










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.14 supl.1 Bogotá dez. 2009
MORFOLOGÍA FLORAL Y POLINIZACIÓN DE ORQUÍDEAS: EL SEGUNDO LIBRO DE CHARLES DARWIN
Floral Morphology and Pollination in Orchidaceae: Charles Darwin s Second Book
RODRIGO B. SINGER1. Ph. D. 1Depto. Botânica, Instituto de Biociências. Universidade Federal do Rio Grande do Sul. Porto Alegre, Brasil. rbsinger1@yahoo.com
Presentado 26 de noviembre de 2009, aceptado 15 de diciembre de 2010, correcciones 19 de mayo de 2010.
RESUMEN
El segundo libro de Darwin fue íntegramente dedicado a la morfología floral y polinización de diversos grupos de orquídeas de regiones templadas y tropicales. Este libro fue publicado en 1862 y parece haber sido concebido como una fuente de pruebas o un complemento para ideas sugeridas en El origen de las especies, en especial la noción sobre las ventajas del cruzamiento entre individuos diferentes, aunque sean hermafroditas (como es el caso de las orquídeas). La gran diversidad de morfologías florales y las diversas estrategias reproductivas que promueven la polinización cruzada en Orchidaceae fascinaron a Darwin, quien utilizó a este grupo de plantas como modelo para apoyar sus ideas. Darwin describió por primera vez y de modo impecable estrategias reproductivas como la protandria en orquídeas terrestres y la producción de flores imperfectas (unisexuales) en Catasetum, entre muchas otras contribuciones. Se analizan las ideas y propuestas de Darwin en este libro a la luz de nuestros conocimientos actuales y se muestran en gran parte correctas y vigentes.
Palabras clave: evolución, morfología floral, Orchidaceae, orquídeas, polinización, polinización cruzada.
ABSTRACT
Darwin s second book was totally dedicated to the floral functional morphology and pollination of temperate and tropical orchids. This book was published in 1862 and was likely conceived as an assemblage of evidence supporting ideas that were proposed in -On The Origin of The Species-; namely, the advantages of the intercrossing between different coespecific individuals, even if they are hermaphrodite (like the orchids). The great floral diversity and the outstanding number of reproductive strategies that promote cross-pollination in Orchidaceae fascinated Darwin who, in turn, used this plant group as a model to support his ideas. Darwin described for the first time and in a very accurate way, orchid reproductive strategies that clearly promote cross-pollination, such as protandry in terrestrial orchids and the production of unisexual flowers in Catasetum, among many other important contributions. Darwin s ideas and proposals regarding Orchidaceae are analyzed on the light of our present knowledge and prove to be mostly correct and valid.
Key words: cross-pollination, evolution, floral morphology, Orchidaceae, orhids, pollination.
CONTEXTO HISTÓRICO Y FINALIDAD DE LA OBRA
En 1862, pocos años después la publicación de El origen de las especies (Darwin, 1859), Darwin publicó su segundo libro, íntegramente dedicado a la morfología funcional y a la polinización de diversos grupos de orquídeas terrestres y epífitas. Esta obra tuvo el pomposo título de On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effect of intercrossing y ya desde el título despierta curiosidad. ¿Por qué un autor con tan vastos conocimientos en tantas áreas de la historia natural de tantos grupos biológicos escoge solamente a las orquídeas para su segundo libro? La respuesta aparecerá luego en el prólogo del libro (Darwin, 1862), pero antes veamos un poco el contexto histórico en el cual fue escrito.
Después del viaje alrededor del mundo en el H. M. S. Beagle, Darwin regresó al Reino Unido, se casó con su prima, Emma Wedgwood y se estableció en una bella casa, conocida como Down House, en el Condado de Kent, no muy lejos de Londres. En esta casa Darwin llevó una vida bastante recluida y escribió su legado, incluido El origen de las especies (Darwin, 1859), el libro de las orquídeas (Darwin, 1862) y el resto de su magnífica obra. En esta casa de campo Darwin tuvo un invernadero donde cultivó, entre otras cosas, orquídeas recibidas directamente del Jardín Botánico Real, hoy mundialmente conocido con el nombre de Kew Gardens.
De este modo, y a lo largo de años, Darwin efectuó detalladas observaciones de morfología floral en orquídeas cultivadas provenientes de las Américas, Asia y Madagascar. Además, Darwin fue lo suficientemente afortunado de vivir en una región que albergaba poblaciones naturales de orquídeas terrestres nativas. En Down House hay un sendero que hoy está flanqueado por árboles, pero que parece haber sido mucho más abierto y descampado en aquella época. Hoy ese sendero o camino es mundialmente conocido con el nombre que Darwin le dió: Sandwalk. Darwin acostumbraba caminar en el Sandwalk en compañía de un perro mientras meditaba sus ideas. Al final de este sendero Darwin encontraba una región más o menos rocosa, donde existían poblaciones naturales de orquídeas terrestres en las que él pudo hacer diversas y completas observaciones. Además, Darwin contó con la gentil colaboración de naturalistas de diferentes partes del mundo que generosamente compartieron sus informaciones con él o le enviaron plantas vivas o preservadas de algún modo que permitiese examinar sus flores.
Pero retomemos la pregunta inicial: ¿por qué las orquídeas como tema central de un libro? Pues porque Darwin percibió que gran parte de las diversas adaptaciones florales de las orquídeas pueden tornar el proceso de polinización más eficiente en muchos aspectos. Al mismo tiempo, Darwin notó que algunas adaptaciones florales tienen como función atraer y direccionar el comportamiento de los polinizadores en las flores y aumentar las posibilidades de polinización cruzada. En otras palabras, las orquídeas como grupo taxonómico son óptimas para estudiar evolución.
Ya en el prólogo del libro sobre orquídeas Darwin dice
In my volume ‘On the Origin of Species I have given only general reasons for my belief that it is apparently a universal law of nature that organic beings require an occasional cross with another individual; or, which is almost the same thing, that no hermaphrodite fertilises itself for a perpetuity of generations. Having been blamed for propounding this doctrine without giving ample facts, for which I had not, in that work, sufficient space, I wish to show that I have not spoken without having gone into details.
O sea, Darwin utilizó este segundo libro para sustentar ideas que propuso en El origen de las especies. Luego, Darwin dijo:
I have been led to publish this little treatise separately, as it has become inconveniently large to be incorporated with the rest of the discussion on the same subject. And I have thought, that, as Orchids are universally acknowledged to rank amongst the most singular and most modified forms in the vegetable kingdom, the facts to be presently given might lead some observers to look more curiously into the habits of our several native species. An examination of their many beautiful contrivances will exalt the whole vegetable kingdom in most persons estimation.
Así, este tratado está profundamente emparentado con El origen de las especies, pero Darwin juzgó mejor publicarlo como un trabajo separado. Entonces, el segundo libro de Darwin de alguna manera es una continuación o apéndice de El origen de las especies, utilizando las orquídeas como una especie de modelo biológico para explicar ideas antes sugeridas y, más concretamente, la necesidad y ventajas del cruzamiento entre individuos de la misma especie aunque sean hermafroditas, como en el caso de las orquídeas.
ESTRUCTURA DE LA OBRA
El libro consta de siete capítulos ilustrados con grabados en blanco y negro. Los grabados fueron hechos por un artista (G. M. Sowerby), con base en diagramas elaborados por el propio Darwin. En general, estas ilustraciones consisten en detalles de la morfología floral de los diferentes géneros que abarca la obra. En los cuatro primeros capítulos la obra trata de orquídeas de hábito terrestre oriundas del Reino Unido. Los capítulos cinco y seis tratan de orquídeas tropicales, principalmente de hábito epífito. Finalmente, el séptimo capítulo trata de encontrar homologías entre las estructuras observadas e intenta dar algunas explicaciones generales sobre todo lo observado. Los diferentes grupos de orquídeas estudiados por Darwin siguen una clasificación explícita, lo que torna a esta obra muy didáctica, pues los diferentes taxa son descritos y analizados por grupos taxonómicos y de un modo comparativo. La clasificación de orquídeas adoptada por Darwin es la de Lindley, 1830-1840, que aunque está lejos de ser idéntica, tiene bastantes puntos en común con la clasificación actualmente más aceptada (Chase et al., 2003), lo que facilita bastante la comparación de las observaciones de Darwin con lo que sabemos hoy de estos grupos de orquídeas.
MORFOLOGÍA FLORAL DE ORCHIDACEAE
Darwin era un excelente morfólogo y antes de comenzar con sus observaciones focales, brinda al lector con un resumen de morfología floral de Orchidaceae, de modo a establecer un marco o contexto de comprensión. En general, los términos morfológicos utilizados por Darwin son idénticos o muy similares a los que se utilizan en la actualidad (Dressler, 1993) y solo se harán en este artículo distinciones cuando hay discrepancias. Las flores de las orquídeas, como las de la gran mayoría de las monocotiledóneas, son 3-meras (Fig. 1), con un verticilo externo de tres sépalos y uno interno de tres pétalos (Fig. 1). El pétalo mediano es mayor, frecuentemente más colorido y/u ornamentado, y en general se conoce como labelo (Fig. 1). El androceo y el gineceo de la inmensa mayoría de las orquídeas están fusionados en una única estructura denominada columna (Fig. 1 ;Fig. 2A-C). La mayoría de las orquídeas presenta una única antera fértil (Fig. 2A) y mucho más raramente dos o tres. En la mayoría de los casos, el polen se encuentra aglutinado en unidades discretas llamadas polinias. El color, número, formato y textura de las polinias varía de acuerdo con el grupo taxonómico de orquídeas (Fig. 2B-C). El gineceo consiste de tres lóbulos estigmáticos, siendo que parte del lóbulo estigmático mediano se torna no-receptiva. Esta región no receptiva es llamada rostelo y puede secretar una sustancia pegajosa (viscid matter, para Darwin) que auxilia en el proceso de polinización adhiriendo las polínias al polinizador. En otras orquídeas, parte del rostelo se convierte en una superficie destacable que, cuando es removida por los polinizadores, se lleva adheridas a las polínias. Esta estructura destacable se denomina viscídio (Fig. 2B-C), pero Darwin, 1862, la llamaba -disco víscido- (viscid disc). El conjunto de las polinias y cualquier otra estructura o secreciones florales que auxilien su remoción polinario (Fig. 2B-C).

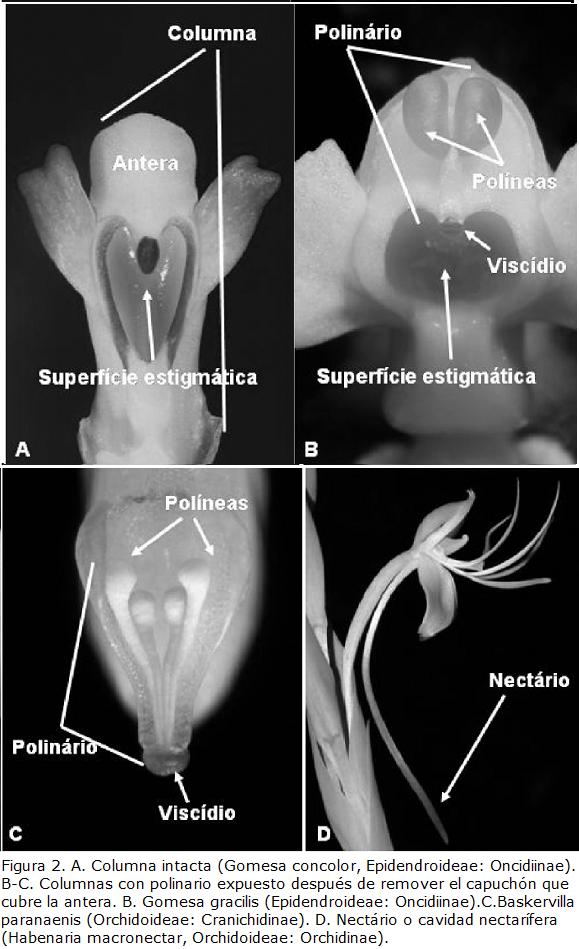
A lo largo del libro, Darwin llamó la atención sobre la compleja morfología floral de las orquídeas que él estudió, intentando ofrecer explicaciones funcionales para las estructuras que él describe. Así, con frecuencia las orquídeas que ofrecen néctar lo hacen en estructuras largas, como espolones que, en general, son prolongamientos del labelo (Fig. 2D). El néctar suele estar presente en el fondo de estos espolones, como una columna de líquido (Darwin, 1862). Nectarios, cavidades nectaríferas o que Darwin interpretó como nectaríferas fueron descritas en especies de los géneros Orchis, Habenaria, Spiranthes, Peristylus, Cattleya, Dendrobium y Angraecum, entre otros (Darwin, 1862). Para Darwin está claro que la presencia de estos largos nectarios o cavidades indica qué insectos con probóscides o lenguas largas deben ser los polinizadores; de otro modo no se entendería la longitud de estas estructuras o la presencia del néctar al final de largas cavidades.
Darwin mencionó haber encontrado néctar disponible en las orquídeas terrestres Gymnademia conopsea, Gymnademia albida, Habenaria bifolia, Habenaria chlorantha y Peristylus o Habenaria viridis (el propio Darwin dudaba sobre la identidad taxonómica de esta última planta).
Darwin, 1862, analizó flores frescas de varias especies de Orchis, sin encontrar néctar disponible en los supuestos nectarios. Sin embargo, Darwin percibió que los tejidos que forman estas cavidades son muy suculentos y liberan abundante secreción cuando son cortados. Darwin, 1862, entonces propone que aunque en estas flores no hay néctar disponible, las probóscides de los polinizadores deben lacerar los tejidos haciendo que los insectos puedan ingerir esta savia como si fuese néctar. Sprengel, 1793, ya había propuesto que estas son flores que engañan a sus polinizadores aparentando ofrecer néctar, una idea que Darwin, 1862, rechazó vehementemente. Curiosamente, lo que hoy sabemos sobre la polinización de estas especies de Orchis (van der Cingel, 1995) parece darle la razón a Sprengel, 1793: todo indica que estas orquídeas no producen néctar y que los polinizadores aprenden a reconocerlas como flores sin recompensa en pocos días de contacto en el ambiente. Así, la polinización y fructificación son limitadas, pues después de algunos días los insectos pasan a evitarlas (van der Cingel, 1995).
Es importante resaltar el momento histórico en que Darwin, 1862, escribió esta obra. En aquella época se sabía solo una ínfima parte de lo que sabemos hoy sobre la biología floral de este grupo de plantas. Para entender mejor esto, es necesario hacer un pequeño comentario sobre recompensas florales.
Se entiende por recompensa floral cualquier tejido o secreción que pueden ser recolectados en las flores por algún grupo de animales que los utilizan en alguna parte de su ciclo de vida. Hoy sabemos que diversos grupos de orquídeas pueden ofrecer recompensas florales muy diferentes del néctar, tales como aceites, tricomas, resinas y compuestos aromáticos (van der Cincel, 2001; Singer et al., 2006; Stpiczynska et al., 2007). Por otro lado, sabemos también que una parte significativa de las orquídeas no ofrece ningún tipo de recompensa a sus polinizadores y que en ellas el proceso de polinización involucra engaño a los polinizadores, a veces con estrategias sorprendentes y sofisticadas (Pouyanne, 1917; Ayasse, 2006; Jersáková et al., 2006; Singer et al., 2006). En la época de Darwin se creía que las orquídeas, en esencia, ofrecían néctar a sus polinizadores. Además, la inmensa mayoría de los estudios de polinización hasta ese entonces habían sido hechos en Europa, cuya diversidad es mucho menor que la de los trópicos. Así, numerosas estrategias de polinización no eran ni siquiera conocidas en aquel tiempo.
En muchos casos, los detallados estudios morfológicos de Darwin, 1862, ayudarán a elucidar asuntos que serán tratados a seguir, como la existencia de diversos mecanismos funcionales que promueven la polinización cruzada, la existencia de tipos florales diferentes dentro de un mismo taxón y otras muchas inferencias importantes. En el capítulo 7, Darwin intentó encontrar patrones y buscar homologías dentro de la diversidad encontrada en Orchidaceae. Es importante resaltar que, lamentablemente, parte de las propuestas de Darwin, 1862, en este capítulo parten de una premisa equivocada, lo que comprometerá su validez. Para Darwin, el labelo (pétalo mediano) era una estructura de naturaleza mixta constituida por un pétalo y dos estambres fusionados. Hoy sabemos claramente que desde un punto de vista ontogenético el labelo es solamente un pétalo (Endress, 1994).
OBSERVACIONES DE POLINIZACIÓN
Debido a la compleja morfología floral descrita con anterioridad, la verificación de la polinización de las orquídeas es relativamente más fácil que en otros grupos de plantas. Esto es así porque (como fue explicado antes) en la gran mayoría de estas plantas el polen no está suelto, sino que es removido en su totalidad, en la forma de polinarios que son macroscópicos y fáciles de visualizar (Fig. 3A-B). A su vez, como la inmensa mayoría de las orquídeas el polen sale en su totalidad, es relativamente fácil percibir si una flor ha donado su polen o no y esta es una herramienta muy útil que será bastante utilizada por Darwin, 1862.

Darwin admitió haber hecho muy pocas observaciones de polinización. Gran parte de los datos que aparecen en su libro le fueron brindados por corresponsales en diferentes partes del mundo. Según el propio Darwin, 1862, en veinte años de trabajo con orquídeas él solo pudo ver unas mariposas buscando néctar en flores de Orchis pyramidalis y Gymnademia conopsea. Uno de estos colaboradores (Mr. F. Bond), por ejemplo, le envió a Darwin 23 especies de lepidópteros con polinarios de orquídeas que, de acuerdo con Darwin, en su gran mayoría pertenecían a Orchis pyramidalis. Los polinarios de esta orquídea se adhieren a lo largo de la probóscide de las mariposas; uno de los grabados originales (Darwin, 1862, Fig. 4: p. 21) muestra la cabeza de una mariposa, Acontia luctuosa, con la probóscide extendida y siete polinarios de O. pyramidalis adheridos. Darwin mencionó también haber observado polinarios de Habenaria sp. adheridos en lepidópteros.
A pesar de haber visto pocos insectos en flores, Darwin observó inflorescencias de algunas especies en algunas localidades (especialmente el Condado de Kent, Devonshire y Surrey) y percibió una notable variación en el número de flores intactas, flores que actuaron como donadoras de polen y flores que actuaron como receptoras de éste. Para Darwin, 1862, estaba claro que estas diferencias reflejaban discrepancias locales en la abundancia y actividad de insectos que actúan como agentes polinizadores. Darwin, 1862, observó inflorescencias de Ophrys muscifera a lo largo de años y pudo comprobar (aunque no vio a los polinizadores) que solo una parte de los polinarios eran removidos y una parte de las flores se tornaba en frutos, clara prueba de estar sucediendo polinización por algún grupo de animales. De un total de 207 flores observadas, menos de la mitad fueron visitadas. Darwin, 1862, reconoció que no entendía como podían ser polinizadas estas flores y citó la teoría del botánico y orquidólogo Robert Brown, quien creía que las flores de Ophrys imitaban a abejas, presumiblemente como una forma de protección, evitando visitas de otros insectos. Hoy sabemos que la naturaleza de la polinización en estas flores es muy diferente. En esencia, las flores mimetizan a las hembras vírgenes de ciertos Hymenoptera. Los zánganos de estos Hymenoptera intentan copular con las flores y las polinizan durante el proceso (Ayasse, 2006; Pouyanne, 1917; Singer et al., 2006).
AUTOPOLINIZACIÓN
Darwin dedicó una parte significativa del Capítulo II a detallar el mecanismo de autopolinización espontánea de Ophrys apifera. En esta orquídea terrestre europea el polinario se curva espontáneamente, saliendo de la antera y tocando pasivamente la superficie estigmática, lo que permite la autopolinización y la consiguiente fertilización. Este mecanismo es generalizado en esta especie y, como Darwin percibió, todas las flores de O. apifera se tornan en frutos, lo que no sucede con las especies no-autógamas. El mecanismo de autopolinización de O. apifera ha sido comprobado por otros autores (van der Pijl y Dodson 1966; van der Cincel, 1995).
MECANISMOS QUE PROMUEVEN LA POLINIZACIÓN CRUZADA
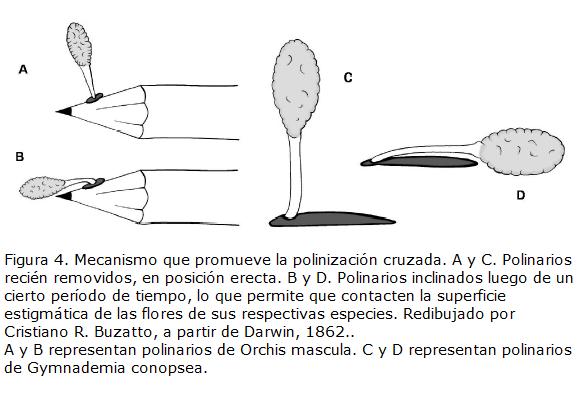
Darwin llamó la atención para diversos mecanismos que promueven la polinización cruzada tanto en orquídeas terrestres como epífitas. Así, Darwin, 1862, encontró un mecanismo semejante en Orchis mascula, Ophrys aranifera, Gymnademia conopsea y en especies de Oncidium, Odontoglossum, Vanda, Aerides, Sarcanthus, Saccolabium, Acropera, Maxillaria y otros. En estas orquídeas, cuando el polinario es removido, sale prácticamente erecto (Fig. 4A-C). Si el polinizador insistiese en visitar la misma flor o una muy próxima, el ángulo en que se encuentran todavía las polinias no les permitirá contactar la superficie estigmática. Después de un tiempo variable, se produce una inclinación del polinario (Fig. 4B-D). En esta posición, las polinias podrán contactar fácilmente la superficie estigmática. Como Darwin reconoció, este mecanismo promueve hasta un cierto punto la polinización cruzada y dificulta en cierto modo la autopolinización, pues entre la retirada del polinario por el polinizador y la deposición de las polinias en una superficie estigmática tendrá que mediar un breve período de tiempo, dando chance a que el polinizador se haya alejado de la planta de la que retiró el polen originalmente.
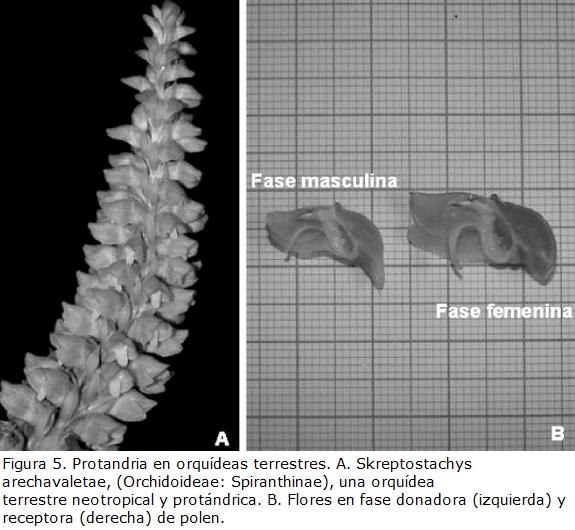
Darwin fue el primero en mencionar la existencia de protandria en orquídeas terrestres (Fig. 5A) del género Spiranthes (Spiranthinae). Protandria es el fenómeno por el cual flores perfectas (con androceo y gineceo) actúan primero como donadoras de polen (Fig. 5B) y solo después de un breve período de tiempo pueden actuar como receptoras de éste (Fig. 5B). Flores de Spiranthes recién abiertas solo actúan como donadoras de polen, pues la columna y el labelo se encuentran muy próximos. Así, el pasaje bajo la columna es muy estrecho y esto apenas permite que los polinizadores inserten sus piezas bucales para colectar néctar en la base del tubo floral y retiren el polinario adherido a ellas. Algunos días después, el ángulo entre la columna y el labelo se torna más abierto, exponiendo la superficie estigmática. En este momento la flor puede actuar como receptora de polen (Fig. 5B). Nuevamente, se trata de una adaptación morfológica que tiende a promover la polinización cruzada. La presencia de protandria ha sido confirmada en diversas especies de Spiranthes, así como en muchas orquídeas terrestres neotropicales filogenéticamente relacionadas (Singer et al., 2006).
LA FAMOSA ANGRAECUM SESQUIPEDALE
Mucha gente asocia el libro de orquídeas de Darwin con la polinización de Angraecum sesquipedale (una orquídea de Madagascar de la subtribu Angraecinae sensu Chase et al., 2003) por polillas Sphingidae de la especie Xantopan morganii praedicta. Casi todos los años diversas revistas de divulgación ilustran materias con reconstrucciones de la polinización de esta orquídea por este insecto, acreditándole a Darwin haber -predicho- esta interacción. Sin embargo, esta interacción (en la opinión de quien escribe) no ha sido satisfactoriamente demostrada hasta el presente. Es importante resaltar que hay abundante documentación sobre la polinización de otras orquídeas Angraecinae por polillas Sphingidae (Nilsson, 1998a; Nilsson, 1998 b; Nilsson et al., 1985; Nilsson et al., 1987) pero faltan todavía estudios concluyentes sobre Angraecum sesquipedale.
Darwin estudió en detalles el nectario de esta orquídea y afirmó que:
A whip-like green nectary of astonishing length hangs down beneath the labellum. In several flowers sent me by Mr. Bateman I found the nectaries eleven and a half inches long, with only the lower inch and a half filled with very sweet nectar. What can be the use, it may be asked, of a nectary of such disproportional length? We shall, I think, see that the fertilisation of the plant depends on this length and on nectar being contained only within thelower and attenuated extremity. Itis, however, surprising that any insect should be able to reach the nectar: our English sphinxes have probosces as long as their bodies: but in Madagascar there must be moths with probosces capable of extension to a length of between ten and eleven inches!- (Darwin, 1862, pág. 85).
Así, Darwin sugirió que en Madagascar debían existir polillas Sphingidae con una extensión posible de probóscide de hasta once pulgadas. Más tarde Darwin dijo:
The pollinia would not be withdrawn until some huge moth, with a wonderfully long proboscis, tried to drain the last drop. If such great moths were to become extinct in Madagascar, assuredly the Angræcum would become extinct- (...) We can thus partially understand how the astonishing length of the nectary may have been acquired by successive modifications. As certain moths of Madagascar became larger through natural selection in relation to their general conditions of life, either in the larval or mature state, or as the proboscis alone was lengthened to obtain honey from the Angræcum and other deep tubular flowers, those individual plants of the Angræcum which had the longest nectaries (and the nectary varies much in length in some Orchids), and which, consequently, compelled the moths to insert their probosces upto the very base, would be fertilised. These plants would yield most seed, and the seedlings would generally inherit longer nectaries; and so it would be in successive generations of the plant and moth. Thus it would appear that there has been a race in gaining length between the nectary of the Angræcum and the proboscis of certain moths; but the Angræcum has triumphed, for it flourishes and abounds in the forests of Madagascar, and still troubles each moth to insert its proboscis as far as possible in order to drain the last drop of nectar- (Darwin, 1862, p. 86-87).
Así, Darwin nos propone un posible contexto para la evolución del largo nectario de Angraecum: una -competencia- o carrera entre polinizador y polinizado.
Mucho de lo que se lee sobre el asunto ha sido distorsionado o exagerado. La polilla Xantopan morganii praedicta (Sphingidae), el supuesto polinizador, fue descrita bastante después del libro de Darwin (Rothschild y Jordan, 1903). En 1997, un entomólogo (Wasserthal, 1997) ofreció orquídeas Angraecinae (incluyendo Angraecum sesquipedale) para polillas Sphingidae nativas de Madagascar confinadas en ambientes cerrados, en laboratorio. En estas condiciones experimentales Xantopan morganii praedicta actuó como polinizador de Angraecum sesquipedale. Wasserthal (Wasserthal, 1997; Wasserthal, 1998) propuso una teoría alternativa para la evolución de los larguísimos nectarios en orquídeas Angraecinae. Para este autor (Wasserthal, 1997; Wasserthal, 1998), las larguísimas probóscides de algunas polillas Sphingidae de Madagascar pueden haber surgido como una adaptación de estas polillas para poder huir más fácilmente de arañas predadoras que las esperarían en las flores. Wasserthal (Wasserthal, 1997; Wasserthal, 1998) menciona en passant haber hecho trabajo de campo en Madagascar sin haber observado la polinización de A. sesquipedale. Como única evidencia -de campo-, Wasserthal (Wasserthal, 1997; Wasserthal, 1998) cita haber capturado un ejemplar de X. morganii praedicta con un viscídio de un polinario de orquídea adherido en su probóscide. Wasserthal (Wasserthal, 1997; Wasserthal, 1998) identificó este fragmento de polinario como siendo de A. sesquipedale, pero sin ofrecer una argumentación convincente a favor de ello. Dada la relevancia histórica de este tema, quien escribe no considera que este asunto haya sido debidamente documentado hasta el presente. Para otros autores (por ejemplo, Micheneau et al., 2009), el trabajo de Wasserthal (Wasserthal, 1997; Wasserthal, 1998) es la demostración de la -predicción- de Darwin. Quien escribe piensa que la interacción entre Xantopan morganii praedicta y Angraecum sesquipedale es, de hecho, muy probable, pero que todavía necesita ser demostrada de manera inequívoca.
OBSERVACIONES CON ORQUÍDEAS NEOTROPICALES
A lo largo de su libro, Darwin trató de adaptaciones morfológicas de otras orquídeas neotropicales. Darwin, 1862, estudió en detalle la morfología floral de numerosas especies de Cattleya (Fig. 6A) y propuso un mecanismo de polinización en que el rostelo debe humedecer el dorso del insecto polinizador y las polinias deben quedarse allí adheridas al ser removidas. Darwin sugirió que estas flores deben ser polinizadas por abejorros (Fig. 6B), no solo por los requisitos funcionales de las flores (los abejorros parecen ser morfológicamente muy apropiados para actuar como polinizadores debido a su tamaño y fuerza), sino también debido a evidencias circunstanciales. Darwin mencionó haber recibido un abejorro capturado en un invernadero con el polinario adherido en el dorso de su tórax, entre las alas. Este abejorro pertenecía a la especie Bombus hortorum y le fue enviado desde el British Museum por un investigador de apellido Smith, quien a su vez lo recibió de Sir W. C. Trevelyan, el horticultor que capturó el insecto.

Esta propuesta de Darwin es mucho menos conocida y, sin embargo, tiene muchas evidencias a su favor. Estudios recientes con especies actualmente incluidas en Cattleya (pues este género ha sido recientemente expandido con base en estudios filogenéticos) demuestran que los abejorros son particularmente importantes en la polinización de este género (Smidt et al., 2006; Silva-Pereira et al., 2007). Es importante resaltar que otras abejas neotropicales grandes (por ejemplo, abejas carpinteras - Fig. 6B-; euglossinas, etc.; Singer et al., 2006) y hasta colibríes pueden actuar como polinizadores de otras especies, pero el mecanismo de polinización de laCattleya es esencialmente el descrito por Darwin, 1862.
Darwin, 1862, efectuó también observaciones en las diminutas y complejas flores de algunas orquídeas Pleurothallidinae, especialmente con Masdevallia fenestrata (hoy Cryptophoranthus fenestratus) y admitió no entender como sucedía la polinización. Hoy sabemos que las orquídeas Pleurothallidinae (un grupo exclusivamente neotropical) son un grupo esencialmente polinizado por dípteros (moscas y grupos afines; Dressler, 1993; Singer et al., 2006).
Darwin dedicó casi un capítulo entero del libro a las orquídeas Catasetidae (hoy Catasetiinae, sensu Chase et al., 2003), a las que llamó -las más notables entre todas las orquídeas-. En particular, Darwin hizo observaciones muy detalladas en relación al género Catasetum (Fig. 6C-D). En aquella época, las especies de éste género estaban distribuidas en tres géneros diferentes: Myanthus, Monachanthus y Catasetum. Darwin realizó observaciones que permitieron confirmar que estos nombres estaban siendo empleados para formas funcionalmente diferentes de los mismos taxa. Catasetum era utilizado para plantas estaminadas, Monachanthus para flores carpeladas y Myanthus para flores hermafroditas, que hoy sabemos que son estériles.
Hoy en día el hecho por el cual las formas florales hayan sido clasificadas como géneros diferentes nos puede parecer muy extraño, pero sucede que las flores estaminadas (Fig. 6C) y carpeladas (Fig. 6D) de la misma especie de Catasetum son muy diferentes. Mientras las flores estaminadas son ricamente fimbriadas, coloridas y efímeras (pocos días de duración) (Fig. 6C), las carpeladas son verdes, rígidas, poco o nada ornamentadas y mucho más duraderas (Fig. 6D). Las conclusiones de Darwin se apoyaron también en observaciones precedentes (citadas por el propio Darwin) efectuadas por Sir Robert Schomburgk, Robert Lindley y Mr. Rodgers indicando la observación de producción de flores de Myanthus, Monachanthus y Catasetum en la misma planta.
Darwin citó ipsisliteris una frase de Lindley: -such cases shake to the foundation of all our ideas of the stability of genera and species-. Por otro lado, Darwin citó también observaciones adicionales de Sir Robert Schomburgk indicando que mientras las plantas silvestres de Cataseum y Myanthus no fructificaban, las de Monachanthus lo hacían abundantemente. Todo esto llevó a Darwin, 1862, a entender que en realidad los tres supuestos géneros no eran más que formas masculinas, femeninas y hermafroditas de un mismo género.
Hoy sabemos que las plantas que crecen en óptimas condiciones de luz y con buena nutrición tienden a producir solamente flores carpeladas. Plantas que crecen en locales sombreados y con nutrición más deficiente producen flores estaminadas. Así, la polinización cruzada en Catasetum es promovida a través de la producción de flores unisexuales, generalmente en plantas separadas. Un dimorfismo semejante es citado por Darwin para el género Cycnoches, que es filogenéticamente muy próximo de Catasetum.
Ya que hablamos de Catasetum, es importante resaltar que Darwin fue el primero en describir correctamente la morfología funcional de las flores estaminadas. En estas, el polinario se encuentra en tensión y ligeramente curvado. La columna tiene dos apéndices laterales que, al ser tocados, promueven, literalmente el -disparo- o eyección del polinário sobre el dorso del polinizador. En aquella época todavía no se sabía quien era el polinizador de estas flores. Hoy sabemos (Romero y Nelson, 1986, Singer et al., 2006) que son zánganos de abejas euglossinas, que mientras colectan sustancias aromáticas tocan involuntariamente los apéndices de la columna. En este momento, el polinário es eyectado en el dorso de las abejas, donde se adhiere firmemente. Las flores carpeladas, claro, no tienen polinario y producen las mismas substancias aromáticas que atraen a las euglossinas. Así, las abejas tenderán a permanecer más tiempo en las flores carpeladas, pues el impacto del polinário en el dorso parece ser un poco -traumático- (Romero y Nelson, 1986) para estos insectos, que al parecer evitan por un breve período las flores estaminadas y pasan a visitar las carpeladas, lo que promueve la polinización cruzada (la única posible en Catasetum).
Darwin también fue el primero en describir el mecanismo de eyección del polinário en Mormodes, otro género próximo de Catasetum, pero con flores perfectas. Las flores intactas tienen la columna curvada justo encima del labelo. La antera tiene un apéndice apical que está apuntado contra el labelo. Cuando este apéndice apical es tocado, el polinario es eyectado en el dorso del polinizador. Además de esto, la columna que ya ha donado su polinario, sufre una torsión gradual, al final de la cual la superficie estigmática queda expuesta, haciendo que la flor pueda ahora actuar como receptora de polen. En otras palabras, las flores de Mormodes son protándricas (actúan primero como donadoras de polen y después como receptoras de éste). Darwin, 1862, describió correctamente la eyección del polinario en este género, así como el hecho de que el polinario recién eyectado está parcialmente enrollado y se desenrolla gradualmente. Sin embargo, Darwin, 1862, aparentemente no percibió el cambio de ángulo de la columna en relación al labelo y la consecuente exposición de la superficie estigmática.
CONCLUSIÓN
¿QUÉ IDEAS PODEMOS SINTETIZAR DEL SEGUNDO LIBRO DE DARWIN?
Hay un conjunto de ideas que podemos extraer del segundo libro de Darwin, 1862. A pesar de los muchos avances en el estudio de la polinización de este grupo de plantas, estas ideas continúan esencialmente correctas y han servido, consciente o inconscientemente de norte para muchos estudios posteriores. Estas ideas subyacentes son:
- En las orquídeas hay una fuerte correlación entre la morfología floral y la polinización.
- La compleja morfología floral de las orquídeas tiende a tornar el proceso de polinización más eficiente.
- La mayoría de las orquídeas es polinizada por algún agente animal. La morfología floral de ciertas orquídeas se correlaciona con atributos morfológicos de sus polinizadores.
- Hay adaptaciones de morfología floral de muchos grupos de orquídeas que promueven la polinización cruzada.
- La polinización cruzada supone beneficios para las orquídeas y esto se demuestra por la gran cantidad de adaptaciones morfológicas que la promueven.
Como en muchas otras áreas, Darwin, 1862, resultó pionero en el estudio de la biología reproductiva de las orquídeas, un importante y diverso grupo de plantas. No por acaso, el estudio de la biología reproductiva y morfología funcional de las orquídeas está, desde entonces, fuertemente asociado a las ideas de evolución y adaptación. Desde fines del siglo XX, con la llegada y aceptación de la metodología cladística y la biología molecular, surge la posibilidad de reconstruir la filogenia (y, por lo tanto, la historia evolutiva) no solo de las orquídeas, sino de todos los grupos de organismos. Esto nos trae la oportunidad de poner a prueba ideas evolutivas como las de Darwin. En breve, la filogenia de la mayor parte de los grupos taxonómicos será conocida. Entonces será posible profundizar en el conocimiento de aspectos ecológicos dentro de estos grupos y podremos tener una idea sólida de cómo de hecho evolucionaron. ¿Realmente hay una carrera adaptativa entre las orquídeas Angraecinae y sus polinizadores?, ¿Cuántas veces evolucionó la protandria en las orquídeas terrestres?, ¿La producción de flores unisexuales en Catasetiinae surgió una o más de una vez? Tal vez estas serían las preguntas que Darwin querría responder hoy, pero cabrá a los investigadores actuales y futuros buscar las respuestas.
AGRADECIMIENTOS
A la Cátedra José Celestino Mutis de la Universidad Nacional de Colombia, sede Bogotá, por la oportunidad de volver a este asunto tan interesante y bonito. En especial a Lina Caballero Villalobos (Universidad Nacional de Colombia), por su ayuda en muchos aspectos y a Cristiano Roberto Buzatto (Universidade Federal do Rio Grande do Sul) por redibujar los esquemas de Darwin para este trabajo. Agradezco también, a dos revisores anónimos que ayudaron en mucho a mejorar el lenguaje y redacción de este trabajo.
BIBLIOGRAFÍA
AYASSE M. Floral scent and pollinator attraction in sexually deceptive orchids. En: Dudareva N, Pichersky E., editores. Biology of floral scent; 2006. p. 219-241.
[ Links ]CHASE MW, CAMERON K, BARRETT RL, FREUDENSTEIN JV. DNA data and Orchidaceae systematics: a new phylogenetic classification. En: Dixon KW, Barrett RL, Cribb P, editores. Orchid conservation. Kota Kinabalu: Natural History Publication (Borneo); 2003:69-89.
[ Links ]DARWIN C. The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray; 1859.
[ Links ]DARWIN C. On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effect of intercrossing. London: John Murray; 1862.
[ Links ]DRESSLER RL. Phylogeny and classification of the orchid family. Portland: Dioscorides Press; 1993.
[ Links ]ENDRESS PK. Diversity and evolutionary biology of tropical flowers. Cambridge, Mass: Cambridge University Press; 1994.
[ Links ]JERSÁKOVÁ J, JOHNSON SD, KINDLMANN P. Mechanisms and evolution of deceptive pollination in orchids. Biol Rev. 2006;81:219-235.
[ Links ]LINDLEY J. The genera and species of orchidaceous plants. Ridways, London. 1830-1840.
[ Links ]MICHENEAU C, JOHNSON SD, FAY M. Orchid pollination: from Darwin to the present day. Bot J Linn Soc. 2009;161:1-19.
[ Links ]NILSSON LA. Deep flowers for long tongues: reply from L. A. Nilsson. Trends Ecol Evol. 1998a;13:509.
[ Links ]NILSSON LA. Deep flowers for long tongues. Trends Ecol Evol. 1998b;13:259-260.
[ Links ]NILSSON LA, JONSSON L, RASON L, RANDRIANJOHANY E. Monophily and pollination mechanisms in Angraecum arachnites Schltr. (Orchidaceae) in a guild of long-tongued hawk-moths (Sphingidae) in Madagascar. Bot J Linn Soc. 1985;26:1-19.
[ Links ]NILSSON LA, JONSSON L, RALISON L, RANDRIANJOHANY E. Angraecoid orchids and hawkmoths in central Madagascar: specialized pollination systems and generalist foragers. Biotropica. 1987;19:310-318.
[ Links ]POUYANNE A. La fécondation des Ophrys par les insectes. Bulletin de la Société d Histoire Naturelle del Afrique du Nord. 1917;8:6-7.
[ Links ]ROMERO GA, NELSON CE. Sexual dimorphism in Catasetum orchids: forcible pollen emplacement and male flower competition. Science. 1986;232:1538-1540.
[ Links ]ROTHSCHILD LW, JORDAN K. A revision of the lepidopterous family Sphingidae. Novitates Zoologicae. 1903;9(S):1-972.
[ Links ]SILVA-PEREIRA V, SMIDT EDC, BORBA EL. Isolation mechanisms between two sympatric Sophronitis (Orchidaceae) species endemic to Northeastern Brazil. Plant Syst Evol. 2007;269(3-4):171-182.
[ Links ]SINGER RB, MARSAIOLI A, FLACH A, REIS MG. The ecology and chemistry of pollination in Brazilian orchids: recent advances. Chapter 64. En.: J. da Silva, editor. Floriculture, Ornamental and Plant Biotechnology. Middlessex: Global Science Books; 2006;4:570-583.
[ Links ]SMIDT EC, SILVA-PEREIRA V, BORBA EL. Reprodutive biology of two Cattleya (Orchidaceae) species endemic to north-eastern Brazil. Plant Species Biol. 2006;21(2):85-91.
[ Links ]SPRENGEL CK. Das entdeckte Geheimniss der Natur im Bau und in der Befruchtung der Blumen. Berlin: Friedrich Vieweg; 1793.
[ Links ]STPICZYNSKA M, DAVIES KL, GREGG A. Elaiophore diversity in three contrasting members of Oncidiinae (Orchidaceae). Bot J Linn Soc. 2007;155:135-148.
[ Links ]VAN DER CINGEL NA. An atlas of orchid pollination:European orchids. Rotterdam: A. A. Balkema Publishers; 1995.
[ Links ]VAN DER CINGEL NA. An atlas of orchid pollination: America, Africa, Asia and Australia. Rotterdam: A. A. Balkema Publishers; 2001.
[ Links ]VAN DER PIJL L, DODSON CH. Orchid flowers, their pollination and evolution. Coral Gables: University of Miami Press; 1966.
[ Links ]WASSERTHAL LT. The pollinators of the Malagasy star orchids Angraecum sesquipedale, A. sororium and A. compactum and the evolution of extremely long spurs by pollinator shift. Bot Acta. 1997;110:343-359.
[ Links ]WASSERTHAL LT. Deep flowers for long tongues. Trends Ecol Evol. 1998;13:459-460.
[ Links ]
