friese 1900, por recolectar el 97,2% de sus cargas polínicas con granos de polen de M. pigra (59,4%) y de S. diversifolium (37,8%). Igualmente a Lasioglossum sp.113 en cuyas cargas polínicas se encontró 61,8% de P. aduncum, 36,4% de Warszewiczia coccinea y apenas 1,3% de M. pigra. Estas dos especies de abejas constituyen grupos separados en la Figura 3; T. dorsalis colectó diez tipos polínicos (Tabla 1, lo cual permite señalar que su nicho trófico es más amplio que el de Lasioglossum sp.113. Con relación a este último taxón se podría inferir que, aunque M. pigra estaba en floración, cuando la visitó ya tenía las cargas polínicas prácticamente completas, por cuanto se proveyó de granos de polen (98,2%) de las dos plantas antes mencionadas.
En cuanto a la diversidad polínica encontrada en las cargas de las abejas (Tabla 1) y representada en la Fig. 4, sobresalen T. muzoensis y T. dorsalis, especies que conforman el grupo con la mayor riqueza de tipos polínicos (10-12). El segundo grupo incluye a T. nigerrima, T. angustula, S. pectoralis, S. aff. pectoralis, M. fasciata, A. mellifera y Anthodioctes sp.1, con una riqueza de tipos polínicos intermedia (6-8). El tercer grupo, constituido por Lasioglossum sp.113, T. fuscipennis, N. mellaria y Lasioglossum sp.117, presenta la menor riqueza (2-4 tipos polínicos). Existen diferencias estadísticamente significativas entre los grupos primero y tercero; las especies de abejas que integran el tercer grupo podrían considerarse oligolécticas, en tanto que las del primero, polilécticas, ya que visitaron a un mayor número de plantas en busca de polen, una de las recompensas ofrecidas por las flores a sus visitantes. Algunos aspectos importantes del presente trabajo a manera de conclusión son los siguientes:
- El tipo polínico de M. pigra muestra el mayor porcentaje acumulado de colecta y está presente en las cargas polínicas de todas las especies de abejas capturadas visitándola. Esto permite concluir que esta planta pionera, ampliamente distribuida en la zona de Porce II, es una fuente importante de polen para las comunidades de abejas que medran en su hábitat.
- Otras especies vegetales importantes en la dieta de varias de las especies de abejas capturadas son M. pudica, P. aduncum, S. diversifolium, W. coccinea y P. guajava; sus nichos tróficos les permiten explotar estas plantas en floración y como consecuencia de ello disminuye la competencia interespecífica por el recurso polínico ofrecido por M. pigra.
- M. pigra es una especie vegetal polifílica en la cual se observaron visitas de 11 taxones de abejas. Sin embargo, los resultados de este estudio indican la alta constancia floral de T. nigerrima, T. angustula, T. fuscipennis, T. muzoensis y S. pectoralis, en cuyas cargas polínicas se encontró más de 90% de sus granos de polen; acorde con lo anterior, este gremio de abejas tiene una probabilidad más alta de ser el que poliniza a M. pigra, en la zona de influencia del embalse Porce II, aunque el sistema de polinización de esta planta posiblemente involucra a otros insectos.
A la Universidadad Nacional de Colombia, Sede Medellín.
BIBLIOGRAFÍA
AIZEN M, VASQUEZ D, SMITH-RAMIREZ C. Historia natural y conservación de los mutualismos planta-animal del bosque templado de Sudamérica austral. Rev Chil Hist Nat. 2002;75(1):1-33.
[ Links ]ANDRADA A, VALLE A, ARAMAYO E, LAMBERTO S, CANTAMUTTO M. Análisis polínico de las mieles de las sierras australes de la provincia de Buenos Aires, Argentina. Invest agrar Prod prot veg. 1998; 13(3):265-275.
[ Links ]CANO JA. Utilización de un sistema de información geográfico SIG en la elaboración de un plan de uso y manejo de la cobertura vegetal [trabajo de grado: Ingeniero Forestal]. Medellín, Universidad Nacional de Colombia; 1995.
[ Links ]ENDRESS PK. Diversity and evolutionary biology of tropical flowers. Cambridge University Press, Cambridge. 1994; pp. 367-390.
[ Links ]ERDTMAN G. The acetolysis method. A revised description. Svensk Bot Tidskr. 1960;54:561-564.
[ Links ]FONNEGRA R. Métodos de estudio palinológico. Centro de Investigaciones Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, Colombia; 1991.
[ Links ]GIRON M. Análisis palinológico de la miel y la carga de polen colectada por Apis mellifera en el Suroeste de Antioquia, Colombia. Boletín Museo Entomológico Universidad del Valle. 1995;3(2):35-54.
[ Links ]GOLDBLATT P, MANNINNG J, BERNHARDT P. Adaptive radiation of pollination mechanisms in Sparaxis (Iridaceae: Ixioideae). Adansonia. 2000;22(1):57-70.
[ Links ]JONGGJITVIMOL T, WATTANACHAIYINGCHAROEN W. Pollen Food Sources of the Stingless Bees Trigona apicalis Smith, 1857, Trigona collina Smith, 1857 and Trigona fimbriata Smith, 1857 (Apidae, Meliponinae) in Thailand. Nat Hist J Chulalongkorn Univ. 2006;6(2):75-82.
[ Links ]MICHENER CD. The bees of the world. The Johns Hopkins University Press, Baltimore & London; 2000.
[ Links ]MILLS LS, SOULE ME, DOAK DF. The keystone species concept in ecology and conservation. Bioscience. 1993;43(4):219-224.
[ Links ]NATES G, RODRIGUEZ A, PARRA A. El uso de la palinología en estudios con abejas. LAPD Newsletter. 2002;2(1):5.
[ Links ]PROCTOR M, YEO P, LACK A. The Natural History of Pollination. The Bath Press, Great Britain; 1996.
[ Links ]SABUGOSA-MADEIRA B, ABREU I, RIBEIRO H, GOMEZ A, CUNHA M. A scientific note on honey bee foraging activity and airborne pollen Flow. Apidologie. 2007;38:122-123
[ Links ]SLAA EJ, CEVAAL E, SOMMEIJER MJ. Floral constancy in Trigona stingless bees foraging on artificial flower patches: a comparative study. J Apic Res. 1998;37(3):191-198.
[ Links ]SMITH- PARDO AH. Abejas (Hymenoptera: Apoidea) de la zona de influencia del Embalse Porce II (Antioquia, Colombia) [tesis de maestría Maestría en Entomología]. Medellín, Universidad Nacional de Colombia; 1999.
[ Links ]TERRABA A, ANDRÉS C, DÍEZ MJ. Análisis polínico de mieles en el Parque Natural Sierra de Aracena y Picos de Aroche. Botanica Complutensis. 2003;28:121-126.
[ Links ]VERGERON P. Interprétation statistique des resultants en matière d'analyse pollinique des miels. Ann Abeille 1964;7:349-364.
[ Links ]VOGEL S, WESTERKAMP C. Pollination: An integrating factor of biocenoses. In: Seitz A, Loeschcke V. Species consrvation: A population -Biological approach. Birkhäuser, Basel. 1991. p. 159-170.
[ Links ]WILLIAMS NH, WHITTEN WM. Orchid floral fragrances and male euglossine bees: methods and advances in the last sesquidecade. Biol Bull. 1983;164(3):335-395.
[ Links ]
