










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.14 n.3 Bogotá set./dez. 2009
PAPEL DE LAS PROTEÍNAS REGULADORAS Y ACCESORIAS DEL VIH-1 EN LA PATOGÉNESIS DE ESA INFECCIÓN
Role of the regulatory and accesory proteins of HIV-1 in its pathogenisis
XIOMARA ÚSUGA1, M.Sci.; YORLEIDY RUIZ2, Bacterióloga y Laboratorista Clínica, CARLOS JULIO MONTOYA3, D.Sci.; MARÍA TERESA RUGELES4, D.Sci.
1 Grupo de Inmunovirología, Universidad de Antioquia, Medellín
2 Institución Universitaria Colegio Mayor de Antioquia, Medellín
3 Grupo de Inmunovirología, Universidad de Antioquia, Medellín
4 Coordinadora Grupo de Inmunovirología, Universidad de Antioquia, Medellín. Correspondencia: María Teresa Rugeles, Calle 62 N.º 52-59, Torre 2, Laboratorio 532. Sede de Investigación Universitaria, Universidad de Antioquia. Medellín, Colombia. Tel.: (4) 219 6482. Fax: (4) 219 6481. mtrugel@catios.udea.edu.co
Presentado 18 de septiembre de 2008, aceptado 20 de marzo de 2009, correcciones julio 27 de 2009
RESUMEN
Desde el descubrimiento del virus de inmunodeficiencia humana tipo 1 (VIH-1) como agente etiológico del síndrome de inmunodeficiencia adquirida (SIDA) se han descrito los procesos más importantes que hacen parte del ciclo replicativo del virus y que a su vez participan de la fisiopatología tan compleja que caracteriza a esta infección. A pesar de los avances realizados en el desarrollo de medicamentos antirretrovirales y de los logros alcanzados en el control de la replicación viral, hechos que se reflejan en un aumento en la expectativa y calidad de vida de los individuos infectados, la terapia actual no permite una reconstitución inmunológica total y está acompañada de efectos tóxicos secundarios y de la aparición de resistencia viral. Esto ha obligado a mantener la búsqueda constante de nuevos blancos terapéuticos que ofrezcan alternativas en la lucha contra esta pandemia. Hasta hace pocos años se creía que las proteínas accesorias y reguladoras del VIH1 no ejercían un papel significativo en el ciclo replicativo del virus y en la patogénesis de la infección; sin embargo, estudios recientes indican que estas proteínas ejercen funciones esenciales en diferentes etapas del proceso replicativo y por ende son responsables de muchos efectos asociados a la patogénesis viral. Por estos hallazgos, las proteínas accesorias y reguladoras del VIH-1 constituyen un blanco promisorio en el desarrollo de nuevos medicamentos que complementen los antirretrovirales disponibles en la actualidad. En esta revisión se describe la función de las proteínas reguladoras y accesorias del VIH-1 en el ciclo replicativo viral y su participación en el proceso patogénico de esta infección.
Palabras clave: VIH-1, proteínas accesorias, proteínas reguladoras, patogénesis viral.
ABSTRACT
Since the discovery of HIV-1 as the etiological agent of the acquired immunodeficiency syndrome (AIDS), the main processes involved in its replication cycle and responsible for the complex physiopathology of this infection have been described. Despite the advances in the development of new antiretrovirals and their impact in the quality and life expectancy of infected individuals, the current therapy does not allow a complete immune reconstitution and is also associated with deleterious side effects and the appearance of viral resistance. Therefore the search for new therapeutic targets is required to face this pandemic. The role of the accessory and regulatory proteins of the HIV-1 in the replication cycle and in the pathogenesis of the infection has been ignored for several years now; however, recent studies indicated that these proteins play essential roles in the replication cycle, being responsible for several processes associated to viral pathogenesis. These findings have underlined the importance of these proteins as promissory targets in the development of new therapeutic agents. In this review, we detailed the role of each one of the HIV-1’s regulatory and accessory proteins in the replicative cycle and in the pathogenesis of this infection.
Key words: HIV-1, accessory proteins, regulatory proteins, viral pathogenesis
INTRODUCCIÓN
Después de 27 años desde la descripción de los primeros casos de infección por el virus de la inmunodeficiencia humana tipo 1 (VIH-1), aproximadamente 33 millones de personas están infectadas en el mundo y más de 30 millones han muerto como consecuencia de esta infección, con el agravante de que la incidencia de la infección por el VIH-1 continúa en aumento (UNAIDS, 2008).
A partir de todos los estudios realizados buscando esclarecer el ciclo replicativo y la patogénesis viral, se ha acumulado un conocimiento muy valioso que ha permitido el desarrollo de la terapia antirretroviral actual y el establecimiento de otros posibles blancos terapéuticos. Sin embargo, aún se desconocen muchos aspectos de esta infección que siguen siendo objeto de intensa investigación. En los últimos años se han encontrado evidencias que indican el papel que juegan las proteínas accesorias y reguladoras del VIH1 en la patogénesis viral, sugiriendo la posibilidad de intervenir terapéuticamente para alterar su función. A continuación se definirán las funciones que desempeñan las proteínas accesorias Vpu, Vpr, Vif y Nef y las proteínas reguladoras Tat y Rev del VIH-1 durante el ciclo replicativo y la participación de cada una de ellas en la patogénesis de esta infección.
GENERALIDADES DEL VIH-1
El VIH-1, principal agente causal del síndrome de inmunodeficiencia adquirida (SIDA), pertenece al género Lentivirus, subfamilia Orthoretrovirinae, familia Retroviridae. Es un retrovirus esférico de unos 100 nm de diámetro, con tres estructuras superpuestas: una envoltura, una matriz esférica y una cápside cónica que contiene el genoma viral. El VIH-1, como cualquier otro retrovirus, posee como genoma dos moléculas lineares iguales de ARN de cadena sencilla. Además, hacen parte del virión las proteínas Vpr, Nef, Vif y las enzimas transcriptasa reversa (TR), integrasa (IN) y proteasa (PR; Fig. 1).
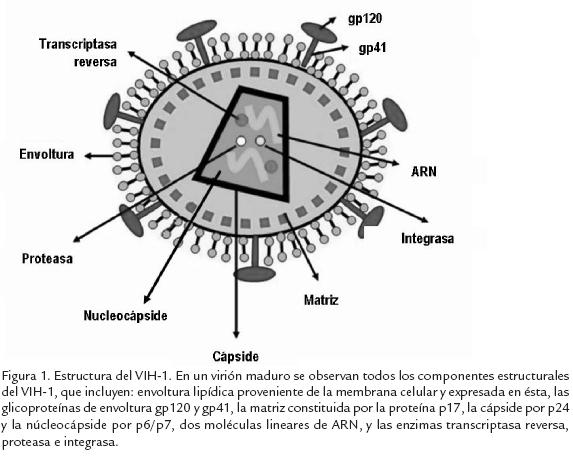
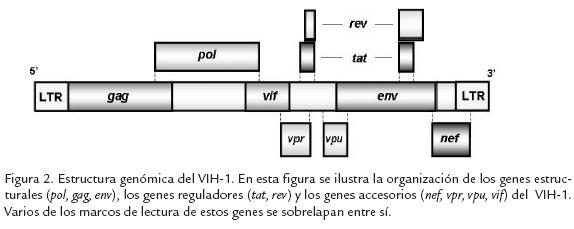
El genoma del VIH-1 contiene nueve genes (Fig. 2). El gen gagcodifica para las proteínas estructurales de la cápside (CA o p24), de la nucleocápside (NC o p6) y de la matriz (MA o p17). El gen polcodifica para las enzimas TR, IN, y PR, mientras que el gen envcodifica para las glicoproteínas gp41 y gp120, que interactúan con las moléculas que permiten la entrada del virus a la célula blanco. Los seis genes restantes codifican para proteínas no estructurales: los genes taty revcodifican para las proteínas reguladoras que reciben el mismo nombre, y los genes vpu, vpr, vify nefcodifican para las proteínas accesorias con el mismo nombre.
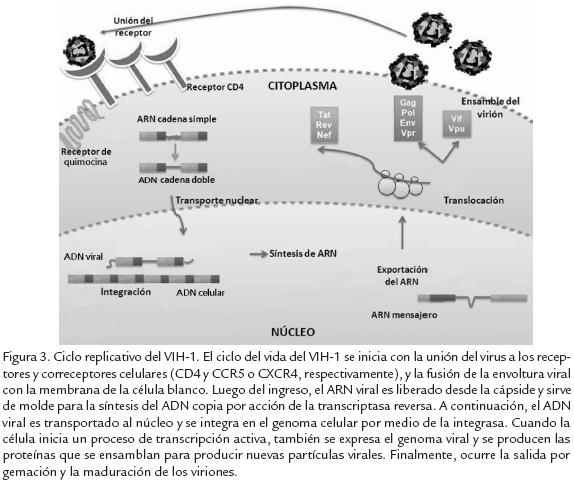
Las proteínas mencionadas anteriormente están implicadas en el proceso de replicación viral, el cual incluye las siguientes etapas (Fig. 3): unión del virión a los receptores de la célula y fusión de la envoltura viral con la membrana celular; entrada de la cápside y liberación del genoma viral al citoplasma; síntesis del ADN copia por la TR; transporte nuclear de este ADN e integración en el genoma de la célula hospedera; transcripción del ARN viral, exportación al citoplasma y síntesis de las proteínas virales; ensamblaje del virión y salida por gemación de las partículas virales, y finalmente maduración extracelular de los viriones.
PATOGÉNESIS DE LA INFECCIÓN POR EL VIH-1
La patogénesis de la infección producida por el VIH-1 es un proceso variable y complejo que depende de factores ambientales, virales y del hospedero. La eliminación masiva de linfocitos T CD4+, característica principal de esta infección, se ha explicado por mecanismos directos e indirectos (Cohen et ál., 2006). Entre los fenómenos directos se encuentran: alteración de la membrana celular y posterior lisis celular, por la salida masiva de viriones o por la interacción de las glicoproteínas de envoltura viral con las moléculas CD4 celulares, ambas expresadas en la membrana celular, mediando eventos de autofusión; toxicidad celular, como consecuencia de la transcripción masiva del genoma viral, ya que la maquinaria metabólica celular queda a disposición de la replicación del virus impidiendo la realización de las funciones vitales, lo que conduce a disfunción celular y muerte posterior; y, finalmente, eliminación de células por medio de la respuesta inmune contra el VIH-1, que incluye la activación de linfocitos T CD8+ citotóxicos que tienen la capacidad de reconocer proteínas virales presentadas en moléculas del complejo mayor de histocompatibilidad clase I (CMH-I) en la superficie de la célula infectada, activando mecanismos efectores que eliminan esas células (Badley et ál., 2000).
De otro lado, entre los mecanismos indirectos se destacan los siguientes: la gp120 soluble puede inducir vías de señalización pro-apoptóticas a través de receptores de quimocinas tales como CXCR4 (Roggero et ál., 2001); el complejo gp120/gp41, anclado en la superficie de células infectadas, puede inducir apoptosis de células no infectadas que expresan las moléculas CD4 y los receptores de quimocinas CXCR4 y CCR5, por tres mecanismos diferentes: intercambio de lípidos de la membrana plasmática por un proceso de hemifusión seguido por una muerte rápida; formación de sincitio por fusión celular (citogamia) y posterior fusión nuclear (cariogamia), originándose estructuras multinucleadas no funcionales que son detectadas por mecanismos de control celular induciendo la eliminación de estas células, y por último, las células infectadas en proceso de apoptosis al fusionarse con células no infectadas transmiten señales apoptóticas, fenómeno conocido como apoptosis “contagiosa” (Andreau et ál., 2004). Adicionalmente, la producción de IFN-α regula positivamente la expresión de TRAIL en los linfocitos T CD4+ y de su receptor de muerte celular, la molécula DR5 (Herbeuval et ál., 2005). Finalmente, la inducción de muerte celular como consecuencia del estado de hiperactivación inmunológica en que se encuentran las células del sistema inmune (Hel et ál., 2006).
Adicionalmente, se ha determinado en los últimos años que las proteínas accesorias y reguladoras del VIH-1 no sólo son indispensables para la replicación del virus, sino que también representan factores de virulencia críticos que han evolucionado para potenciar los efectos citopáticos directos e indirectos responsables de la patogénesis viral (Seelamgari et ál., 2004).
PROTEÍNAS REGULADORAS DEL VIH-1
Tat (Transactivador transcripcional). La regulación de la expresión génica del VIH-1 por el transactivador Tat es un paso crítico en el ciclo replicativo del virus; sin embargo, a diferencia de otros factores transcripcionales que suelen interactuar con el ADN, Tat se une al ARN (Edwards et ál., 2005). La proteína Tat interactúa con una estructura tipo asa que se forma en el ARN viral en transcripción y que está localizada en el extremo 5’ de todos los transcritos virales, la cual se conoce como TAR (elemento de respuesta a la transactivación, del inglés Transactivation response element) y que se encuentra más específicamente en la región R del promotor LTR. Allí Tat induce una modificación de la cromatina al unirse con complejos de proteínas que la remodelan (Pumfery et ál., 2003) y estimula el reclutamiento de otras proteínas para formar un complejo de ribo-nucleoproteínas que favorece la eficiencia de la ARN polimerasa II (D’Orso y Frankel, 2009), participando así en la iniciación y elongación transcripcional, respectivamente.
Además de su importante papel en la replicación viral, Tat también es un inmunomodulador clave en la patogénesis del VIH-1 y sus efectos se observan tanto en células no infectadas como en infectadas debido a que es una proteína que se secreta de la célula. Tat interfiere con la vía de señalización del receptor de IFN-γ a través de la inducción de la actividad de la proteína supresora de la señalización de citocina-2 (SOCS-2, del inglés suppressor of cytokine signaling-2) que impide la fosforilación de STAT1 y la consecuente transcripción de genes específicos regulados por el IFN-γ alterando la respuesta inmune del hospedero y favoreciendo la supervivencia del virus.
De otro lado, Tat se ha relacionado con el daño de la función neuronal observado en la demencia asociada a la infección por VIH-1. En un cultivo primario de astrocitos, la proteína Tat del VIH-1 clada B incrementó los niveles de los agentes neuropatogénicos indoleamina-2,3-dioxigenasa y kinurenina (Samikkannu et ál., 2009). Además, Tat promueve la formación de un complejo macromolecular en la membrana plasmática de las neuronas que incluye las proteínas LRP (del inglés lipoprotein receptor-related protein), PSD95 (del inglés postsynaptic density protein-95), el receptor del N-metil-d-ácido aspártico (NMDA) y la sintasa de óxido nítrico neuronal (nNOS), complejo que desencadena la apoptosis de las neuronas. Estos hallazgos sugieren que Tat puede ser uno de los blancos del tratamiento en la neuropatogénesis de la infección por VIH-1 (Eugenin et ál., 2007; Kim et ál., 2008).
La proteína Tat tiene efectos proinflamatorios al activar el factor transcripcional NF-κB, induciendo la producción de las citocinas y quimocinas MCP-1, RANTES, TNF-α e�IL-6 en un cultivo de astrocitos de ratón (El-Hage et ál., 2008). También, Tat puede hacer parte de los mecanismos responsables de la eliminación de linfocitos T dado que induce la expresión del gen Foxo3a, el cual es un activador transcripcional de los genes fasl y trail involucrados en la vía extrínseca de la apoptosis, y de los genes puma, noxa y bim que participan en la vía intrínseca de la apoptosis. Estos hallazgos indican que el VIH-1 puede inducir la muerte de las células T CD4+ a través de múltiples vías apoptóticas (Dabrowska et ál., 2008).
Rev (Regulador de la expresión viral). Es una proteína esencial para la replicación viral, debido a su función transportadora de los transcritos virales entre el núcleo y el citoplasma. Una vez sintetizada en el citoplasma, Rev pasa al núcleo donde interactúa con la secuencia RRE (del inglés Rev response element) presente en los transcritos virales semiprocesados y sin procesar; esta interacción favorece el reclutamiento de proteínas nucleares, entre ellas Crm1 y Ran-GTP, para formar un complejo de exportación que lleva los diferentes transcritos hacia el citoplasma, para que sean traducidos por los ribosomas en proteínas estructurales y no estructurales del virus (Askjaer et ál., 1998; Emerman y Malim, 1998). Esta proteína es talvez la que aparentemente menos efectos patogénicos ejerce; solo se ha descrito que está presente en el espacio extracelular neuronal y que su presencia allí está relacionada con la demencia asociada a la infección por VIH-1 (van de Bovenkamp et ál., 2002).
PROTEÍNAS ACCESORIAS DEL VIH-1
Vpu (Proteína viral desconocida, del inglés Viral protein unknown). Vpu es una fosfoproteína transmembranal, cuya función principal en el ciclo replicativo es favorecer la liberación de partículas virales desde la superficie de una célula infectada. Esta proteína ejerce varios efectos patogénicos a través de múltiples mecanismos; uno de los más conocidos es la disminución de la expresión en superficie de la molécula CD4, pues Vpu se une a esta molécula luego de que es sintetizada en el retículo endoplasmático (RE) y luego la une a enzimas que facilitan la entrada de la molécula CD4 en la vía de degradación asociada al RE (Lindwasser et ál., 2007). Además, Vpu disminuye la expresión de las moléculas del CMH-I en la superficie de las células infectadas, al retener las cadenas nacientes de este complejo en el RE (Kamp et ál., 2000). Por medio de estos dos mecanismos, el virus puede evadir la respuesta inmune.
Vpu forma canales iónicos selectivos para cationes en las bicapas lipídicas, aumentando la permeabilidad de la membrana celular (Montal, 2003). Aunque varios años atrás se sugirió que la formación de esos canales podría estar relacionada con un incremento en la salida de viriones (Gonzalez y Carrasco, 2001) o con algún requerimiento del virus de una concentración específica de iones intracelulares, no se puede descartar que dichos canales afecten la integridad de la membrana, causando alteraciones en la concentración de cationes intracelulares que afecten algunas vías de señalización. Por ejemplo, las neuronas son células muy sensibles que mueren a consecuencia de cambios iónicos intracelulares. Finalmente, Vpu promueve la apoptosis de linfocitos infectados al inhibir la degradación del inhibidor kappa B, lo cual resulta en la supresión de la actividad del factor NF-κB y en la activación de la vía de las caspasas, al inhibir la expresión de genes antiapoptóticos dependientes de NF-κB tales como los que codifican para Bcl-xL, A1/Bfl-1 y del factor 1 asociado al receptor TNF (Traf1; Akari et ál., 2001).
Nef (Factor negativo, del inglés Negative Factor). Esta es una proteína soluble multifuncional, crítica para la replicación viral in vivo. Su importancia en el ciclo viral se ha observado en algunos individuos infectados, en quienes la alteración funcional de Nef se asoció con cargas virales bajas y con una progresión lenta o no progresión de la infección hacia el SIDA (Anderson y Hope, 2003). Esta proteína, al hacer parte del virión, participa en procesos iniciales de la infección tales como: la entrada viral, promoviendo la fusión a la célula blanco (Schaeffer et ál., 2001); la señalización requerida para la replicación viral, al unirse a cinasas celulares (Renkema y Saksela, 2000; Linnemann et ál., 2002); y la modulación de las balsas de lípidos presentes en la envoltura de los viriones (Brugger et ál., 2007). Aunque no se sabe exactamente a través de cuál mecanismo esta proteína potencia la capacidad infecciosa de las partículas virales y optimiza la replicación viral, se cree que es el resultado de la sumatoria de todas las acciones anteriormente descritas o de otros mecanismos aún desconocidos.
Nef tiene la capacidad de regular negativamente la expresión de las moléculas CD1a, CD1d, CD3, CD28, CD74, CD80, CD86, factor de necrosis tumoral, el receptor de transferrina, la proteína de hemocromatosis HFE y varios miembros de la familia de los receptores de quimocinas (Michel et ál., 2006). Además, Nef modula negativamente la expresión de las moléculas CD4 y del CMH-I, aunque por mecanismos diferentes a los utilizados por Vpu. El efecto de Nef sobre la expresión de CD4 ocurre tempranamente en el ciclo de vida viral; para esto, Nef se une a CD4 e induce su endocitosis por dos mecanismos: rompimiento del complejo CD4-p56 sobre la superficie celular, lo que permite la internalización de CD4 para su degradación por la vía lisosomal; o actuando como enlace entre la molécula CD4 y el complejo de proteínas adaptadoras (Piguetet ál., 1998). La regulación negativa de la expresión en superficie del CMH-I, es ejercida por Nef al incrementar la endocitosis del CMH-I desde la membrana celular e inducir la acumulación de las moléculas endocitadas en el sistema trans-Golgi (Greenberg et ál., 1998). La regulación negativa de todas estás moléculas por Nef contribuye a una alteración de la respuesta inmune y a la evasión de la respuesta mediada por los linfocitos T citotóxicos.
Algunos reportes sugieren que Nef también puede afectar la biosíntesis y el transporte de algunas clases de lípidos celulares tales como el colesterol (van ‘t Wout et ál., 2005), alterando la composición de las balsas de lípidos de la membrana de las células infectadas (Brugger et ál., 2007). Adicionalmente, Nef interactúa con cinasas tales como fosfatidili-nositol-3-cinasa y Lck (Saksela et ál., 1995; Baur et ál., 1997; Linnemann et ál., 2002) y con otras proteínas celulares, lo que le permite interferir con procesos de señalización intracelular (Renkema y Saksela, 2000). Por ejemplo, una de las alteraciones se presenta en la vía de señalización del TCR afectando la activación de las células T. Nef también puede actuar como un inductor intracelular de eventos distales de señalización en la vía del TCR en ausencia de estimulación exógena; sin embargo, en presencia de estimulación exógena del TCR, la señalización es atenuada por esta proteína viral (Schindler et ál., 2006; Fackler et ál., 2007).
Vpr (Proteína viral reguladora, del inglés Viral protein regulatory). La proteína Vpr se encuentra asociada al virión, y presenta tres funciones principales: el transporte del complejo de preintegración (CPI) al núcleo, la retención del ciclo celular en la fase G2 y la expresión del ADN viral no integrado. La acción de Vpr en el transporte del CPI consiste en incrementar la afinidad de la interacción entre este complejo y componentes del poro nuclear, favoreciendo la translocación del CPI desde el citoplasma al núcleo. Vpr no entra al núcleo con el CPI, solo interactúa con proteínas citoplasmáticas y del poro nuclear, como la carioferina-α, la importina-α, la nucleoporina CG1 y hVIP/mov34 (Le Rouzic et ál., 2002; Ramanathan et ál., 2002). Este fenómeno es importante porque permite la integración del ADN viral en células que no se dividen, como monocitos y macrófagos (Le Rouzic et ál., 2002).
Adicionalmente, Vpr está implicada en la inhibición de la expansión clonal al bloquear la división celular deteniendo las células en la fase G2 del ciclo; este bloqueo se da en células que están proliferando, y cuando se prolonga la célula entra en apoptosis . La detención del ciclo celular es mediada por tres mecanismos diferentes: inactivación por fosforilación de la proteína reguladora cdc25, lo que conduce a la inhibición de cdk1/ciclina B y, en última instancia, a la detención en la fase G2; activación de la vía de respuesta ATR ante el ADN dañado, que involucra a las proteínas Rad17 y Hus1 (Roshal et ál., 2003); a través de la regulación negativa de la cinasa MEK2 (Yoshizuka et ál., 2005). El bloqueo del ciclo celular mediado por Vpr proporciona el tiempo necesario para la síntesis de proteínas virales y el ensamblaje del virión (Hrimech et ál., 2000). La proteína Vpr, expresada de novo o liberada desde los viriones luego de la entrada viral, participa en la expresión del ADN viral no integrado. Este proceso no requiere localización nuclear de la proteína viral, ni la detención del ciclo celular mediado por Vpr (Poon y Chen, 2003). Posterior a la activación de esa expresión ejercida por Vpr, se incrementa la transcripción del ARN sin procesar de gag y el ARN procesado de nef (Poon et ál., 2007). Como otro mecanismo patogénico, Vpr exhibe una acción moduladora dual de la apoptosis, inhibiéndola o estimulándola, por medio de varios mecanismos: suprime la actividad del NF-κB y, por consiguiente, disminuye la expresión de las citocinas IL-2, IL4, IL-10, IL-12 y TNF-α (Seelamgari et ál., 2004); regula positivamente la molécula antiapoptótica BcL-2 y otros genes relacionados con la apoptosis (Patel et ál., 2002); forma canales iónicos en las bicapas lipídicas, permitiendo la entrada de sodio y creando un desbalance catiónico (Piller et ál., 1999); y altera la permeabilidad de la membrana mitocondrial al interactuar con el complejo del poro de transición. Su acción proapoptótica se ha relacionado con muerte neuronal y con el desarrollo de encefalopatías en paciente infectados por el VIH-1 (Jones et ál., 2007), mientras que su acción antiapoptótica sobre las células vecinas puede conducir a la preservación de un conjunto de células susceptibles facilitando la persistencia y expansión del virus.
Vif (Factor de infectividad viral, del inglés viral infectivity factor). La proteína Vif aunque hace parte del virión, también es expresada en las etapas tardías de la infección. Estudios sobre la función que ejerce Vif durante la infección por el VIH-1 sugieren que actúa desde las etapas iniciales de la infección hasta la síntesis de proteínas virales, debido a que virus mutados en Vif exhiben una baja actividad de la TR, una capacidad reducida en el proceso de elongación del ADN viral y de la síntesis de proteínas del VIH-1. Aunque se ha demostrado que Vif tiene la capacidad de unirse al ARN viral, no se conoce el mecanismo por el cual podría participar en la transcripción reversa (Dettenhofer et ál., 2000). Vif posee un dominio 90RKKR93, el cual se asemeja a una señal de localización nuclear, lo que sugiere que esta proteína viral está implicada en la regulación del tráfico del CPI al núcleo, posiblemente inhibiendo su transporte (Friedler et ál., 1999). Por otro lado, esta proteína ha sido relacionada con el ensamblaje e infectividad de la progenie viral, pues una fracción importante de Vif está pre-sente en el sitio de la membrana plasmática en el que tiene lugar el ensamblaje del virus (Mitsuya et ál., 1990). Trabajos recientes han demostrado que la proteína Vif induce un retardo en la fase G2 del ciclo celular a través de la inducción de la ubiquitinación y degradación de una proteína celular desconocida, la cual es requerida para la eficiente transición de la fase G2 a M, y que es diferente de APOBEC3D/E, G y F (DeHart et ál., 2008).
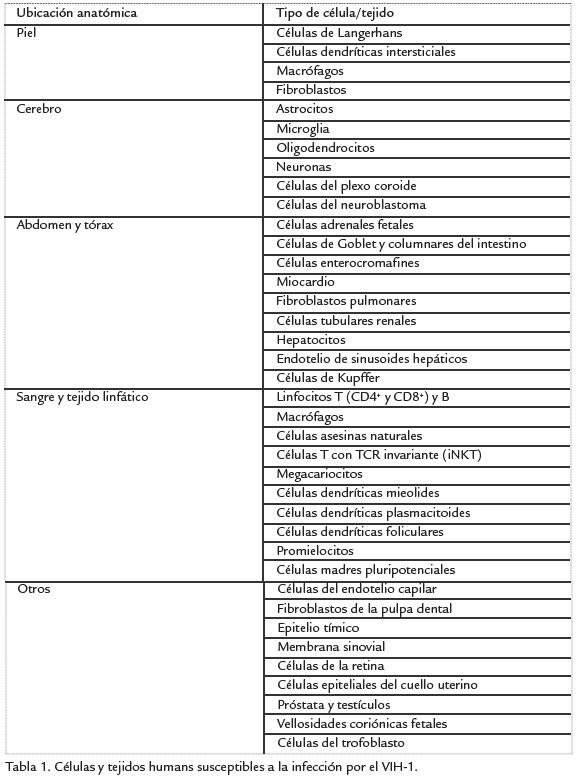
La proteína Vif evita el reconocimiento del sistema inmune innato al prevenir la encapsidación de las proteínas APOBEC en las partículas virales, por medio de la inducción de la poliubiquitinación y degradación mediada por el proteasoma de APOBEC (Goila-Gaur y Strebel, 2008). Los miembros de la familia APOBEC corresponden a deaminasas de citidina celulares, cuya función es inducir mutaciones en el genoma viral que afecten la capacidad infecciosa de los virus, desencadenando inmunidad innata contra la infección por retrovirus y protegiendo a las células de retro-elementos móviles endógenos. La mayoría de efectos patogénicos de las proteínas accesorias y reguladoras del VIH-1 se han descrito en algunas de las células de origen hematopoyético y del sistema nervioso central, las cuales son susceptibles a la infección y en ellas el VIH-1 se replica (infección productiva) aunque de manera variable según el tipo de célula. Sin embargo, en el humano el rango de células y tejidos susceptibles a la infección por el VIH-1 es más amplio (Tabla 1); y aunque en otras células la replicación del VIH1 no es significativa (infección no productiva), se ha sugerido la posibilidad de que ellas también sufran los efectos patogénicos de las proteínas del VIH-1, aumentando la gama de alteraciones tisulares inducidas por la infección con el VIH-1 en el organismo.
PROTEíNAS REGULADORAS Y ACCESORIAS COMO BLANCO TERAPéUTICO
El desarrollo de una terapia antiviral contra el VIH-1 ha sido objetivo de una intensainvestigación desde el descubrimiento de este virus. Hasta el momento, se han desa-rrollado medicamentos que mejoran la calidad de vida y prolongan significativamente la12 expectativa de vida de las personas infectadas, al inhibir las enzimas virales y bloquear potentemente la capacidad del virus para replicarse. Sin embargo, los medicamentos actuales están asociados con marcados efectos secundarios, resistencia viral al tratamiento, persistencia de reservorios virales, regímenes complicados para la toma del tratamiento y un costo alto ; por lo tanto, hay una búsqueda constante de nuevos blancos terapéuticos. Teniendo en cuenta el conocimiento actual sobre el papel que ejercen las proteínas reguladoras y accesorias del VIH-1 en su ciclo replicativo y la importancia de éstas como factores de virulencia, estas moléculas se han convertido en un blanco terapéutico muy importante. El uso potencial de estas proteínas virales en protocolos de inmunoterapia tiene algunas ventajas tales como: intervención en una etapa más temprana del ciclo viral, su poca variabilidad y su inmunogenicidad. Sin embargo, los estudios en este campo apenas se están iniciando.
La principal estrategia está en encontrar moléculas que permitan apuntar a la inhibición de la síntesis de estas proteínas virales, a su degradación, a la inhibición de su función o al bloqueo de la interacción entre las proteínas virales y las celulares. El desarrollo tecnológico ha permitido encontrar componentes promisorios, como pequeñas moléculas naturales y sintéticas, péptidos, proteínas y oligonucleótidos como el ARN de interferencia. Sin embargo, el uso de estos componentes para la inhibición de las proteínas accesorias y reguladoras del VIH-1 es aún un campo en desarrollo, que todavía no ha ido más allá de la experimentación in vitro. Hasta el momento se han identificado los siguientes compuestos: la hematoxilina como un supresor de la interacción entre Vpr y la importina-α, inhibiendo la traslocación del CPI (Suzuki et ál., 2009); el damnacanthal, un componente del noni (Morinda citrifolia) que inhibe la muerte celular inducida por Vpr (Kamata et ál., 2006); y la quercetina, un flavonoide proveniente de la planta Houttuynia que demostró su capacidad para alterar la función de Vpr (Shimura et ál., 1999). Adicionalmente, como inhibidor de la interacción Tat-TAR se desarrolló una molécula sintética llamada TR87, la cual se une a TAR (Hwang et ál., 2003).
Por otro lado, la importancia de la inmunidad humoral y celular para controlar o prevenir la progresión de la infección, ha conducido a desarrollar vacunas terapeúticas o profilácticas que incluyen proteínas reguladoras y/o accesorias para inducir una respuesta inmune contra ellas. Estas vacunas están basadas en el uso de plásmidos de ADN o de vectores virales, de proteínas inactivas o biológicamente activas, e incluso de péptidos. Algunas de ellas han sido investigadas en ensayos clínicos y pre-clínicos, y en general han demostrado ser seguras, buenas inductoras de linfocitos T citotóxicos y T ayudadores, permitiendo controlar los niveles de viremia (Ferrantelli et ál., 2004).
CONCLUSIÓN
El estudio de la patogénesis del VIH-1 sigue siendo un área de intensa investigación. Los trabajos realizados hasta el momento han permitido una mejor compresión del ciclo replicativo viral y de los mecanismos por medio de los cuales se da la eliminación masiva de los linfocitos T CD4+, fenómeno que finalmente lleva al estado de inmunodeficiencia característico del SIDA. Este conocimiento ha sido la base fundamental para la implementación de las estrategias terapéuticas que están actualmente en uso clínico
o en fases avanzadas de su desarrollo. Sin embargo, los efectos secundarios asociados al uso de los antirretrovirales disponibles y la aparición de cepas virales resistentes hacen prioritaria la búsqueda de nuevos blancos terapéuticos. Aunque desde hace muchos años se reconoce la importancia de las proteínas reguladoras en el proceso de replicación del VIH-1, hasta hace pocos años se creía que las proteínas accesorias del VIH1 no tenían un papel significativo durante el ciclo replicativo del virus. Sin embargo, estudios recientes indican que estas proteínas, al igual que las proteínas reguladoras, ejercen funciones importantes en diferentes etapas del proceso replicativo. Algunas de sus acciones son realizadas por la interacción con factores celulares, intervención en vías de señalización, modulación de la apoptosis y regulación de la expresión de moléculas de superficie. Todos estos efectos finalmente potencian la replicación viral y favorecen la evasión de la respuesta inmune. Por estas razones, las proteínas reguladoras y accesorias se constituyen en un blanco potencial para drogas antivirales que podrían llegar a ser el complemento ideal de la terapia actual.
BIBLIOGRAFÍA
AKARI H, BOUR S, KAO S, ADACHI A, STREBEL K. The human immunodeficiency virus type 1 accessory protein Vpu induces apoptosis by suppressing the nuclear factor kappaB-dependent expression of antiapoptotic factors. J Exp Med. 2001;194(9):1299-1311.
[ Links ]ANDERSON JL, HOPE TJ. Recent Insights into HIV Accessory Proteins. Curr Infect Dis Rep. 2003;5(5):439-450.
[ Links ]ANDREAU K, PERFETTINI JL, CASTEDO M, METIVIER D, SCOTT V, PIERRON G, et ál. Contagious apoptosis facilitated by the HIV-1 envelope: fusion-induced cell-to-cell transmission of a lethal signal. J Cell Sci. 2004;117(Pt 23):5643-5653.
[ Links ]ASKJAER P, JENSEN TH, NILSSON J, ENGLMEIER L, KJEMS J. The specificity of the CRM1-Rev nuclear export signal interaction is mediated by RanGTP. J Biol Chem. 1998;273(50):33414-33422.
[ Links ]BADLEY AD, PILON AA, LANDAY A, LYNCH DH. Mechanisms of HIV-associated lymphocyte apoptosis. Blood. 2000;96(9):2951-2964.
[ Links ]BAUR AS, SASS G, LAFFERT B, WILLBOLD D, CHENG-MAYER C, PETERLIN BM. The N-terminus of Nef from HIV-1/SIV associates with a protein complex containing Lck and a serine kinase. Immunity. 1997;6(3):283-291.
[ Links ]BRUGGER B, KRAUTKRAMER E, TIBRONI N, MUNTE CE, RAUCH S, LEIBRECHT I, et ál. Human immunodeficiency virus type 1 Nef protein modulates the lipid composition of virions and host cell membrane microdomains. Retrovirology. 2007;4:70.
[ Links ]COHEN OC, VACCAREZZA M, FAUCI A. Inmunología de la Infección por el Virus de la Inmunodeficiencia Humana. Philadelphia: Elsevier; 2006.
[ Links ]DORSO I, FRANKEL AD. Tat acetylation modulates assembly of a viral-host RNA-protein transcription complex. Proc Natl Acad Sci U S A 2009;106(9):3101-3106.
[ Links ]DABROWSKA A, KIM N, ALDOVINI A. Tat-induced FOXO3a is a key mediator of apoptosis in HIV-1-infected human CD4+ T lymphocytes. J Immunol. 2008;181(12):8460-8477.
[ Links ]DEHART JL, BOSQUE A, HARRIS RS, PLANELLES V. Human immunodeficiency virus type 1 Vif induces cell cycle delay via recruitment of the same E3 ubiquitin ligase complex that targets APOBEC3 proteins for degradation. J Virol. 2008;82(18):9265-9272.
[ Links ]DETTENHOFER M, CEN S, CARLSON BA, KLEIMAN L, YU XF. Association of human immunodeficiency virus type 1 Vif with RNA and its role in reverse transcription. J Virol. 2000;74(19):8938-8945.
[ Links ]EDWARDS TE, ROBINSON BH, SIGURDSSON ST. Identification of amino acids that promote specific and rigid TAR RNA-tat protein complex formation. Chem Biol. 2005;12(3):329-337.
[ Links ]EL-HAGE N, BRUCE-KELLER AJ, KNAPP PE, HAUSER KF. CCL5/RANTES gene deletion attenuates opioid-induced increases in glial CCL2/MCP-1 immunoreactivity and activation in HIV-1 Tat-exposed mice. J Neuroimmune Pharmacol. 2008;3(4):275-285.
[ Links ]EMERMAN M, MALIM MH. HIV-1 regulatory/accessory genes: keys to unraveling viral and host cell biology. Science. 1998;280(5371):1880-1884.
[ Links ]EUGENIN EA, KING JE, NATH A, CALDERON TM, ZUKIN RS, BENNETT MV, et ál. HIV-tat induces formation of an LRP-PSD-95-NMDAR-nNOS complex that promotes apoptosis in neurons and astrocytes. Proc Natl Acad Sci USA. 2007;104(9):3438-3443.
[ Links ]FACKLER OT, ALCOVER A, SCHWARTZ O. Modulation of the immunological synapse: a key to HIV-1 pathogenesis? Nat Rev Immunol. 2007;7(4):310-317.
[ Links ]FERRANTELLI F, CAFARO A, ENSOLI B. Nonstructural HIV proteins as targets for prophylactic or therapeutic vaccines. Curr Opin Biotechnol. 2004;15(6):543-556.
[ Links ]FRIEDLER A, ZAKAI N, KARNI O, FRIEDLER D, GILON C, LOYTER A. Identification of a nuclear transport inhibitory signal (NTIS) in the basic domain of HIV-1 Vif protein. J Mol Biol. 1999;289(3):431-437.
[ Links ]GOILA-GAUR R, STREBEL K. HIV-1 Vif, APOBEC, and intrinsic immunity. Retrovirology. 2008;5:51.
[ Links ]GONZALEZ ME, CARRASCO L. Human immunodeficiency virus type 1 VPU protein affects Sindbis virus glycoprotein processing and enhances membrane permeabilization. Virology. 2001;279(1):201-209.
[ Links ]GREENBERG ME, IAFRATE AJ, SKOWRONSKI J. The SH3 domain-binding surface and an acidic motif in HIV-1 Nef regulate trafficking of class I MHC complexes. Embo J. 1998;17(10):2777-2789.
[ Links ]HEL Z, MCGHEE JR, MESTECKY J. HIV infection: first battle decides the war. Trends Immunol. 2006;27(6):274-281.
[ Links ]HERBEUVAL JP, GRIVEL JC, BOASSO A, HARDY AW, CHOUGNET C, DOLAN MJ, et ál. CD4+ T-cell death induced by infectious and noninfectious HIV-1: role of type 1 interferon-dependent, TRAIL/DR5-mediated apoptosis. Blood. 2005;106(10):3524-3531.
[ Links ]HRIMECH M, YAO XJ, BRANTON PE, COHEN EA. Human immunodeficiency virus type 1 Vpr-mediated G(2) cell cycle arrest: Vpr interferes with cell cycle signaling cascades by interacting with the B subunit of serine/threonine protein phosphatase 2A. Embo J. 2000;19(15):3956-3967.
[ Links ]HWANG S, TAMILARASU N, KIBLER K, CAO H, ALI A, PING YH, et ál. Discovery of a small molecule Tat-trans-activation-responsive RNA antagonist that potently inhibits human immunodeficiency virus-1 replication. J Biol Chem. 2003;278(40):39092-39103.
[ Links ]JONES GJ, BARSBY NL, COHEN EA, HOLDEN J, HARRIS K, DICKIE P, et ál. HIV1 Vpr causes neuronal apoptosis and in vivo neurodegeneration. J Neurosci. 2007;27(14):3703-3711.
[ Links ]KAMATA M, WU RP, AN DS, SAXE JP, DAMOISEAUX R, PHELPS ME, et ál. Cell-based chemical genetic screen identifies damnacanthal as an inhibitor of HIV-1 Vpr induced cell death. Biochem Biophys Res Commun. 2006;348(3):1101-1106.
[ Links ]KAMP W, BERK MB, VISSER CJ, NOTTET HS. Mechanisms of HIV-1 to escape from the host immune surveillance. Eur J Clin Invest. 2000;30(8):740-746.
[ Links ]KIM HJ, MARTEMYANOV KA, THAYER SA. Human immunodeficiency virus protein Tat induces synapse loss via a reversible process that is distinct from cell death. J Neurosci. 2008;28(48):12604-12613.
[ Links ]LE ROUZIC E, MOUSNIER A, RUSTUM C, STUTZ F, HALLBERG E, DARGEMONT C, et ál. Docking of HIV-1 Vpr to the nuclear envelope is mediated by the interaction with the nucleoporin hCG1. J Biol Chem. 2002;277(47):45091-45098.
[ Links ]LINDWASSER OW, CHAUDHURI R, BONIFACINO JS. Mechanisms of CD4 downregulation by the Nef and Vpu proteins of primate immunodeficiency viruses. Curr Mol Med. 2007;7(2):171-184.
[ Links ]LINNEMANN T, ZHENG YH, MANDIC R, PETERLIN BM. Interaction between Nef and phosphatidylinositol-3-kinase leads to activation of p21-activated kinase and increased production of HIV. Virology. 2002;294(2):246-255.
[ Links ]MICHEL N, GANTER K, VENZKE S, BITZEGEIO J, FACKLER OT, KEPPLER OT. The Nef protein of human immunodeficiency virus is a broad-spectrum modulator of chemokine receptor cell surface levels that acts independently of classical motifs for receptor endocytosis and Galphai signaling. Mol Biol Cell. 2006;17(8):3578-3590.
[ Links ]MITSUYA H, YARCHOAN R, BRODER S. Molecular targets for AIDS therapy. Science. 1990;249(4976):1533-1544.
[ Links ]MONTAL M. Structure-function correlates of Vpu, a membrane protein of HIV-1. FEBS Lett. 2003;552(1):47-53.
[ Links ]PATEL CA, MUKHTAR M, HARLEY S, KULKOSKY J, POMERANTZ RJ. Lentiviral expression of HIV-1 Vpr induces apoptosis in human neurons. J Neurovirol. 2002;8(2):86-99.
[ Links ]PIGUET V, CHEN YL, MANGASARIAN A, FOTI M, CARPENTIER JL, TRONO D. Mechanism of Nef-induced CD4 endocytosis: Nef connects CD4 with the mu chain of adaptor complexes. Embo J. 1998;17(9):2472-2481.
[ Links ]PILLER SC, EWART GD, JANS DA, GAGE PW, COX GB. The amino-terminal region of Vpr from human immunodeficiency virus type 1 forms ion channels and kills neurons. J Virol. 1999;73(5):4230-4238.
[ Links ]POON B, CHANG MA, CHEN IS. Vpr is required for efficient Nef expression from unintegrated human immunodeficiency virus type 1 DNA. J Virol. 2007;81(19):10515-10523.
[ Links ]POON B, CHEN IS. Human immunodeficiency virus type 1 (HIV-1) Vpr enhances expression from unintegrated HIV-1 DNA. J Virol. 2003;77(7):3962-3972.
[ Links ]PUMFERY A, DENG L, MADDUKURI A, DE LA FUENTE C, LI H, WADE JD, et ál. Chromatin remodeling and modification during HIV-1 Tat-activated transcription. Curr HIV Res. 2003;1(3):343-362.
[ Links ]RAMANATHAN MP, CURLEY E, SU M, CHAMBERS JA, WEINER DB. Carboxyl terminus of hVIP/mov34 is critical for HIV-1-Vpr interaction and glucocorticoid-mediated signaling. J Biol Chem. 2002;277(49):47854-47860.
[ Links ]RENKEMA GH, SAKSELA K. Interactions of HIV-1 NEF with cellular signal transducing proteins. Front Biosci. 2000;5:D268-283.
[ Links ]ROGGERO R, ROBERT-HEBMANN V, HARRINGTON S, ROLAND J, VERGNE L, JALECO S, et ál. Binding of human immunodeficiency virus type 1 gp120 to CXCR4 induces mitochondrial transmembrane depolarization and cytochrome c-mediated apoptosis independently of Fas signaling. J Virol. 2001;75(16):7637-7650.
[ Links ]ROSHAL M, KIM B, ZHU Y, NGHIEM P, PLANELLES V. Activation of the ATR-mediated DNA damage response by the HIV-1 viral protein R. J Biol Chem. 2003;278(28):25879-25886.
[ Links ]SAKSELA K, CHENG G, BALTIMORE D. Proline-rich (PxxP) motifs in HIV-1 Nef bind to SH3 domains of a subset of Src kinases and are required for the enhanced growth of Nef+ viruses but not for down-regulation of CD4. Embo J. 1995;14(3):484-491.
[ Links ]SAMIKKANNU T, SAIYED ZM, RAO KV, BABU DK, RODRIGUEZ JW, PAPUASHVILI MN, et ál. Differential regulation of indoleamine-2,3-dioxygenase (IDO) by HIV type 1 clade B and C Tat protein. AIDS Res Hum Retroviruses. 2009;25(3):329-335.
[ Links ]SCHAEFFER E, GELEZIUNAS R, GREENE WC. Human immunodeficiency virus type 1 Nef functions at the level of virus entry by enhancing cytoplasmic delivery of virions. J Virol. 2001;75(6):2993-3000.
[ Links ]SCHINDLER M, MUNCH J, KUTSCH O, LI H, SANTIAGO ML, BIBOLLETRUCHE F, et ál. Nef-mediated suppression of T cell activation was lost in a lentiviral lineage that gave rise to HIV-1. Cell. 2006;125(6):1055-1067.
[ Links ]SEELAMGARI A, MADDUKURI A, BERRO R, DE LA FUENTE C, KEHN K, DENG L, et ál. Role of viral regulatory and accessory proteins in HIV-1 replication. Front Biosci. 2004;9:2388-2413.
[ Links ]SHIMURA M, ZHOU Y, ASADA Y, YOSHIKAWA T, HATAKE K, TAKAKU F, et ál. Inhibition of Vpr-induced cell cycle abnormality by quercetin: a novel strategy for searching compounds targeting Vpr. Biochem Biophys Res Commun. 1999;261(2):308-316.
[ Links ]SUZUKI T, YAMAMOTO N, NONAKA M, HASHIMOTO Y, MATSUDA G, TAKESHIMA SN, et ál. Inhibition of human immunodeficiency virus type 1 (HIV-1) nuclear import via Vpr-Importin alpha interactions as a novel HIV-1 therapy. Biochem Biophys Res Commun. 2009;380(4):838-843.
[ Links ]UNAIDS. 2008 Report on the global AIDS epidemic. 2008 [4 de abril de 2009]; Disponible en: http://www.unaids.org/en/KnowledgeCentre/HIVData/GlobalReport/ 2008/2008_Global_report.asp
[ Links ]VAN T WOUT AB, SWAIN JV, SCHINDLER M, RAO U, PATHMAJEYAN MS, MULLINS JI, et ál. Nef induces multiple genes involved in cholesterol synthesis and uptake in human immunodeficiency virus type 1-infected T cells. J Virol. 2005;79(15):10053-10058.
[ Links ]VAN DE BOVENKAMP M, NOTTET HS, PEREIRA CF. Interactions of human immunodeficiency virus-1 proteins with neurons: possible role in the development of human immunodeficiency virus-1-associated dementia. Eur J Clin Invest. 2002;32(8):619-627.
[ Links ]YOSHIZUKA N, YOSHIZUKA-CHADANI Y, KRISHNAN V, ZEICHNER SL. Human immunodeficiency virus type 1 Vpr-dependent cell cycle arrest through a mitogen-activated protein kinase signal transduction pathway. J Virol. 2005;79(17):11366-11381.
[ Links ]
