










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.14 supl.1 Bogotá dez. 2009
EL PROCESO EVOLUTIVO EVOLUCIONA: DEL GENOMA AL SOCIOMA Y VUELTA
The Evolutionary Process Evolves: From the Genome to the Sociome and Back
JOAO MUÑOZ-DURÁN1, Ph. D. 1Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá. jvmunozd@unal.edu.co
Presentado 27 de noviembre de 2009, aceptado 5 de marzo de 2010, correcciones 25 de mayo de 2010.
RESUMEN
Una revisión de las principales características de los sistemas biológicos muestra la imposibilidad de una reducción ontológica y epistemológica para su estudio. Lo anterior contrasta con la forma simplista, genocéntrica, en forma predominante se analiza la evolución de dichos sistemas. Una alternativa que podría conciliar el estudio de los fenómenos evolutivos con las características más notorias de los sistemas biológicos es la hipótesis de “selección jerárquica”, que ha sido debatida, pero no lo suficientemente desarrollada. Propongo que de esta hipótesis se derivan las siguientes implicaciones. Primero, el mecanismo de evolución evoluciona a medida que surgen innovaciones y se conforman nuevos niveles de organización. Es así como en el transcurso de la evolución han surgido de manera secuencial tres niveles en los cuales opera la selección: genes, organismos y grupos de organismos que cooperan. Los organismos son la unidad preponderante de selección, y con su emergencia los genes, considerados aisladamente, dejaron de ser objetos directos de selección. El nivel de selección más reciente es el de los grupos de organismos que cooperan, cuyo origen coincide con la evolución del cuidado parental y de estrategias de enseñanza y aprendizaje. Segundo, existen por lo menos dos maneras interdependientes en que la información relevante para el proceso evolutivo se codifica y almacena: el genoma y el “socioma”. Propongo el concepto de “socioma” como un nivel en el que ocurren procesos evolutivos complementarios a los que suceden en el genoma. Como argumentaré, la acumulación a lo largo de múltiples generaciones de alteraciones en el socioma tiene como efecto la relajación, el reforzamiento o la generación de nuevas presiones de selección. Entre el genoma y el socioma se establece un bucle de afectación recíproca que impulsa la evolución de numerosos grupos taxonómicos.
Palabras clave: complejidad, selección jerárquica, propiedades emergentes, socioma, interacción genomasocioma.
ABSTRACT
A review of the main characteristics of biological systems shows that they are irreducible from ontological and epistemological stand points. In contrast, the evolution of these systems is predominantly studied from simplistic, genocentric approaches. An alternative that may conciliate the study of evolution with the main characteristics of biological systems is the hierarchical selection hypothesis, which has been discussed, but not developed enough. I propose that from this hypothesis the following implications can be derived. First, the evolutionary process evolves itself simultaneously with the origin of evolutionary novelties and the establishment of new levels of biological organization. Three levels of selection have evolved: genes, organisms, and groups of organisms that cooperate. Organisms are the preponderant units of selection and since their evolutionary emergence, genes considered as isolated units are no longer direct objects of selection. Groups of organisms that cooperate constitute the most recent level of selection. Its origin is associated with the evolution of parental care, and the emergence of learning and teaching strategies. Second, there are two interdependent ways in which relevant information for the evolutionary process is encoded and transmitted: genome and sociome. I propose the sociome concept as a level in which evolutionary process take place that are complementary to those observed in the genome. I argue that the accumulation of modifications at the sociome level along multiple generations relaxes, strengthens or creates new selective pressures. There is a loop of reciprocal affectation between the genome and sociome that promotes evolution in several taxa.
Key words: complexity, hierarchical selection, emergent properties, sociome, interaction genomesociome.
LA ESENCIA DE LOS SISTEMAS BIOLÓGICOS RESIDE EN LA INTERACCIÓN Y, POR TANTO, SON IRREDUCIBLES DESDE EL PUNTO DE VISTA ONTOLÓGICO Y EPISTEMOLÓGICO
El objetivo de esta sección es revisar algunas de las características de los sistemas complejos y la manera como estas se expresan en los sistemas biológicos. Esto servirá de contexto para argumentar por qué las aproximaciones genocentristas en biología son insuficientes para entender la estructura, organización y evolución de los sistemas biológicos. Algunos de los ejemplos que utilizo para sustentar mis afirmaciones a lo largo de este trabajo pueden ser objeto de interpretaciones diferentes, como cabría esperarse durante el desarrollo de un debate académico fértil y justo.
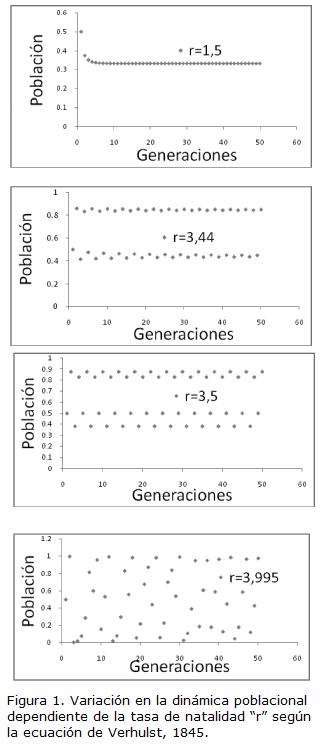
NoLinealidad. Las ecuaciones que describen los sistemas complejos incluyen factores que se relacionan de manera no lineal. Esta propiedad los hace sensibles a cambios sutiles en los valores de algunos parámetros y a ligeras perturbaciones en las condiciones iniciales del sistema, lo que dificulta predecir su comportamiento futuro. Así, los sistemas complejos, y entre ellos los biológicos, se pueden comportar de manera organizada o, por el contrario, lo pueden hacer de manera inestable y errática (Nicolis y Prigogine, 1989). Un ejemplo sencillo en el campo de la física es un conjunto de moléculas de agua bajo condiciones variables de temperatura y presión. Bajo ciertos valores de estas variables las moléculas de agua forman cristales, una estructura ordenada. Esas mismas moléculas, bajo condiciones diferentes de temperatura y presión, pueden moverse de manera aleatoria en las fases líquida y gaseosa. En biología un ejemplo clásico es la ecuación de Verhulst (1845; Briggs y Peat, 1991) que describe de manera sencilla el crecimiento de una población que depende de dos factores: la tasa de natalidad “r” y el tamaño de la población en un instante “N”. En la Figura. 1 podemos ver que para valores de la tasa de natalidad menores a tres (r < 3) es posible predecir el tamaño de la población en cualquier generación. Cuando “r” adquiere valores dentro de un determinado rango (3 > r < 3,4495) la población oscila alrededor de dos valores estables. Para valores de “r” superiores al rango anterior y menores a 3,7, la población oscila alrededor de cuatro valores. Para valores de “r” entre 3,7 y 4,0 aparecen nuevas bifurcaciones y la población se comporta de manera caótica, con fluctuaciones aleatorias, lo que dificulta hacer predicciones del tamaño poblacional. Lo anterior muestra que los sistemas biológicos, como todos los sistemas complejos, no son de por sí ordenados o caóticos; pueden ser lo uno o lo otro, dependiendo de las condiciones bajo las cuales se desarrollan.
Numerosas partes constitutivas, densa red de interacciones y mecanismos de retroalimentación positiva y negativa. Esta propiedad hace que los sistemas complejos sean complicados de describir, entender, modelar y explicar. Probablemente esta es la característica que ha estimulado el desarrollo de aproximaciones reduccionistas para estudiar los sistemas complejos. No se me ocurre un sistema biológico que no exprese esta propiedad, desde el nivel de los elementos que componen una membrana celular, pasando por la organización a nivel celular, las comunidades de microorganismos que se pueden encontrar bajo una roca cualquiera, las colonias de insectos sociales, las sociedades humanas y la organización de comunidades ecológicas a escalas geográficas.
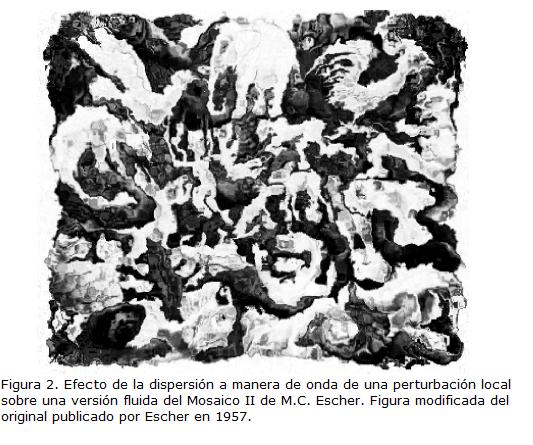
Conectividad entre las partes constitutivas. Dada esta propiedad, cualquier perturbación local será propagada mediante la red de interacciones e interdependencias y así afectará, de manera directa o indirecta, a todos los elementos constituyentes del sistema. Mauritis Cornelis Escher en 1957 elaboró una litografía titulada “Mosaico II” que me parece ilustra bastante bien la conectividad de los sistemas biológicos, aunque no la describe formalmente. Podríamos tomar este mosaico como la representación parcial de un ecosistema. En esta obra hay figuras humanas y animales que podrían representar la diversidad de fauna en un ecosistema; también hay figuras humanoides de animales míticos, e incluso de una guitarra, que podrían reflejar aspectos de la cultura humana. Las figuras están pintadas unas en blanco y otras en negro, y es interesante notar que todas las de un mismo color están conectadas como en una sucesión alrededor del cuadro; así, pareciera que cada figura es la extensión, la continuación de las figuras del mismo color con las que se enlaza. Más aún, las figuras de un color delinean, dan forma, son causa y razón de las figuras del color opuesto. Las figuras negras emergen de la conexión y continuidad de las figuras blancas y viceversa.
Los sistemas biológicos están organizados y estructurados de manera muy semejante a la anterior interpretación del Mosaico II de Escher, solo que en este caso el mosaico no está elaborado sobre una superficie rígida, sino sobre una superficie fluida. Imaginemos el mosaico pintado sobre la superficie de una piscina, primero aislada del entorno de tal manera que las figuras conservan su forma y posición relativa. Ahora imaginemos que una gota de agua cae sobre algún punto de la superficie liquida. Esto genera una perturbación que se dispersa en forma de onda y distorsiona la silueta de una figura inicial y después la de las figuras vecinas. A medida que la onda viaja las alteraciones son cada vez mayores. De esta manera, incluso piezas que nunca estuvieron en contacto directo se afectan las unas a las otras dada la cadena de conexiones e interdependencias (Fig. 2). Los ejemplos biológicos abundan. Una mutación en un gen regulador puede afectar marcadamente la expresión fenotípica de un organismo. A un nivel mayor, la introducción de especies foráneas en un ecosistema es una perturbación que puede afectar las redes tróficas y provocar por un lado la extinción y por otro la sobreabundancia de especies nativas. Uno de los temas más acuciosos de conservación está relacionado con los efectos de cambio climático en altas latitudes. En estas zonas el plancton y las algas dependen de la temperatura baja del mar. Los océanos en altas latitudes han experimentado cambios en el ambiente físico desde hace varias décadas lo que ha reducido las poblaciones de plancton y algas. En la Antártica esta reducción, junto con la sobreexplotación, ha provocado una reducción marcada en las densidades y tamaño corporal promedio de kril (Euphausia superba), una especie clave en la cadena trófica que se alimenta de fitoplancton y algas. El kril es a su vez consumido por peces, pingüinos, otras aves marinas, focas, leones marinos y ballenas entre otros depredadores, cuyas poblaciones también han decaído (Thomson et al., 2000; Reid y Croxall, 2001; Arnold et al., 2006). Así, un efecto localizado sobre un elemento (fitoplancton y algas) se difunde y altera otras especies dada la cadena de interdependencias y conexiones en el ecosistema.
Autoorganización. Esta se define como la tendencia espontánea y constante de los sistemas a generar patrones globales de comportamiento. El ejemplo más clásico es el tipo de reacciones químicas de BelousovZhabotinsky. En estas reacciones se establece un ciclo de oscilación en la concentración de iones debido a procesos de reducción y oxidación sucesivos. Lo anterior genera un patrón repetitivo de cambio en el color de la reacción y la generación de ondas concéntricas. Así, una solución que comúnmente se presenta como un líquido homogéneo puede, bajo ciertas condiciones de composición y concentración de los reactantes, formar patrones espaciales y temporales de organización de manera espontánea (Rossi y Liveri, 2009). En biología hay abundantes ejemplos, el más conocido es el de Dictyostelium discoideum, una ameba que depreda sobre bacterias y habita bajo la hojarasca. En condiciones de baja disponibilidad de nutrientes algunas amebas sintetizan AMP cíclico (cAMP) que se secreta hacia el medio y se difunde. Lo anterior estimula un patrón organizado de movimiento espiralado o en círculos concéntricos de otras amebas en la vecindad en dirección al punto de concentración de cAMP. A medida que estas amebas se mueven también secretan y difunden hacia el entorno cAMP, lo que a su vez aumenta la superficie de concentración de este atrayente químico y estimula aún más amebas a integrarse a las ondas de movimiento. Cuando las amebas son estimuladas por primera vez se tornan refractarias a un nuevo estímulo de cAMP de tal manera que el desplazamiento de las células sucede siempre en una única dirección, hacia el centro, en el que se ubican las amebas que iniciaron la síntesis de cAMP (Steinbock et al., 1993; Pálsson, 2009). Así, las amebas, que de otra forma estarían dispersas de una manera irregular, forman un patrón espacial y temporalmente organizado. Otros ejemplos de autoorganización en biología son las ondas espiraladas de propagación de calcio en Xenopus laevis (Lechleiter et al., 1991), las ondas espiraladas de excitación en músculo cardiaco (Davidenko et al., 1992), los cardúmenes de peces, enjambres de abejas, bandadas de aves, y la distribución de las manadas de elefantes en las sabanas africanas, entre otros.
Dependencia contextual. Los patrones de organización de los sistemas biológicos no dependen exclusivamente de la red de interacciones entre sus partes constitutivas, dependen también de las interacciones que el sistema establece con su entorno. Así, estos sistemas están determinados, constreñidos de alguna manera, por las características de los sistemas que los incluyen, aunque una gran diversidad de los atributos morfológicos, fisiológicos y de comportamiento de los organismos están determinados genéticamente, la manera, o el grado, en el que un gen (o grupo de genes) y su producto se expresan depende de la organización de las redes genéticas en las que el gen se encuentra inmerso (vecindad genética) y del contexto ambiental en el que un organismo se desarrolla. La anemia falciforme, por ejemplo, está asociada a la variabilidad en un solo locus, pero su expresión depende de los alelos en dos loci vecinos; esta es la razón por la cual la enfermedad se presenta en algunas poblaciones y en otras no (Dover et al., 1981; Powars et al., 1989; Hatchwell y Greally, 2007).
Un ejemplo de la dependencia contextual del ambiente es la determinación del sexo en algunas especies de peces sapo, familia Chaunacidae, que viven en las profundidades del océano Pacífico. En estas especies no hay cromosomas sexuales y por tanto no hay diferencias genéticas entre machos y hembras. Después de la eclosión, los alevinos madurarán como hembras si antes de alcanzar un determinado umbral de desarrollo no han entrado en contacto con un pez adulto de su especie. Si por el contrario entran en contacto con un coespecífico adulto, una hembra en tal caso, entonces se adhieren a su epitelio y se desarrollan como machos con hábitos parasíticos. Los ejemplos abundan, pero para no extenderme demasiado, basta con decir que la plétora de rasgos fenotípicos semejantes en especies que han evolucionado de manera independiente, pero bajo condiciones ambientales similares, y que en lenguaje formal se denominan homoplasias, son evidencia indiscutible de la dependencia contextual en los sistemas biológicos.
Dependencia histórica. Los sistemas biológicos son el resultado de un proceso histórico. Todos los atributos fenotípicos de los individuos biológicos se han originado en algún momento de su historia evolutiva. Algunos de ellos representan respuestas adaptativas a condiciones ambientales pasadas; otros son el resultado de restricciones de tipo funcional, estructural o de desarrollo ontogenético.
Las proteínas que regulan funciones vitales básicas, como la replicación de ADN y ARN, la transcripción y la traducción, pueden considerarse homologías primigenias heredadas, con ligeras modificaciones, por todas las formas de vida de los ancestros que habitaron la tierra primitiva hace unos 3.800 m.a. Nosotros, como todos los mamíferos, heredamos del ancestro común de todos los cordados hace unos 525 m.a. el cordón nervioso hueco en posición dorsal, la glándula tiroides, y otras estructuras que aparecen en algún momento de nuestro desarrollo embrionario y que después se pierden, entre ellas el notocordo, las hendiduras faríngeas y la cola postanal. Los huesos de nuestras extremidades los heredamos de un pez crossopterigio hace unos 400 m.a.; el atlas y las vértebras sacras de un ancestro anfibio a mediados del Devónico, hace unos 380 m.a. Las membranas extraembrionarias indispensables para nuestro desarrollo embrionario, el axis y el paladar secundario son parte de la herencia de nuestros ancestros reptialianos durante el Carbonífero. Más recientemente heredamos de un ancestro sinápsido, hace unos 230 m.a., los tres huesecillos del oído medio, la mandíbula compuesta por un único hueso, los glóbulos rojos anucleados, las glándulas mamarias y la lactación. El cuidado parental, el pulgar oponible, la postura bipedal, el lenguaje, el fuego, las letras, las matemáticas, las ciencias y las artes son atributos que adquirimos hace un “segundo” en términos de tiempo geológico. La dependencia histórica es un atributo conspicuo de los sistemas biológicos.
Organización jerárquica y propiedades emergentes. Un sistema complejo está compuesto por partes y a su vez, este mismo sistema, es parte constitutiva de otro nivel jerárquico mayor. Esta serie estructural se extiende hacia arriba y hacia abajo en una secuencia de niveles de organización que va desde los genes, pasando por células, tejidos, órganos, organismos, poblaciones, comunidades y biomas, hasta la tierra misma considerada como un superorganismo. En un nivel cualquiera de esta jerarquía las partes constituyentes interactúan y forman patrones globales de comportamiento que caracterizan el nivel siguiente de organización. Estos patrones globales de comportamiento se definen como propiedades emergentes. Esta es la razón por la cual los intentos de entender un sistema biológico mediante la atomización y descripción de algunas de las partes que lo conforman es una aproximación que aunque necesaria, no es suficiente para lograr este objetivo. Además de la atomización se requiere del estudio de la red de interacciones entre las partes que conforman un determinado sistema y del estudio de la red de interacciones que ese sistema establece con su entorno.
La sociabilidad es un excelente ejemplo de una propiedad emergente a nivel de un grupo de organismos. Esta es un rasgo que evolucionó en algunas especies de abejas y avispas, en todas las especies de hormigas y termitas, y en unas cuantas especies de aves y mamíferos. La sociabilidad solo puede ser atribuida a un grupo de individuos y no de manera independiente a cada uno de los organismos que conforman un grupo social. Las especies sociales exhiben rasgos que resultan de la interacción entre organismos, es el caso de las estrategias cooperativas de forrajeo, de defensa, de cuidado y protección de individuos juveniles y adultos discapacitados, de aprendizaje y enseñanza, entre otros (Muñoz Durán, 2009).
Irreductibilidad ontológica y epistemológica. Se puede decir que del efecto combinado de las anteriores propiedades de los sistemas biológicos se derivan dos de sus características fundamentales: primero, la esencia de los sistemas biológicos no reside en las partes que los conforman –genes, células, organismos, o poblaciones–, reside en la red de interacciones entre éstas partes y en las interacciones que el sistema establece con su entorno. Segundo, los sistemas biológicos son irreducibles desde el punto de vista ontológico y, por tanto, epistemológico.
Cabe entonces preguntarse hasta qué punto los conceptos, hipótesis y métodos utilizados comúnmente en teoría evolutiva tienen en cuenta las anteriores características de los sistemas biológicos para explicar su cambio a través del tiempo.
EVOLUCIÓN Y GENOCENTRISMO
El mensaje principal de la teórica evolutiva clásica es sencillo: el proceso evolutivo es estimulado por mutaciones aleatorias que alteran la información genética, lo que a su vez genera variabilidad fenotípica a nivel de los organismos. Los organismos con rasgos que les confieren una mayor probabilidad de sobrevivencia y reproducción, en el contexto de competición por recursos limitados, dejarán mayor descendencia y, de esta manera, sus genes experimentarán un aumento en la frecuencia relativa en generaciones siguientes. En el contexto anterior, la selección natural se define como la perpetuación –sobrevivencia y reproducción– diferencial de organismos coespecíficos que difieren en uno o más rasgos fenotípicos.
Desde la publicación en 1859 del Origen de la Especies se acepta que los organismos son por excelencia las unidades de selección. Sin embargo, el desarrollo de la teoría sintética de la Evolución y la genética de poblaciones durante la década de los años 30 del siglo pasado, desplazó la atención hacia el papel que el cambio en las frecuencias génicas tiene para explicar el fenómeno evolutivo. El desarrollo posterior de la biología molecular y la profusión de modelos matemáticos, estadísticos y de computador con cierto poder explicativo y capacidad de predicción ha enfatizado la idea de genes como unidades de selección.
Esta tendencia genocentrista se ha reforzado con afirmaciones que resumen la interpretación que del mundo tienen prestigiosos biólogos evolutivos. La primera referencia histórica de la que tengo conocimiento se remonta a Theodosius Dobzhansky, el mismo que en 1973 acuñó la famosa frase “Nothing in biology makes sense except in the light of evolution” (Dobzhansky, 1973). Varias décadas antes, en 1937, Dobzhansky también afirmó: “Since evolution is a change in the genetic composition of populations, the mechanisms of evolution constitute problems of population genetics” (Dobzhansky, 1937). La obra de investigadores como Richard Dawkins ha vigorizado aún más las aproximaciones reduccionistas en evolución. En la página XVI de su libro “River out of Eden: A Darwinian view of life” (Dawkins, 1995) aparece una figura que sintetiza el mensaje principal de la obra de Dawkins: la vida, desde el origen hasta el presente, es un flujo de nucleótidos de ADN y ARN; el resto de entidades biológicas, particularmente los organismos, tan solo son vehículos que aseguran la persistencia y el paso de generación a generación de los genes que confieren mayores ventajas en la interacción con el entorno. Esta visión genocéntrica del mundo sigue siendo común en las nuevas generaciones de biólogos moleculares. Es el caso de Sean B. Carroll, quien en uno de sus libros más recientes “The making of the fittest: DNA and the ultimate forensic record of evolution” (Carroll, 2006) utiliza las siguientes frases: “Each step in evolution, we now know, is taken and recorded in DNA” (Cada evento evolutivo sucede y se registra en las secuencias de ADN; p. 33); más adelante en la misma página escribe: “The unique DNA sequence of each species is a complete record of the present. It is an inventory of all the genes used to build and operate that creature” (Las secuencias únicas de ADN en una especie constituyen un registro completo del presente. Son un inventario de todos los genes usados para construir y operar esa criatura; p. 33). En la página 57 asevera “The source of all variety is mutation” (La fuente de toda variación es la mutación).
Durante la mayor parte de la historia de la teoría evolutiva nos hemos aproximado al estudio de la evolución desde un punto de vista simple y reduccionista. Esta tradición pareciera tener como guía el paradigma según el cual la vida es una propiedad que reside en los genes. Quizás esta tendencia explicativa esté incentivada precisamente por la dificultad de abordar, desde el punto de vista conceptual y metodológico, algunas de las características mencionadas de los sistemas biológicos, entre ellas: el alto número de partes constitutivas, la densa red de interacciones y bucles de retroalimentación, su dependencia contextual e histórica, y su organización jerárquica. Aunque es mucho lo que hemos avanzado y aprendido del estudio de los genes, y aunque son muchas las aplicaciones prácticas que hemos derivado, y a sabiendas de que aún nos queda mucho por descubrir a este nivel, es necesario extender el marco conceptual de la teoría evolutiva más allá de la aproximación genocentrista para reconocer la complejidad de los entes que evolucionan. La hipótesis de selección jerárquica es una alternativa adecuada, ya que como veremos en la siguiente sección, considera las propiedades descritas de los sistemas complejos y de ella se derivan implicaciones que enriquecen nuestra percepción de la evolución.
SELECCIÓN JERÁRQUICA: PROPIEDADES EMERGENTES, EFECTOS ASIMÉTRICOS E INDIVIDUOS BIOLÓGICOS
Una alternativa teórica para explicar la evolución es la selección jerárquica. Según esta propuesta la selección natural puede actuar de manera simultánea en varios niveles de organización biológica. Esta idea resulta de la combinación de dos propuestas tempranas en la historia de la teoría evolutiva. La primera es la intuición del propio Darwin respecto de la necesidad conceptual de la selección a nivel de especies para explicar el proceso de diversificación. De esta manera se extienden las unidades de selección para incluir entidades biológicas por encima del nivel de los organismos. En varias secciones del Origen de las Especies, Darwin hace una analogía con el proceso de competencia entre organismos y menciona que la competición a nivel de especies puede conllevar a la extinción de las especies menos dotadas y a la sobrevivencia de las especies más hábiles. Darwin enfatiza que la competencia puede ser más intensa entre especies que derivan de un mismo ancestro y que por tanto son más semejantes desde el punto de vista fenotípico y ecológico. Así Darwin insinúa que la selección puede operar en instancias que reúnan todos los requisitos indispensables para su acción: variación heredable, y relación entre variación y éxito relativo.
La segunda idea que amplió el espectro de acción de la selección natural para trabajar sobre unidades a nivel infraorganísmisco fue el mecanismo de “selección germinal” propuesto a finales del siglo XIX por August Weismann, uno de los biólogos teóricos más importantes de la historia de la biología. Entre los temas sobre los que reflexionó Weismann está el mecanismo de la herencia. Según Weismann las unidades de la herencia son los “bióforos” y un tipo particular de estos que llamó los “determinantes” (Rostand, 1985). Según su línea de argumentación, los bióforos se agrupan para formar los cromosomas1, los que a su vez residen y permanecen aislados de las condiciones del entorno en el núcleo de las células de la línea germinativa. De esta manera Weismann termina con la dependencia de la teoría darwiniana sobre el esquema de herencia propuesto por Lamarck: la herencia de caracteres adquiridos, bien por el efecto del uso y el desuso o por el efecto de las condiciones ambientales. Esta ruptura conceptual caracteriza lo que hoy conocemos como neodarwinismo (Rostand, 1985; Gould, 2002). Weismann también elabora el argumento del “Allmacht selection”, que se podría traducir como la “selección todopoderosa” u “omnipotente”, y desarrolla la teoría de la recombinación sexual como mecanismo que genera variabilidad a nivel de las partículas de la herencia (Mayr, 1982; Rostand, 1985; Gould, 2002). Es precisamente en el contexto de estas dos últimas ideas que Weismann propone la “selección germinal”, es decir la que se establece entre los bióforos de tal manera que los más aptos se imponen sobre los más débiles. Esta es la idea base que el biólogo alemán utiliza para explicar la degeneración y eventual eliminación de estructuras previamente adaptativas. La teoría de la “selección germinal” fue abandonada por el efecto combinado de varios hitos en la historia de la biología. Primero, el redescubrimiento de las leyes de la herencia de Mendel en 1900 de manera independiente, y casi simultánea, por los botánicos Hugo De Vries, Carl Correns y Erich Tschermark. Segundo, el descubrimiento del fenómeno de la mutación por Hugo de Vries a inicios del siglo XX. Las mutaciones a gran escala descritas por De Vries no solo ponen de manifiesto una nueva fuente de variación en adición a la recombinación sexual propuesta por Weismann, sino que también promovió el desarrollo de la teoría saltacionista para explicar las discontinuidades fenotípicas que se observan en algunos grupos taxonómicos. Además de lo anterior, la noción de “bióforos” de Weismann fue desestimada debido a la confirmación por parte de Thomas Hunt Morgan, durante la segunda década del siglo pasado, de la existencia de las unidades de la herencia mendeliana, o genes como los llamó Wilhelm Johansen en 1909, en los cromosomas de la drosófila (Rostand, 1985).
Por otro lado, la intuición de Darwin respecto de la selección a nivel de especies, y en un sentido más general la selección de grupos de organismos, fue también duramente criticada y abandonada casi por completo después de que el matemático, estadístico y genetista Ronald A. Fisher argumentara durante la tercera década del siglo XX que aunque la selección a nivel de grupos es teóricamente posible, este proceso es altamente improbable y además difícil de observar y cuantificar. Fisher fundamentó su crítica en la relación que existe entre la potencia de la acción de la selección natural, el número de individuos sobre la cual opera, la disponibilidad de variabilidad y la velocidad con que ésta se genera (Fisher, 1930). Fisher hizo notar que a medida que ascendemos en la jerarquía biológica desde genes, pasando por organismos, grupos sociales, y poblaciones hasta llegar al nivel de especies, hay una disminución en el número de entidades presentes en cada nivel (con la excepción de que en ocasiones el número de organismos en una especie puede ser mayor que el número de genes que la conforman). Así, a medida que ascendemos en la jerarquía biológica, la potencia de la selección natural para guiar el cambio evolutivo disminuye y, de manera simultánea, aumenta el efecto de los procesos estocásticos. A medida que pasamos de niveles bajos a niveles más altos, también disminuye la variabilidad, que es la materia prima del proceso de selección, lo que a su vez reduce la efectividad de la selección, por lo menos la de tipo disruptiva y direccional, no necesariamente la de tipo estabilizadora. Por último, en niveles supraorganísmicos los tiempos generacionales son más largos, por tanto la tasa de generación de variabilidad es más baja y esto reduce aún más las oportunidades para la acción del proceso selectivo.
A pesar de la sensatez y aceptación de las críticas de Fisher a los intentos de extender la acción de la selección natural para trabajar sobre entidades biológicas por encima del nivel de los organismos, existen varios argumentos, algunos de ellos descuidados durante la discusión inicial y otros de desarrollo reciente, que le dan un nuevo impulso a la hipótesis de selección jerárquica. Además de argumentos de estilo teórico también existe evidencia de selección a nivel de especies basada en la comparación del éxito evolutivo de linajes que varían respecto de propiedades emergentes. Es el caso de linajes de carnívoros sociales con tasas de extinción significativamente más altas que las medidas en linajes que viven en solitario o en pequeños grupos familiares (Muñoz Durán, 2002; Muñoz Durán, 2009).
El primer argumento a favor de la selección jerárquica se refiere a la asimetría de efectos que la alteración de las propiedades emergentes en un nivel particular tiene sobre el arreglo jerárquico (Valentine y May, 1996). En otras palabras, cambios en niveles superiores afectan la organización y funcionamiento de todos los niveles jerárquicos inferiores. En contraste, cambios en niveles inferiores de la jerarquía (i.e. alteraciones en la secuencia de nucleótidos) no ejercen, en la mayoría de los casos, un efecto marcado sobre la estructura, la organización o el desarrollo de procesos en niveles superiores. Un ejemplo puede ser el de un grupo social en el que los organismos que lo conforman cooperan de manera efectiva en la búsqueda y defensa de recursos, en el cuidado de juveniles y adultos discapacitados, y en general en todas las actividades de la vida cotidiana. Estos grupos tendrán ventajas ecológicas y de competición respecto de otros grupos sociales en los cuales hay falencias de cooperación. Con el tiempo, los grupos de cooperadores efectivos tendrán mayores probabilidades de éxito ecológico y de formación de nuevos grupos derivados. Así mismo, los organismos que lo integran tendrán mayor probabilidad de sobrevivencia y reproducción y, a largo plazo, los genes presentes en estos grupos de cooperadores efectivos incrementarán su frecuencia en la población. Sin embargo, se debe reconocer que en ocasiones las alteraciones en niveles inferiores se expresan en los superiores. Es el caso por ejemplo de mutaciones en genes reguladores que suelen tener efectos fenotípicos significativos y pueden incluso conducir al origen de innovaciones evolutivas que caracterizan grupos taxonómicos completos.
El segundo argumento se refiere a la posibilidad de extender las unidades de selección para incluir diversas entidades biológicas, de naturaleza taxonómica o ecológica, siempre y cuando se cumpla con los requisitos para la acción de la selección: variación en las propiedades que caracterizan dichas entidades, las distintas variantes deben tener efectos diferentes sobre la eficacia biológica de las entidades que las portan y, por último, esta variación debe ser heredada de alguna manera. Este argumento sencillo y a la vez contundente fue propuesto por Lewontin en 1970 y representa el primer intento acertado2, del que tengo conocimiento, de retomar la discusión sobre las unidades de selección. Lewontin además argumentó que las dos primeras condiciones son generales y que la herencia puede proceder de manera mendeliana, citoplasmática o cultural entre otras formas posibles (para una revisión actualizada sobre mecanismos de herencia epigenética ver Jablonka y Lamb, 2006). Para la época, la discusión a favor o en contra de la selección en niveles múltiples giraba más en torno a argumentos de estilo teórico que sobre el análisis de la evidencia (Lewontin, 1970). Esta misma línea de argumentación fue retomada algunos años después por varios autores, entre ellos Arnold y Fristrup quienes incluso publicaron un artículo titulado “Teoría de la evolución por selección natural: una expansión jerárquica”. En este artículo los autores argumentan que “alelos, individuos, y especies son todos ejemplos de entidades que expresan variación en las propiedades en la que se basa la selección natural: diversificación (reproducción), persistencia (sobrevivencia) y herencia de caracteres” (Arnold y Fistrup, 1982).
El tercer argumento a favor de la selección jerárquica se esboza dos años después de la defensa de Lewontin y corresponde a una de las principales implicaciones del patrón macroevolutivo de equilibrio puntuado propuesto por Eldredge y Gould en 1972: la competencia y selección a nivel de especies. Estas implicaciones fueron puestas de manifiesto en trabajos posteriores, particularmente por Elisabeth Vrba, en los que se aclararon detalles del proceso de selección entre especies y se enfatizó que este proceso depende de la acción simultánea de la selección a nivel de organismos y grupos de organismos (Eldredge y Gould, 1972; van Valen, 1973; Stanley, 1975;vanValen, 1975; Gould, 1982; Vrba, 1984a, b; Vrba y Eldredge, 1984; Vrba, 1985; Vrba y Gould, 1986). Incluso se aclaró que en ocasiones estos dos procesos de selección pueden actuar de manera antagónica; es decir, en algunos casos los grupos de organismos pueden ser seleccionados a pesar de las implicaciones negativas que este proceso pueda tener para los organismos que conforman el grupo y viceversa. En esta época se postularon varios modelos de selección actuante sobre características emergentes por encima del nivel de los organismos, entre ellos los de selección dependiente del contexto, selección de grupo y selección de multiniveles. Estos modelos fueron discutidos desde el punto de vista teórico, mientras que la proposición de hipótesis concretas, la colección de evidencia a favor o en contra de las mismas, el desarrollo de métodos de colección y análisis de datos, el desarrollo de modelos de herencia epigenética y la cuantificación de esta herencia fueron temas que no recibieron la suficiente atención y en los que aún hay mucho por hacer.
El último argumento en favor de la selección jerárquica que me parece importante mencionar fué el desarrollo del concepto de individuo biológico. De la discusión planteada durante la década de los ochenta se puede inferir que un individuo biológico se caracteriza por tener un origen y final discretos, localización espacial, propiedades emergentes, alguna forma de reproducción sexual o asexual, y por interactuar con su entorno como un todo coherente. Esta caracterización permitió reafirmar algunas entidades biológicas previamente propuestas como unidades de selección e incluir algunas nuevas. Así mismo permitió establecer por lo menos dos jerarquías importantes desde el punto de vista evolutivo, las entidades incluidas en cada una de estas jerarquías varía según el autor (Arnold y Fistrup, 1982; Gould, 1982; Eldredge y Salthe, 1984; Vrba y Eldredge, 1984; Eldredge, 1985; Levinton, 1988). La primera es de tipo genealógico y se extiende desde los genes, pasando por los organismos, y especies, hasta los grupos monofiléticos. La segunda jerarquía es la ecológica que se extiende desde los genes, los organismos, grupos familiares, grupos sociales, especies eusociales, y poblaciones hasta culminar en las comunidades ecológicas.
Entre estas dos jerarquías, la ecológica expresa de manera más clara varias de las propiedades de los sistemas complejos. Entre ellas, que cada nivel está compuesto por numerosas partes que interactúan, la existencia de interdependencias entre los individuos que componen cada nivel y entre individuos en niveles consecutivos de la jerarquía, la autoorganización, y la dependencia histórica y contextual.
Sin embargo, desde el punto de vista de la selección jerárquica, las dos jerarquías presentan limitaciones significativas. En primera instancia, para algunos de los niveles propuestos es difícil argumentar la manera en que se expresan ciertas propiedades de los individuos biológicos. Por ejemplo, no es claro cómo las comunidades ecológicas, y otros niveles más inclusivos de la jerarquía ecológica, manifiestan las propiedades de origen y final discretos, reproducción, herencia, e interacción como un todo coherente con su entorno. De otro lado, los grupos monofiléticos de la jerarquía genealógica solamente se pueden describir en términos de propiedades agregativas y no con base en propiedades emergentes propiamente dichas. Las propiedades agregativas de un sistema biológico representan resúmenes estadísticos de las partes que las constituyen.
Lo anterior me lleva a argumentar que existen solo tres unidades de selección: sistemas de secuencias moleculares autoreplicantes (genes en un sentido amplio y como las seguiré llamando en lo que resta de este artículo), organismos y grupos de organismos que cooperan. Así mismo, que la principal implicación que se puede derivar de la hipótesis de selección jerárquica es que el mecanismo de evolución evoluciona a medida que surgen innovaciones y se conforman nuevos niveles de organización.
INNOVACIONES EVOLUTIVAS Y CONFORMACIÓN DE NIVELES DE SELECCIÓN
En el transcurso de la evolución han aparecido tres niveles en los cuales opera la selección. Estos niveles han surgido de manera secuencial y en ellos se codifica y transfiere información necesaria para la sobrevivencia y multiplicación de los individuos biológicos propios del nuevo nivel que se origina, así como para la de los individuos en niveles de organización antecedentes estructural y temporalmente.
Los individuos biológicos que evolucionaron primero y sobre los cuales opera la selección natural son los sistemas de secuencias moleculares autoreplicantes de ARN. Este es el más antiguo, podríamos decir ancestral, de los niveles de selección. Ensayos de laboratorio han permitido imaginar un escenario probable de origen de la vida hace unos cuatro mil millones de años que resultó en individuos con metabolismo extremadamente simple, rodeados por una membrana, con la capacidad de replicarse, y compuestos por cadenas cortas de ARN y proteínas que catalizaban su síntesis de manera recíproca. En una fase posterior, se pudieron adicionar cadenas de ADN.
Debido al estado precoz de los mecanismos de control de la replicación es fácil imaginar que en las etapas tempranas, partiendo desde los primeros polímeros con capacidad de autorreplicación, se observan los requisitos indispensables para la acción de la selección natural: variación, capacidad diferencial de replicación y herencia. La selección pudo entonces trabajar a nivel de las secuencias de ARN, ADN y las proteínas que estas codificaban para mantener funciones vitales y fundamentales comunes a todas las formas de vida como son la replicación de ARN y ADN, la transcripción de una cadena de ADN a ARN mensajero y la traducción a una secuencia de aminoácidos para conformar una proteína. Las secuencias de nucleótidos y aminoácidos que demostraron ser más eficientes para mantener este metabolismo sencillo han sido sometidas durante cerca de cuatro mil millones de años a un proceso de selección estabilizadora y las podemos encontrar hoy, con ligeras modificaciones, en todas las formas de vida: bacterias, arqueobacterias y eucariotes (protistos, plantas, hongos y animales).
Aunque el proceso de selección a nivel de genes debió ser muy agitado en la tierra primitiva, particularmente cuando aparecieron las primeras entidades basadas en ARN, este proceso dejó de operar, se extinguió, cuando aparecieron individuos biológicos más complejos, compuestos por un mayor número y diversidad de secuencias de ARN, ADN y proteínas, y con metabolismos más sofisticados. Estos individuos unicelulares (bacterias y arqueobacterias) son los primeros representantes del segundo nivel de selección: el organísmico. Estos individuos fueron y siguen siendo seleccionados no por la posesión de una forma particular de un gen, sino por las nuevas propiedades que emergen de la interacción entre las diversas secuencias que los conformaban. Los organismos representantes de estos dos reinos están conformados por miles de genes y proteínas que cooperan para el funcionamiento y reproducción. En otras palabras, estos organismos debieron ser seleccionados por su habilidad para interactuar como un todo coherente con su entorno. El proceso de selección a nivel de organismos es sin duda la forma prevalente de selección en términos históricos y taxonómicos. Este nivel de selección opera desde hace unos 3.800 m.a. y ha acompañando toda la historia de diversificación de las bacterias, arqueobacterias y posteriormente de los eucariotas.
Un tercer nivel de selección se establece cuando aparece por primera vez el cuidado parental acompañado de estrategias de aprendizaje y enseñanza. Este es el tercer y más reciente nivel de selección: el de los organismos que cooperan. El cuidado parental ha evolucionado de manera independiente en varios grupos taxonómicos. En el caso particular de los vertebrados se observan una gran diversidad de especies que despliegan algún tipo de cuidado parental. En la mayoría de las ocasiones el cuidado parental se reduce a la protección de los huevos fecundados y, en algunas ocasiones, a la protección y alimentación de los neonatos. Sin embargo, en algunos linajes de reptiles, aves y mamíferos este cuidado se extiende para incluir relaciones de enseñanza y aprendizaje mediante las cuales se transfiere información necesaria para la sobrevivencia.
En la mayoría de estas especies la relación más duradera entre organismos es la que se establece entre la madre y sus crías. El cuidado maternal es la forma más sencilla y antigua en la que se expresa el cuidado parental. En este sentido podríamos decir que las madres son una de las mejores invenciones de la naturaleza. En otras especies, particularmente aves y mamíferos, el cuidado parental puede incluir la actividad de los dos padres. En un número relativamente bajo de especies también se observa cuidado aloparental ejercido por hermanos de camadas previas que permanecen por periodos extendidos con los padres. Incluso, hay especies que viven en grupos sociales más complejos, en los que individuos no emparentados pueden cooperar con el cuidado y entrenamiento de neonatos yjuveniles. El cuidado parental y aloparental involucra la transmisión de información a las crías sobre el tipo de recursos que pueden ser utilizados, dónde buscarlos, cómo obtenerlos, qué especies son competidoras o depredadoras, cómo evadirlas o enfrentarlas, entre otra información que afecta las probabilidades de sobrevivencia y reproducción de las nuevas generaciones.
Basado en lo anterior propongo la noción de socioma. Como argumentaré a continuación este es un concepto complementario al de genoma y es central para entender la acción de la selección natural a nivel de los grupos de organismos que cooperan.
SOCIOMA
Por “socioma” me refiero a la información que no se codifica genéticamente y que afecta las probabilidades de sobrevivencia y reproducción de los organismos y los grupos a los cuales estos pertenecen. El socioma se expresa en las interacciones sociales y ecológicas, y se codifica, almacena y transmite mediante sistemas de comunicación (vocales, olfativos, posturales, entre otras), a manera de tradiciones, formas de uso, artefactos, y alteraciones al entorno, entre otras. El socioma engloba las propiedades emergentes a nivel de grupo y se hereda de manera epigenética. El socioma varía entre grupos que pertenecen a la misma especie y también entre grupos de especies diferentes, estas variaciones afectan de manera diferencial el desempeño ecológico y el éxito evolutivo de los grupos. Una de las razones para esta variación es que el socioma es histórico y contextodependiente, por tanto es afectado por los eventos y condiciones ambientales particulares en las que se desenvuelve un grupo. El socioma puede variar de manera aleatoria, de forma análoga a las mutaciones que ocurren en el genoma; sin embargo, aunque algunas variaciones del socioma lucen erráticas en sus primeros estadios de desarrollo, estas están dirigidas y resultan de la necesidad de resolver problemas específicos. De esta manera, algunos grupos de organismos tienen la capacidad de modificar su comportamiento cuando las condiciones ambientales cambian y pasan estas modificaciones mediante interacciones de enseñanza y aprendizaje a la siguiente generación. Es así como la evolución del socioma expresa varios atributos lamarckianos, entre ellos la herencia de caracteres adquiridos y el efecto del uso y el desuso. Más aún, la interacción entre el socioma y el genoma abre una nueva dimensión al proceso evolutivo y se convierte en un factor decisivo en la evolución de diversos grupos taxonómicos.
LA EVOLUCIÓN DEL MECANISMO DE SELECCIÓN Y EL BUCLE DE RETROALIMENTACIÓN GENOMASOCIOMA
Una de las implicaciones teóricas más potentes de la hipótesis de selecciónjerárquica es la existencia de un bucle de retroalimentación, o mutua afectación, entre el genoma y el socioma. Las variaciones en el socioma afectan las interacciones del entorno social y ecológico. La acumulación de estas alteraciones a lo largo de varias generaciones tiene como efecto la relajación, el refuerzo o la generación de nuevas presiones de selección. Así, las modificaciones del socioma mantenidas a lo largo de múltiples generaciones pueden afectar el genoma y provocar cambios en las frecuencias génicas que alteran el fenotipo de los organismos y conllevan a cambios en su morfología, fisiología y posiblemente su comportamiento. Los organismos modificados a su vez afectan la red de interacciones al interior de los grupos, y entre estos y su entorno, lo que modifica el régimen de selección a nivel de grupos y provoca con el tiempo nuevas modificaciones al socioma. Así se establece un nuevo ciclo de mutua afectación entre el socioma y el genoma. Este proceso podría esquematizarse como una espiral de retroalimentación que se extiende en el tiempo.
LAS MODIFICACIONES DEL SOCIOMA PUEDEN GENERAR NUEVAS PRESIONES DE SELECCIÓN A NIVEL DEL GENOMA
Un ejemplo es el cambio de hábitos culturales y evolución de la resistencia a la lactosa en humanos. La leche es un alimento rico en carbohidratos, grasas, proteínas, calcio y agua, esta última es de particular importancia en regiones áridas. Sin embargo, la habilidad de digerir lactosa declina en la mayoría de los humanos rápidamente después del destete. Entre las consecuencias de la intolerancia a la lactosa está la diarrea con la consecuente pérdida de agua y una reducción en la disponibilidad de nutrientes debido a la incapacidad de utilizar el recurso leche. Estudios recientes (Tishkoff et al., 2006; Voight et al., 2006) indican que la domesticación de bóvidos y consumo de leche por adultos ocurrió de manera independiente entre 8.000 y 9.000 años en poblaciones del norte de Europa y hace unos 7.000 años en África. Este proceso de domesticación de bóvidos y el aprovechamiento controlado de nuevos recursos representa una innovación a nivel del socioma que trae consigo beneficios, pero que de igual manera impone nuevas presiones de selección. De esto se puede inferir que en poblaciones pastoriles antiguas, especialmente las que habitaron en zonas áridas, los individuos adultos con la capacidad de digerir lactosa tendrían una ventaja respecto de aquellos con intolerancia. Esto explica los niveles comparativamente altos de resistencia a la lactosa que se observan en las poblaciones actuales que descienden directamente de las que domesticaron los bóvidos (Tishkoff et al., 2006). También es importante resaltar que este rasgo es controlado genéticamente (gen LCD) y que evolucionó de manera independiente en poblaciones del norte de Europa y África como resultado de cambios en los hábitos culturales. Como sabemos, los eventos de evolución convergente son uno de los mejores indicadores de procesos adaptativos. Este es un ejemplo claro de la manera en que modificaciones a nivel del socioma imponen nuevos contextos de selección de rasgos controlados a nivel del genoma.
LAS ALTERACIONES DEL SOCIOMA PUEDEN CONDUCIR A LA RELAJACIÓN DE PRESIONES SELECTIVAS A NIVEL DEL GENOMA
Un ejemplo puede ser el invento de herramientas para macerar, lo que constituye claramente una innovación a nivel del socioma, y la disminución de la ventaja mecánica de la musculatura mandibular en humanos, un rasgo controlado genéticamente. Nuestros parientes evolutivos más cercanos, chimpancés, gorilas, orangutanes, macacos y algunos ancestros de nuestra especie, entre ellos los géneros Australopithecus y Paranthropus tienen músculos temporales, mandíbulas inferiores, procesos coronoides, fosas temporales, crestas sagitales y dientes caninos marcadamente más desarrollados que los observados en todas las especies del género Homo, incluida la nuestra. Todo esto hace que la presión ejercida durante la mordida sea mayor en estas especies que en la nuestra. También existe una relación inversa entre la ventaja mecánica de la anatomía cráneo dental y el grado de encefalización. En otras palabras, debido a restricciones de tipo estructural las adaptaciones que confieren mayor fuerza de mordida conllevan a una restricción el volumen de la caja craneal y por tanto una menor capacidad cerebral. El gen MYH16 (Myocine Heavy Chain) codifica la miocina de las fibras musculares craneales (Stedman et al., 2004), en particular en los músculos temporales que se originan en la cresta sagital y se insertan en el proceso coronoides de la mandíbula inferior. Este músculo tiene como función la aducción de la mandíbula. De tal manera que mientras más desarrollado el músculo temporal más fuerza se genera durante el cierre de la boca y por tanto mayor presión durante la mordida, lo que es importante para triturar alimentos o durante encuentros agonísticos intra o interespecíficos.
La causa próxima de un menor grado de desarrollo del músculo temporal en humanos comparado con nuestros parientes evolutivos más cercanos reside en la pérdida de dos bases en la secuencia del gen MYH16 en nuestros ancestros que altera el marco de lectura para la síntesis de miocina (Stedman et al., 2004). Esto altera la expresión del gen, de tal manera que las fibras de nuestro músculo temporal son marcadamente más cortas, por lo tanto los músculos son menos desarrollados que en nuestros parientes cercanos. Se calcula que la mutación que provocó esta alteración del músculo temporal ocurrió alrededor de 2.4 m.a. (Stedman et al., 2004). Sin embargo, la evidencia fósil indica que la reducción en la potencia de mordida en nuestro linaje aparece por primera vez en Homo ergaster, cuyos registros más antiguos datan de 2.0 m.a. (Wood y Lonergan, 2008). Es interesante notar que las primeras herramientas de piedra que fueron utilizadas para triturar alimentos duros como semillas y huesos tibiales de ungulados datan de hace 2.5 m.a. y se encontraron asociadas a restos de Australopithecus garhi (Asfaw et al., 1999; McHenry y Coffing, 2000; Ambrose, 2001; Boisserie et al., 2008). La función de las primeras herramientas de piedra, partir y macerar alimentos duros, es análoga a la ejercida por el músculo temporal en combinación con las mandíbulas y la dentición. Así, la secuencia de cambios evolutivos inicia con la invención de herramientas de piedra para partir y macerar alimentos duros, este evento precedió en por lo menos cien mil años (0.9 m.a.) la mutación genética que resultó en músculos temporales con menor eficiencia mecánica y medio millón de años la edad más antigua de fósiles homínidos que muestran de manera clara una reducción en la fuerza de masticación.
La interpretación del anterior escenario evolutivo es sencilla: una innovación a nivel del socioma relajó presiones selectivas sobre el grado de desarrollo del músculo temporal, un rasgo bajo control genético, lo que permitió que se acumularan mutaciones y la evolución de músculos temporales con ventaja mecánica reducida. Eso a su vez permitió modificaciones en otras áreas de la morfología craneal relacionadas con la función de masticación, como lo son el menor grado de desarrollo del proceso coronoides del dentario y, muy particularmente, de la cresta sagital. Esto a su vez pudo reducir restricciones de tipo estructural que favorecieron un mayor desarrollo de la caja craneal, lo que a su vez permite un crecimiento mayor del cerebro y el desarrollo de nuevas capacidades como el lenguaje y el pensamiento simbólico. Estas modificaciones morfológicas mediadas por el genoma, y sus nuevas potencialidades, a su vez estimulan e impulsan nuevas y numerosas modificaciones a nivel del socioma, entre ellas el desarrollo de nuevas herramientas, la tecnología, las artes y las ciencias. Esta espiral de mutua afectación entre el genoma y el socioma aún continúa jalonando la evolución de nuestro linaje y, con seguridad, la de otros linajes en los que los organismos se agrupan para cooperar.
LAS MODIFICACIONES DEL SOCIOMA PUEDEN REFORZAR PRESIONES DE SELECCIÓN A NIVEL DEL GENOMA
Este tipo de mecanismos puede explicar por qué en humanos hay en promedio tres veces más copias (hasta 14) del gen de la amilasa salivaría (AMY1) que en chimpancés (dos copias). Nuestro linaje tiene una dieta más diversa que la de los chimpancés y de ella hacen parte una gran diversidad de semillas, raíces y tubérculos, algunos de ellos ricos en almidones. Estas substancias son una fuente rica en calorías y nutrientes, algunos particularmente importantes para prevenir la diarrea, una causa importante de muerte en poblaciones humanas. Así, un aumento en la capacidad de digerir y metabolizar almidones conlleva beneficios de tipo nutricional. Lo anterior se puede conseguir mediante un aumento en la síntesis de amilasa, lo que a su vez resulta de un aumento en el número de copias del gen AMY1. De acuerdo a lo anterior es posible imaginar un escenario en el cual hace unos 6.5 m.a, después de la divergencia del ancestro común con los chimpancés, innovaciones a nivel del socioma en nuestro linaje representadas por cambios hacía una dieta más diversa en productos vegetales ricos en almidón (Laden y Wrangham, 2005), pudieron resultar en selección direccional a favor de fenotipos con un mayor número de copias del gen AMY1 y, por tanto, mayor capacidad de síntesis de amilasa. Este proceso pudo ser reenforzado por una innovación más reciente en nuestro socioma: el descubrimiento hace unos 10.000 años de la agricultura y un aumento en el consumo de granos domesticados.
Este mismo tipo de mecanismos –innovaciones en el socioma que refuerzan presiones de selección positiva– puede explicar la diferencia en el número de copias del gen AMY1 entre varias poblaciones humanas actuales cuyas dietas difieren en su contenido de almidones (Coyne y Hoekstra, 2007; Perry et al., 2007). Poblaciones humanas actuales que derivan de ancestros con una dependencia marcada de la agricultura y cuya dieta incluye altos contenidos de almidones (Japón, Americanos de origen Europeo e indígeneas Hadza) tienen un número diploide alto (Promedio=6,7, s.d.=2,35, N=133) de copias del gen AMY1 (Perry et al., 2007). En contraste, poblaciones humanas actuales con tradicón de cazadoresrecolectores (Biaki y Matubit), con hábitos marcadamente pastoriles (Datog), o con hábitos pastoriles y pescadores combinados (Yakut), cuya dieta presenta menos contenido de almidones, tienen un número diploide menor (Promedio=5,4, s.d.=2,04, N=93) de copias de este gen (Perry et al., 2007).
CONCLUSIONES
De las características de los sistemas complejos podemos inferir que los individuos biológicos son irreducibles desde el punto de vista ontológico y, por tanto, epistemológico. Por tanto se requiere una expansión formal de la teoría evolutiva que refleje los atributos de los sistemas cuyo cambio en el tiempo pretende explicar. La hipótesis de selecciónjerárquica, que implica la acción de la selección de manera simultánea sobre entidades biológicas en diferentes niveles de organización, es una alternativa apropiada ya que refleja algunos de los atributos más relevantes de los sistemas biológicos. En particular, esta hipótesis es consecuente con la historia de aparición de innovaciones evolutivas, algunas de las cuales caracterizan nuevos niveles jerárquicos con significado desde el punto de vista de la operación de la selección natural. Desde de este punto de vista podemos decir que el proceso evolutivo también evoluciona. En la historia de la vida han existido por lo menos tres unidades de selección: genes, organismos y grupos de organismos que cooperan. Las poblaciones de genes corresponden al nivel más ancestral sobre el cual operó la selección. Muy probablemente este nivel de selección dejó de ser operante cuando aparecieron los primeros organismos compuestos por múltiples genes y metabolismo complejo. Los niveles de selección restantes, poblaciones de organismos y poblaciones conformadas por grupos de organismos que cooperan contienen información indispensable para el proceso evolutivo contenida en el genoma y el socioma respectivamente. Entre el genoma y el socioma se establece un bucle de retroalimentación, de tal manera que las modificaciones en cualquiera de estos niveles estimula modificaciones en el otro dando lugar a una espiral de mutuas afectaciones que se extiende en el tiempo.
1 Descritos en detalle por el botánico Strasburger en 1880 y bautizados con este nombre por Waldeyer en 1888 (Rostand, 1985).
2 Con el ánimo de recapitular de mejor manera la historia de la discusión sobre la selección grupal, debo mencionar que quién primero retomó esta discusión, aunque con base en un argumento equivocado, fue WynneEdwards en 1963. Este autor argumentó la necesidad de la selección intergrupal para explicar la evolución de grupos sociales (WynneEdwards, 1963). Aunque su intuición es justificada, el centro de su propuesta contiene un error: la competición entre grupos según este autor se basa en diferencias de comportamientos a nivel del los organismos y no, como debe ser, en diferencias a nivel de las características emergentes de los grupos. Este es el error más común en la discusión sobre selección grupal.
AGRADECIMIENTOS
Quiero agradecer al Instituto colombiano para el desarrollo de la ciencia y la tecnología, COLCIENCIAS, por el apoyo financiero del proyecto “Evolución de la socialidad en cánidos – fase I”. Este artículo es parte del contexto teórico derivado del desarrollo de dicha investigación. También agradezco a Rodrigo González Florian por su colaboración en la modelación y edición de la Fig. 1. Muchas gracias a Jesualdo Fuentes, María Camila Vallejo y los evaluadores de este artículo por su lectura cuidadosa y valiosas sugerencias.
BIBLIOGRAFÍA
AMBROSE SH. Paleolithic Technology and Human Evolution. Science. 2001;291(5509):1748-1753.
[ Links ]ARNOLD E, FISTRUP K. The theory of evolution by natural selection: a hierarchical expansion. Paleobiology. 1982;8:113-129.
[ Links ]ARNOLD JM, BRAULT S, CROXALL JP. Albatross Populations in Peril: A Population Trajectory for Black-Browed Albatrosses at South Georgia. Ecol Appl. 2006;16(1):419-432.
[ Links ]ASFAW B, WHITE T, LOVEJOY O, LATIMER B, SIMPSON S, SUWA G. Australopithecus garhi: A New Species of Early Hominid from Ethiopia. Science. 1999; 284(5414):629-635.
[ Links ]BOISSERIE J-R, GUY F, DELAGNES A, HLUKSO LJ, BIBI F, BEYENE Y, et al. New palaeoanthropological research in the Plio-Pleistocene Omo Group, Lower Omo Valley, SNNPR (Southern Nations, Nationalities and People Regions), Ethiopia. C R Palevol. 2008;7(7):429-439.
[ Links ]BRIGGS J, PEAT DF. Espejo y reflejo: del caos al orden. Primera edición. México, D.F., México: Editorial Gedisa S.A.; 1991.
[ Links ]CARROLL SB. The making of the fittest DNA and the ultimate forensic record of evolution. Primera edición. New York: W. W. Norton & Company; 2006.
[ Links ]COYNE JA, HOEKSTRA HE. Evolution of Protein Expression: New Genes for a New Diet. Curr Biol. 2007;17(23):R1014-R1016.
[ Links ]DAVIDENKO JM, PERTSOV AV, SALOMONSZ R, BAXTER W, JALIFE J. Stationary and drifting spiral waves of excitation in isolated cardiac muscle. Nature. 1992;355(6358):349-351.
[ Links ]DAWKINS R. River out of eden: a Darwinian view of life. Primera edición. New York: BasicBooks; 1995.
[ Links ]DOBZHANSKY T. Genetics and the Origin of Species. Primera edición. New York: Columbia University Press; 1937.
[ Links ]DOBZHANSKY T. Nothing in Biology Makes Sense Except in the Light of Evolution. Am Biol Teach. 1973(35):125-129.
[ Links ]DOVER GJ, BOYER SH, PEMBREY ME. F-Cell Production in Sickle Cell Anemia: Regulation by Genes Linked to beta Hemoglobin Locus. Science. 1981;211(4489):1441-1444.
[ Links ]ELDREDGE N. Unfinished synthesis: biological hierarchies and modern evolutionary thought. Primera edición. New York: Oxford University Press; 1985.
[ Links ]ELDREDGE N, GOULD SJ. Punctuated equilibria: An alternative to phyletic gradualism. En: Schopf TJM, editor. Models in paleobiology. San Francisco: Freeman, Cooper and Co; 1972. p. 82-115.
[ Links ]ELDREDGE N, SALTHE SN. Hierarchy and evolution. Ox Surv Evol Biol. 1984;1:184-208. FISHER RA. The genetical theory of natural selection. Primera edición. Oxford: Clarendom Press; 1930.
[ Links ] [ Links ]GOULD SJ. Darwinism and the Expansion of Evolutionary Theory. Science. 1982;216(4544):380-387.
[ Links ]GOULD SJ. The structure of the evolutionary theory. First ed. Cambridge, Massachusetts: The Belknap Press of Harvard University Press; 2002.
[ Links ]HATCHWELL E, GREALLY JM. The potential role of epigenomic dysregulation in complex human disease. Trends Genet. 2007;23(11):588-595.
[ Links ]JABLONKA E, LAMB MJ. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life. Primera edición. Cambridge, Massachusetts: MIT Press; 2006.
[ Links ]LADEN G, WRANGHAM R. The rise of the hominids as an adaptive shift in fallback foods: plant underground storage organs (USOs) and australopith origins. J. Hum. Evol. 2005;49:482-498.
[ Links ]LECHLEITER J, GIRARD S, PERALTA E, CLAPHAM D. Spiral calcium wave propagation and annihilation in Xenopus laevis oocytes. Science. 1991;252(5002):123.
[ Links ]LEVINTON J. Genetics, paleontology and macroevolution. Editorial Cambridge: Cambridge University Press; 1988.
[ Links ]LEWONTIN RC. The units of selection. Annu Rev Ecol Syst. 1970;1(1):1-18.
[ Links ]MAYR E. The growth of biological thought: diversity, evolution and inheritance. Primera edición. Cambridge, Massachusetts: The Belknap Press of Harvard University Press; 1982.
[ Links ]MCHENRY HM, COFFING K. Australopithecus to Homo: Transformations in Body and Mind. Annu Rev Anthropol. 2000;29:125-146.
[ Links ]MUÑOZ-DURÁN J. Correlates of speciation and extinction rates in the Carnivora. Evol Ecol Res. 2002;4(7):963-991.
[ Links ]MUÑOZ-DURÁN J. Sociabilidad en carnívoros: una paradoja evolutiva. En: Zerda LE, editor. Comportamiento Animal y Humano. Bogotá, D.C.: Universidad Nacional de Colombia; 2009. p. 56-66.
[ Links ]NICOLIS G, PRIGOGINE I. Exploring Complexity. Primera edición. New York: W.H. Freeman and Company; 1989.
[ Links ]PÁLSSON E. A cAMP Signaling Model Explains the Benefit of Maintaining Two Forms of Phosphodiesterase in Dictyostelium. Biophy J. 2009;97(9):2388-2398.
[ Links ]PERRY GH, DOMINY NJ, CLAW KG, LEE AS, FIEGLER H, REDON R, et al. Diet and the evolution of human amylase gene copy number variation. Nat Gen. 2007;39(10):1256-1260.
[ Links ]POWARS DR, CHAN L, SCHROEDER WA. The Influence of Fetal Hemoglobin on the Clinical Expression of Sickle Cell Anemia. Ann N Y Acad Sci. 1989;565(Sickle Cell Disease):262-278.
[ Links ]REID K, CROXALL JP. Environmental Response of Upper Trophic-Level Predators Reveals a System Change in an Antarctic Marine Ecosystem. Proc R Soc Lond B. 2001;268(1465):377-384.
[ Links ]ROSSI F, LIVERI MLT. Chemical self-organization in self-assembling biomimetic systems. Ecol Modell. 2009;220(16):1857-1864.
[ Links ]ROSTAND J. Introducción a la historia de la Biología. Primera edición. Bogotá, D.C.: Planeta- De Agostini; 1985.
[ Links ]STANLEY SE. A theory of evolution above the species level. Proc Natl Acad Sci U S A. 1975;72:646-650.
[ Links ]STEDMAN HH, KOZYAK BW, NELSON A, THESIER DM, SU LT, LOW DW, et al. Myosin gene mutation correlates with anatomical changes in the human lineage. Nature. 2004;428:415-418.
[ Links ]STEINBOCK O, SIEGERT F, MULLER SC, WEIJER CJ. Three-Dimensional Waves of Excitation During Dictyostelium Morphogenesis. Proc Natl Acad Sci U S A. 1993;90(15):7332-7335.
[ Links ]THOMSON RB, BUTTERWORTH DS, BOYD IL, CROXALL JP. Modeling the Consequences of Antarctic Krill Harvesting on Antarctic Fur Seals. Ecol App. 2000;10(6):1806-1819.
[ Links ]TISHKOFF SA, REED FA, RANCIARO A, VOIGHT BF, BABBITT CC, SILVERMAN JS, et al. Convergent adaptation of human lactase persistence in Africa and Europe. Nat Gen. 2006;1:1-10.
[ Links ]VALEN LV. Group Selection, Sex, and Fossils. Evolution. 1975;29(1):87-94.
[ Links ]VALENTINE JW, MAY CL. Hierarchies in biology and paleontology. Paleobiology. 1996;22(1):23-33.
[ Links ]VAN VALEN L. A new evolutionary law. Evol Theor. 1973;1:1-30.
[ Links ]VOIGHT BF, KUDARAVALLI S, WEN X, PRITCHARD JK. A map of recent positive selection in the human genome. PLoS Biol. 2006;4(3):e72.
[ Links ]VRBA ES. Pattern and process in the sister-group Alcelaphini-Aepycerotini (Mammalia: Bovidae). En: Eldredge N, Stanley SM, editores. Living Fossils. New York: Springer-Verlag; 1984a.p. 62-79.
[ Links ]VRBA ES. What Is Species Selection. Syst Zool. 1984b;33(3):318-328.
[ Links ]VRBA ES. Testing Models of Macroevolution Examples from Miocene-Recent African Mammals. S Afr J Sci. 1985;81(6):300.
[ Links ]VRBA ES, ELDREDGE N. Individuals, hierarchies and processes: Towards a more complete evolutionary theory. Paleobiology. 1984;10(2):146-171.
[ Links ]VRBA ES, GOULD SJ. The Hierarchical Expansion of Sorting and Selection Sorting and Selection Cannot Be Equated. Paleobiology. 1986;12(2):217-228.
[ Links ]WOOD B, LONERGAN N. The hominin fossil record: taxa, grades and clades. J Anat. 2008;2 (12):354-376.
[ Links ]WYNNE-EDWADS VC. Intergruop selectionin the evolution of social systems. Nature. 1963;200:623-626.
[ Links ]
