










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.14 suppl.1 Bogotá Dec. 2009
DARWIN Y LA DOMESTICACIÓN DE PLANTAS EN LAS AMÉRICAS: EL CASO DEL MAÍZ Y EL FRÍJOL
Darwin and Plant Domestication in the Americas: The Case of Maize and Bean
MARÍA ISABEL CHACÓN SÁNCHEZ1, Ph. D. 1Profesora Asociada, Facultad de Agronomía. Universidad Nacional de Colombia, Bogotá, Colombia. michacons@unal.edu.co
Presentado 5 de octubre de 2009, aceptado 27 de octubre de 2009, correcciones 19 de mayo de 2010.
RESUMEN
En este artículo se revisan los aportes que diversos autores, contemporáneos y posteriores a Darwin, han realizado sobre el estudio del origen de las plantas domesticadas, en especial al tema del origen geográfico de los cultivos. Posteriormente se revisa cómo la evidencia arqueológica junto con la evidencia genética han ayudado a comprender el origen de tres de los cultivos americanos más antiguos e importantes en épocas precolombinas y actuales: el maíz, el fríjol común y el fríjol Lima. Estos cultivos presentan patrones contrastantes de domesticación, mientras el maíz es producto de un solo evento de domesticación, el fríjol común y el fríjol Lima son cada uno de ellos resultado de domesticaciones múltiples e independientes. Se discute la relevancia del fenómeno de las domesticaciones múltiples en nuestra comprensión del origen del síndrome de domesticación y los aportes que la disciplina conocida como evolución de cultivos, junto con otras disciplinas, puede realizar para un mejor entendimiento de los procesos de adaptación en las poblaciones naturales y para acelerar el mejoramiento y la adopción de nuevos cultivos al servicio de la humanidad.
Palabras clave: arqueología, domesticación, evolución de cultivos, fitogeografía, genética.
ABSTRACT
In the present article, the main contributions that Darwin and other scholars, contemporary and posterior to him, made on the subject of plant domestication, in special the geography of crop origins, are reviewed. The article also reviews recent studies where the archaeological and genetic evidence have complemented each other to give a better understanding of the origin of three of the oldest and most important pre-Columbian and current crops: maize, common bean and Lima bean. These crops are examples of contrasting domestication patterns: a single domestication event in the case of maize and multiple and independent domestication events in the case of the common and Lima bean. Finally, the relevance of the phenomenon of multiple domestication in the study of the origin of the domestication syndrome is discussed along with the contribution that the discipline known as Crop Evolution can make in advancing our knowledge about adaptation processes in wild populations and the future of genetic improvement and adoption of new crops relevant for human welfare.
Key words: archaeology, crop evolution, domestication, genetics, phytogeography.
INTRODUCCIÓN
La domesticación de plantas es sin duda uno de los procesos evolutivos más importantes que ha acompañado al surgimiento y mantenimiento de la civilización humana (Diamond, 2002). El término domesticación proviene del Latín domus que significa -hogar- o -lugar de habitación- y hace referencia al hecho de traer una población silvestre, o población fundadora, cerca del lugar de habitación del hombre como es el campo de cultivo, el jardín, etc. (Harlan, 1992). El proceso de la domesticación tradicionalmente se ha considerado como un proceso evolutivo unidireccional en el cual las poblaciones silvestres fundadoras son las que responden (a través de cambios genéticos) a la acción ejercida por el hombre (i.e. selección artificial). Sin embargo, la domesticación podría ser también considerada como un proceso complejo de co-dependencia entre el hombre y las plantas que comienza con una interacción o asociación cercana entre éstos y en varios casos culmina en una mutua y total interdependencia. El maíz provee uno de los mejores ejemplos de una especie cereal totalmente domesticada, la cual depende del hombre para su dispersión (i.e. dispersión de sus semillas) y a su vez se ha convertido en uno de los principales sustentos de la humanidad junto con otras especies cereales como el trigo, la cebada y el arroz (Harlan, 1992).
Durante el proceso de domesticación acontecen progresivamente una serie de cambios morfológicos y fisiológicos, conocidos en su conjunto como el síndrome de domesticación, que favorecen la adaptación de las poblaciones silvestres fundadoras al medio ambiente de cultivo y la dependencia de éstas al hombre para su supervivencia. El síndrome de domesticación afecta principalmente las partes de la planta consumidas o usadas por el hombre (i.e. semillas y frutos) donde usualmente se presenta gigantismo (Fig. 1), y consiste también en cambios asociados al hábito de crecimiento de la planta (desde un hábito indeterminado hacia un hábito determinado), al modo reproductivo (favoreciendo en muchos casos la autogamia), a la pérdida de la dormancia de la semilla y a la pérdida o alteración del mecanismo natural de dispersión de la semilla, entre otros. La mayoría de estos rasgos no son fácilmente reconocibles en el registro arqueológico, sin embargo, el incremento del tamaño de la semilla y la pérdida del mecanismo natural de dispersión de ésta se han convertido en marcadores arqueológicos útiles en el estudio del proceso de la domesticación en los cereales (Fuller, 2007). La evidencia arqueológica basada en estos dos rasgos le permitió a Fuller, 2007, proponer un modelo de domesticación para algunas especies cereales originarias de la media luna fértil, en el cercano Oriente. Según este modelo, una respuesta adaptativa temprana en los cereales es el incremento del tamaño de la semilla, lo que se conoce como estado de semi-domesticación, seguido por la pérdida del mecanismo natural de dispersión de la semilla, lo que se conoce como domesticación sensu stricto. El tiempo que transcurre entre la semi-domesticación y la domesticación sensu stricto se define como la tasa de domesticación, que según este modelo puede tomar entre 1.000 y 1.500 años.

La evidencia genética, complementaria a la evidencia arqueológica, ha sido especialmente útil al establecer el origen biológico y geográfico de diversos cultivos, los cuales pueden tener un origen único (i.e. a partir de una población silvestre fundadora en un área geográfica restringida) o un origen múltiple (i.e. múltiples eventos de domesticación independientes a partir de diversas poblaciones silvestres fundadoras en diferentes regiones geográficas). El estudio del patrón de domesticación de una especie, tanto en espacio como en tiempo, es de particular importancia ya que éste determina en buena parte la estructura genética actual de dicha especie, lo cual es esencial para establecer estrategias de conservación y buen uso de estos recursos genéticos vegetales.
La gran cantidad de variedades vegetales domesticadas que existe maravilló desde un comienzo a Charles Darwin, quién dedicó a este tema el primer capítulo de su obra El origen de las especies (1859) el cual amplió en su obra titulada La variación de animales y plantas bajo domesticación (1868). En la primera parte del presente artículo, y en homenaje a los 200 años del nacimiento de Charles Darwin (1809-1882) y a los 150 años de la publicación de su obra El origen de las especies (1859), se revisan las contribuciones que Darwin y otros autores, contemporáneos y posteriores a él, realizaron sobre el tema del origen y evolución de las plantas domesticadas. En la segunda parte se revisa cómo la evidencia genética y arqueológica se han complementado para establecer los patrones de domesticación de una especie cereal como el maíz y de dos especies leguminosas del género Phaseolus en las Américas, las cuales permiten contrastar modelos de domesticación única y múltiple. Por último se presenta una reflexión sobre la contribución que la disciplina conocida como evolución de cultivos puede hacer para alcanzar una mejor comprensión del proceso de adaptación de las poblaciones silvestres durante el proceso de domesticación lo cual podría llevar en un futuro no muy lejano a la eventual domesticación de nuevos cultivos.
DARWIN, LA DOMESTICACIÓN Y OTROS AUTORES INFLUYENTES
Un tema de especial atención para Darwin era el mecanismo de herencia de los caracteres biológicos, y su interés en la domesticación no se debió solamente a que ésta proveía una analogía muy útil entre la selección artificial y la selección natural, la cual fue central en su libro El origen de las especies, sino también que sus experimentos con razas domesticadas le permitieron abordar temas relacionados con la variabilidad y la herencia, los cuales él consideraba centrales a su teoría de la selección natural (Darwin, 1859; Bartley, 1992). La importancia que Darwin le dio a la herencia se refleja en El origen de las especies cuando escribe: -Any variation which is not inherited is unimportant for us- ... -the laws governing inheritance are quite unknown- (Darwin, 1859, p. 12-13).
En El origen de las especies Darwin inicia con una discusión acerca de las diferencias que él observa entre las plantas cultivadas y sus parientes silvestres, en especial, el mayor grado de variación morfológica que existe entre las diferentes razas domesticadas de una misma especie, citando como ejemplo las diferentes variedades cultivadas de Brassica oleracea (coles de bruselas, coliflor, brócoli y repollo). Darwin señala muy acertadamente que las partes de la planta que muestran las mayores alteraciones morfológicas son las partes más valoradas por el hombre. En este sentido él escribe:
In regards to plants, there is another means of observing the accumulated effects of selection, namely, by comparing the diversity of flowers in the different varieties of the same species in the flower-garden; the diversity of leaves, pods, or tubers, or whatever part is valued in the kitchen-garden, in comparison with the flowers of the same varieties; and the diversity of fruit of the same species in the orchard, in comparison with the leaves and flowers of the same set of varieties.- (Darwin, 1859, p. 27).
Darwin cree que la existencia de la gran diversidad de razas domesticadas se debe al efecto acumulativo de la selección ejercida por el hombre, lo cual puede llevar a una condición en donde ya no es posible la reversión al estado silvestre. Con relación a este tema Darwin escribe:
...very many of the most strongly-marked domestic varieties could not possibly live in a wild state- -One of the most remarkable features in our domesticated races is that we see in them adaptation, not indeed to the animal s or plant s own good, but to man s use or fancy.-... (Darwin, 1859, p. 14, 25).
Adicionalmente, Darwin afirma que en parte nuestra dificultad en identificar el ancestro silvestre de las variedades domésticas se debe a la gran cantidad de cambios acumulados por estas últimas.
Darwin reconoció en el proceso de selección artificial dos componentes: un componente de selección metódica e intencional y un componente de selección inconsciente y no intencional. Al respecto Darwin escribe:
... Although man does not cause variability and cannot even prevent it, he can select, preserve, and accumulate the variation given to him by the hand of nature almost in any way which he chooses; and thus he can certainly produce a great result. Selection may be followed either methodically and intentionally, or unconsciously or unintentionally.- (Darwin, 1868, p. 3-4).
Mientras que la selección metódica es llevada a cabo intencionalmente con el fin de modificar una variedad de acuerdo a un plan preconcebido, la selección inconsciente resulta de aquellas actividades que el hombre realiza para el cultivo, cuidado y preservación de los individuos que él considera más valiosos sin ninguna intención de alterar la variedad domesticada y sin ninguna meta predeterminada (Darwin, 1868). Esta distinción entre estos dos componentes de la selección artificial fue fundamental en el desarrollo de su teoría ya que le permitió establecer una analogía específica entre la selección inconsciente y los procesos de selección natural.
Algunos de los principales rasgos de una planta domesticada, como son la pérdida del mecanismo de dispersión de la semilla y el gigantismo de la semilla, pueden resultar como respuesta a ciertas prácticas agrícolas, especialmente de cosecha, y como respuesta a la mayor competencia que existe entre plántulas de la misma especie en el campo cultivado, sin necesidad de evocar un proceso de selección intencional (Harlan, 1992). Por ejemplo, se considera que algo aparentemente tan simple como es el uso de un método específico de cosecha en el arroz silvestre puede determinar si esta especie cereal es o no domesticada; por ejemplo, si las semillas silvestres son golpeadas en una canasta entonces se cosecharán preferiblemente las semillas que se desprenden fácilmente y no se favorecerá la aparición del fenotipo domesticado (Vaughan et al., 2007).
En su obra La variación de animales y plantas bajo domesticación (1868), Darwin cita innumerables veces a Alphonse de Candolle (1806-1893), gran botánico suizo y autor de la obra Origin of Cultivated Plants con la cual marca el comienzo de la Fitogeografía. De Candolle establece diversos criterios que se deben tener en cuenta en el estudio del origen de las plantas domesticadas, entre ellos, la distribución de ancestros silvestres, la información histórica (usos, costumbres y tradiciones), la información lingüística (nombres locales de las variedades cultivadas) y la evidencia arqueológica (de Candolle, 1883). Posteriormente, Nicholas Ivanovich Vavilov (1887-1943), gran científico ruso y célebre autor de la obra Centers of Origins of Cultivated Plants (1926), e influenciado por las ideas de de Candolle desarrolla aún más el área de la Fitogeografía. Con el objetivo de colectar germoplasma útil que pudiera ser usado en programas de mejoramiento genético vegetal en la antigua Unión Soviética, Vavilov emprendió más de 100 viajes de recolección por todo el mundo y definió el centro de origen de aproximadamente 640 especies domesticadas (Vavilov, 1992). Basado en la premisa que el lugar de origen de un cultivo es el sitio de mayor diversidad de éste, Vavilov propuso inicialmente la existencia de al menos siete centros geográficos de domesticación, es decir, siete áreas relativamente pequeñas en donde un gran número de especies se habrían domesticado. Estos centros son: (1) Mesoamérica, (2) los Andes y la Amazonía, (3) el Mediterráneo, (4) el cercano Oriente, (5) Etiopía, (6) el sureste asiático y el pacífico sur, y (7) China.
Amigo de Vavilov y una de las figuras más influyentes en el desarrollo de la disciplina conocida como evolución de cultivos es el científico norteamericano Jack R. Harlan (1917-1998), autor de las obras clásicas Crops & Man (1992) y The Living Fields our Agricultural Heritage (1995). Durante buena parte de su carrera científica, Harlan se desempeñó como profesor de genética vegetal en el laboratorio de evolución de cultivos en el departamento de Agronomía de la Universidad de Illinois. En Illinois, Harlan tuvo acceso a información de toda índole relacionada con un número alto de cultivos la cual analizó de una manera holística y con rigor científico. Harlan llegó a resultados que en la mayoría de los casos se apartaban de la idea de centros de origen propuesta por Vavilov y concluyó que cada especie posee un patrón propio de domesticación que puede llegar a ser mucho más complejo y difuso que lo propuesto por Vavilov (Harlan, 1992; Harlan, 1995).
En la época de Darwin, de Candolle y Vavilov, la evidencia biológica se basaba principalmente en características morfológicas y agronómicas que tienen la desventaja de estar influenciadas por las condiciones medioambientales. A partir del desarrollo de los marcadores moleculares, la evidencia genética ha jugado un papel cada vez más importante en el estudio de varios aspectos relacionados con el proceso de domesticación (Burger et al., 2008), como por ejemplo la identificación de parientes silvestres (Olsen y Schaal, 1999), la identificación de áreas de domesticación (Matsuoka et al., 2002; Chacón et al., 2005; Kwak et al., 2009), el establecimiento de las bases genéticas del síndrome de domesticación (Xiong et al., 1999; Cai y Morishima, 2000; Burke et al., 2002; Peng et al., 2003) y la influencia de procesos demográficos (Ladizinsky, 1998a) y selectivos (Wang et al., 1999; Yamasaki et al., 2008) sobre la diversidad y estructura genética de las poblaciones domesticadas. A continuación se presenta cómo la evidencia genética se suma a la arqueológica para ayudar a establecer el área de origen y los patrones de domesticación de los cultivos.
PATRONES GEOGRÁFICOS DE LA DOMESTICACIÓN DE CULTIVOS: EL CASO DEL MAÍZ Y EL FRÍJOL
La evolución de las poblaciones silvestres fundadoras bajo domesticación está afectada por procesos no selectivos como la endogamia, el flujo génico y la deriva genética; por eventos demográficos como el cuello de botella y el efecto fundador (reducción de la diversidad genética en la población domesticada con relación a su ancestro silvestre); y por procesos selectivos, principalmente la selección artificial (Ladizinsky, 1998b). Los procesos no selectivos y los procesos demográficos afectarán de manera similar los niveles de diversidad genética en el genoma de las poblaciones, por el contrario, los procesos selectivos (i.e. selección natural y artificial) afectarán de manera específica las regiones del genoma que controlan los rasgos bajo selección y las regiones genómicas en alto desequilibrio de ligamiento con éstas (Black et al., 2001; Luikart et al., 2003). Por esta razón, la evidencia más robusta para establecer el área de origen y el número de veces que las especies vegetales han sido domesticadas proviene de regiones del genoma (nuclear y/o citoplasmático) consideradas -neutrales- desde el punto de vista de la domesticación ya que reflejan mejor la influencia de los eventos demográficos en el genoma. Estudios previos han indicado que algunas especies como el maíz se originaron en un área geográfica restringida y en un solo evento de domesticación (Matsuoka et al., 2002), por el contrario, otras especies como el fríjol común y el fríjol Lima se originaron a partir de eventos de domesticación múltiple (Chacón et al., 2005). La domesticación múltiple provee un modelo de particular interés para aquellos interesados en entender el proceso de adaptación durante la domesticación, en especial, la evolución paralela o convergente del síndrome de domesticación y sus bases genéticas. En el presente artículo no se pretende realizar una revisión exhaustiva de los patrones de domesticación de las plantas cultivadas, para lo cual el lector puede remitirse a excelentes trabajos (Harlan, 1975; Pickersgill y Heiser, 1977; Harlan, 1992; Zohary y Hopf, 2000; Pickersgill, 2007), sino que se enfoca en los patrones geográficos de domesticación contrastantes (único vs. múltiple) tomando como modelo dos de los cultivos más antiguos e importantes en la época precolombina en las Américas como son el maíz y el fríjol (del género Phaseolus).
DOMESTICACIÓN DEL MAÍZ: UN EJEMPLO DE ORIGEN ÚNICO
El maíz (Zea mays L.) es el único cultivo cereal domesticado en las Américas y uno de los cultivos mejor estudiados. En su libro -La variación de animales y plantas bajo domesticación- (Darwin, 1868), Darwin dedica los capítulos IX y X a las plantas cultivadas, entre ellas el maíz. Darwin nos comenta, citando a diferentes autores contemporáneos como de Candolle, que para la época ya había acuerdo entre los botánicos que las diferentes variedades de maíz pertenecían a la misma especie, que su origen era americano (aunque afirma que aún no se conocía su ancestro silvestre) y que su cultivo en las Américas era antiguo. Darwin llama la atención sobre la gran cantidad de -variedades- y -subvariedades- conocidas en la época las cuales mostraban diferencias en aspectos como la altura de la planta, la forma de la mazorca, el número de filas en las cuales se disponen las semillas, el color, el tamaño y la forma de las semillas, el contenido de glucosa, las cualidades culinarias, la resistencia a la sequía y en especial, algo que llamó mucho su atención era la adaptación latitudinal de diversas variedades. Actualmente se ha establecido que el maíz se originó, según evidencia botánica, arqueológica y genética, en el valle del río Balsas en México (Fig. 2) a partir de poblaciones silvestres de teosinte, específicamente de la especie Zea mays subsp. parviglumis Iltis y Doebley (Matsuoka et al., 2002; Hastorf, 2009; Piperno et al., 2009). Sin embargo, tomó varias décadas de arduo debate e investigación establecer quién era el pariente silvestre del maíz debido principalmente a la gran diferencia en la morfología de la inflorescencia femenina entre el maíz domesticado y el teosinte. En el teosinte la inflorescencia femenina es más corta y consiste de cinco a seis granos encapsulados en cubiertas duras en cada una de dos hileras por espiga, mientras que en el maíz se encuentran un número mucho mayor de granos no encapsulados (expuestos) arreglados en cuatro o más hileras. Las excavaciones de Richard MacNeish en los años 1960 (MacNeish, 1964) en el valle de Tehuacán en el centro de México (Fig. 2) recuperaron los restos arqueológicos de mazorcas de maíz domesticado más antiguos en aquella época con una edad de 3.000 años antes del presente (A.P.) y apuntó a México como la zona más probable de origen del maíz. Más tarde, Flannery recuperó restos arqueológicos mucho más antiguos de maíz domesticado con una edad de 6.300 A.P. en Guilá Naquitz, en el estado de Oaxaca en el sur de México (Piperno y Flannery, 2001; Fig. 2). Estos dos sitios arqueológicos están ubicados en regiones particularmente secas las cuales presentan unas condiciones apropiadas para la conservación a largo término de restos vegetales macrobotánicos (i.e. semillas y fragmentos vegetales). En el año 2002, Matsuoka y colaboradores con el uso de la evidencia genética basada en polimorfismos de loci microsatélites lograron establecer, mediante un estudio de distancias genéticas entre diversas especies de teosinte y diversas razas de maíz domesticado, que el maíz tiene un origen único a partir de la especie Z. mays subsp. parviglumis, particularmente de las poblaciones del Valle del río Balsas en el estado de Guerrero en México (Matsuoka et al., 2002; Fig. 2). Este sitio queda alejado de los sitios arqueológicos en Tehuacán y Oaxaca ya mencionados (Fig. 2) y está ubicado en una zona de bosque tropical estacional con condiciones menos favorables para la conservación de restos arqueológicos macrobotánicos. A raíz de este estudio, los arqueólogos concentraron sus investigaciones en el valle del río Balsas con el fin de elucidar el origen del maíz pero esta vez con base en evidencia microbotánica (fitolitos y gránulos de almidón asociados a sedimentos y herramientas de piedra). Estos estudios apoyan la hipótesis del uso del maíz en México, junto con calabazas domesticadas del género Cucurbita, en tiempos relativamente antiguos y en un contexto precerámico alrededor de 8.700 A.P., particularmente en el sitio conocido como Xihuatoxtla (Piperno et al., 2009; Fig. 2). Estos restos se constituyen en los registros arqueológicos más antiguos de maíz domesticado que se tengan hasta la fecha y, junto con la evidencia genética, apuntan al valle del río Balsas como la cuna del maíz. Vale la pena mencionar que estudios recientes de genética cuantitativa y molecular demuestran que relativamente pocas regiones en el genoma controlan las diferencias morfológicas tan dramáticas que existen entre el maíz cultivado y el maíz silvestre, siendo algunas de ellas explicadas por cambios regulatorios y no estructurales (Doebley et al., 2006)
PATRONES DE DOMESTICACIÓN EN EL GÉNERO PHASEOLUS: EVENTOS DE DOMESTICACIÓN MÚLTIPLES
El género Phaseolus, de origen americano, cuenta con más de 70 especies silvestres de las cuales cinco han sido llevadas a domesticación, Phaseolus vulgaris L., Phaseolus acutifolius A. Gray, Phaseolus coccineus L., Phaseolus lunatus L. y Phaseolus dumosusMacfady (Gepts, 1988). El síndrome de domesticación en el género Phaseolus y en general en las leguminosas consiste en rasgos como la presencia de vainas menos dehiscentes, incremento en el tamaño de las semillas (Fig. 1), pérdida de la dormancia e incremento en la variabilidad morfológica de las semillas (Weeden, 2007). Aún se desconoce si las bases genéticas del síndrome de domesticación entre especies o entre variedades domesticadas de la misma especie en el género Phaseolus son similares o no. Los parientes silvestres de P. acutifolius, P. coccineus y P. dumosus se distribuyen en Mesoamérica por lo cual su área putativa de domesticación posiblemente se circunscribe a esta región. Por el contrario, las poblaciones silvestres tanto de P. vulgaris (el fríjol común) como de P. lunatus (el fríjol Lima) están ampliamente distribuidas tanto en Mesoamérica como en sur América, las cuales han sido llevadas a domesticación al menos una vez en cada una de estas dos regiones (Fig. 2) y de manera independiente, por lo cual se constituyen en un excelente modelo para el estudio del fenómeno de la domesticación múltiple.
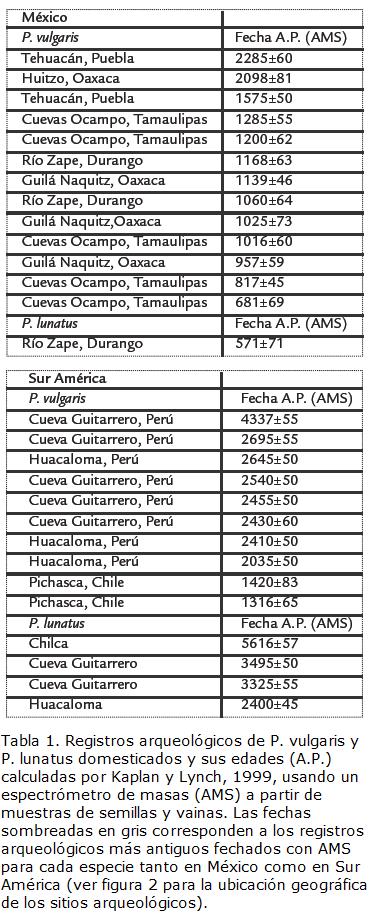
Parte de la evidencia arqueológica recuperada para el fríjol común y el fríjol Lima se muestra en la Tabla 1. Kaplan y Lynch, 1999, usaron un espectrómetro de masas (Accelerator Mass Spectrometry-AMS) para el análisis de restos arqueológicos vegetales (semillas y vainas) de estas dos especies provenientes de varios sitios arqueológicos de México y sur América con el fin de confrontar resultados previos obtenidos con el método indirecto del 14C, el cual se aplica comúnmente a muestras de carbón de madera asociadas a los restos macrobotánicos. La edad más antigua para el fríjol común domesticado proviene de la cueva Guitarrero en Perú con una edad de 4.337±55 A.P. A primera vista, este resultado sugiere que el fríjol común fue posiblemente domesticado en el Perú y posteriormente introducido en Mesoamérica donde los registros arqueológicos son más recientes (Tabla 1; Fig. 2). Un patrón similar se observa para el fríjol Lima domesticado cuyos restos arqueológicos más antiguos provienen de Chilca en el Perú con una edad de 5.616±57 A.P. y son mucho más recientes en México (Tabla 1; Fig. 2).
Phaseolus vulgaris silvestre se distribuye desde el estado de Chihuahua en el norte de México hasta San Luis en el norte de Argentina a una altura entre 700 y 2.700 m (Toro et al., 1990). A lo largo de su rango de distribución el fríjol común silvestre es variable, por ejemplo, las poblaciones silvestres en Mesoamérica presentan semilla pequeña mientras que las poblaciones andinas presentan semillas más grandes. En el área intermedia desde Costa Rica hasta Perú, el tamaño de la semilla es intermedio. En Mesoamérica, el fríjol común silvestre se encuentra en alturas que van desde los 1.500 hasta los 1.900 m sobre el nivel del mar (msnm; Delgado Salinas et al., 1988) mientras que en los Andes se encuentra en alturas que van desde 1.200 hasta 2.500 msnm (Brücher, 1988). Además de las diferencias morfológicas y ecológicas, existen también diferencias a nivel bioquímico y genético a tal punto que se han reconocido dos acervos genéticos en el fríjol común silvestre, el Mesoamericano y el Andino (Gepts et al., 1986; Khairallah et al., 1992; Chacón et al., 2007; Kwak y Gepts, 2009). Igualmente, en el fríjol común domesticado se reconocen dos acervos genéticos, uno de semilla pequeña de razas criollas mesoamericanas (razas Mesoamerica, Durango, Jalisco y Guatemala) y uno de semilla grande de razas criollas andinas (razas Perú, Nueva Granada y Chile). En los últimos años, varios estudios se han enfocado en el origen de las razas criollas y se ha establecido mediante evidencia genético-poblacional que las razas mesoamericanas provienen probablemente de uno o múltiples eventos de domesticación en diferentes regiones de México, mientras que las razas andinas parecen haberse originado en la región de Apurimac-Cuzco en los Andes peruanos a partir de un evento único de domesticación (Fig. 2) (Chacón et al., 2005; Kwak et al., 2009). El estudio de Chacón et al., 2005, estableció, con base en polimorfismos del ADN del cloroplasto, posibles áreas de domesticación en los estados de Durango, Jalisco y Oaxaca en México (Fig. 2), sin embargo no logró diferenciar entre un patrón de origen único y un patrón de orígenes múltiples para las razas mesoamericanas. El estudio de Kwak et al., 2009, identificó un área de posible domesticación en México en la cuenca de los ríos Lerma-Santiago en los estados de Jalisco y Guanajuato (Fig. 2), la cual está cercana al lugar de domesticación en Jalisco propuesto por Chacón et al., 2005. En la Figura 2 se puede observar que las tres áreas probables de domesticación propuestas para el fríjol común no coinciden con la propuesta para el maíz ni tampoco con el sitio arqueológico de Tehuacán donde se encontraron los restos arqueológicos más antiguos de fríjol común domesticado. En el futuro nuevas exploraciones arqueológicas y estudios genéticos pueden ayudar a aclarar la historia de domesticación de las razas mesoamericanas.
Phaseolus lunatus silvestre se distribuye desde Sinaloa en el norte de México hasta el norte de Argentina y en algunas islas del Caribe a alturas que van desde el nivel del mar hasta los 2.000 msnm. Al igual que en el fríjol común, en el Lima silvestre se han identificado dos grupos principales con base en el tamaño de la semilla y la distribución geográfica. Un acervo genético llamado mesoamericano se caracteriza por presentar semilla pequeña y por estar distribuido entre los cero y los 1.600 msnm desde México, pasando por centro América, el Caribe, Colombia y a lo largo de la vertiente oriental de los Andes de Perú, Bolivia y norte de Argentina (Debouck, 2008). El otro acervo genético llamado Andino, se caracteriza por su semilla grande y por estar distribuido a lo largo de la vertiente occidental de los Andes de Ecuador y norte del Perú entre los 400 y 2.000 msnm (Debouck, 2008). Sin embargo, con base en evidencia molecular, específicamente polimorfismos de regiones del ADN del núcleo y del cloroplasto, se ha establecido la existencia de tres acervos genéticos en el fríjol Lima silvestre llamados MI, MII y AI (Serrano-Serrano et al., 2010). MI y MII son acervos de semillas pequeñas y tienen un rango geográfico diferencial, mientras que MI ocupa tierras al norte y al oeste del istmo de Tehuantepec (Fig. 2), MII se encuentra principalmente al sur y al este del istmo de Tehuantepec, en América central, el Caribe y a lo largo de la vertiente oriental de los Andes de Perú, Bolivia y norte de Argentina. AI se caracteriza por tener semillas grandes y está distribuido a lo largo de la vertiente occidental de los Andes de Ecuador y norte del Perú (Fig. 2).
Al igual que las poblaciones silvestres, las poblaciones domesticadas también se clasifican en dos grandes grupos con base en el tamaño de sus semillas: el grupo de razas mesoamericanas de semillas pequeñas y redondeadas (cultivares Sieva y Potato) y el grupo de razas andinas de semillas grandes y aplanadas (cultivares Big Lima, ver Fig. 1; Baudet, 1977). Recientemente y con base en polimorfismos del ADN nuclear y del cloroplasto se ha establecido que las razas mesoamericanas posiblemente se originaron a partir de una sola domesticación en el área de distribución del acervo MI y que el origen de las razas andinas ocurrió mediante una sola domesticación en el área de distribución del acervo AI (Fig. 2; Motta-Aldana et al., comunicación personal). Vale la pena anotar que el área putativa de domesticación en los Andes está alejada del sitio arqueológico Chilca en la costa del Perú en donde se han hallado los registros más antiguos para fríjol Lima domesticado con una edad de alrededor de 5.616 A.P. Estos restos arqueológicos se encuentran en contexto precerámico y tomando en cuenta que el proceso de domesticación puede tomar entre 1.000 y 1.500 años al menos para los cereales (Fuller, 2007), la domesticación del fríjol Lima debió haber iniciado alrededor del año 7.000 A.P. en los Andes de Ecuador y norte del Perú y posteriormente habría sido introducido en otras áreas del Perú (incluyendo la costa). Dado que en el área putativa de domesticación AI del fríjol Lima actual-mente también se encuentra distribuido el fríjol común silvestre, uno se podría entonces preguntar, ¿por qué el fríjol común no fue también domesticado en esa misma área?
REFLEXIÓN FINAL
Gracias al avance, tanto conceptual como tecnológico, en campos aparentemente disímiles como son la arqueología y la genética, se ha logrado una mejor comprensión desde los tiempos de Darwin, De Candolle y Vavilov, del origen de las plantas cultivadas. El estudio de restos arqueológicos microbotánicos recuperados a partir de elementos de piedra y de sedimentos, como los fitolitos y los granos de almidón, abre nuevos escenarios para abordar el estudio del modo y el tiempo de la domesticación de plantas que usualmente ha ocurrido en regiones relativamente húmedas y en donde hay una pobre preservación de restos macrobotánicos. Los avances de la genética y en especial de la genómica, ha permitido el análisis de un número cada vez mayor de regiones del genoma y la identificación de los genes y los mecanismos que controlan el síndrome de domesticación. En este sentido, un estudio más detallado del fenómeno de las domesticaciones múltiples será muy fructífero en un futuro para poder establecer, por ejemplo, si en los procesos evolutivos los genes involucrados y los mecanismos de herencia para un rasgo fenotípico particular son o no similares entre diferentes variedades domesticadas producidas a partir de múltiples eventos de domesticación a nivel intra e interespecífico. Se espera que esta revisión haya generado en el lector inquietudes que conduzcan a un mayor conocimiento de la disciplina conocida como evolución de cultivos, que se encarga del estudio del origen biológico y geográfico de las especies cultivadas y de los procesos selectivos y no selectivos que han influenciado su actual estructura genética. La evolución de cultivos, junto con otras disciplinas, están llamadas a realizar aportes trascendentales que impacten el futuro nutricional y social de la humanidad a través del desarrollo de conocimiento científico y de su aplicación no solo al mejoramiento genético sino a la generación de nuevos cultivos.
BIBLIOGRAFÍA
BARTLEY MM. Darwin and domestication: studies on inheritance. J Hist Biol. 1992;25, 307-333.
[ Links ]BAUDET JC. Origine at classification des espèces cultivées du genre Phaseolus. Belg J Bot. 1977;110:65-76.
[ Links ]BLACK IV, WC, BAER CF, ANTOLIN MF, DUTEAU NM. Population Genomics: Genome-Wide Sampling of Insect Populations. Annu Rev Entomol. 2001;46:441-469.
[ Links ]BRÜCHER H. The wild ancestor of Phaseolus vulgaris in South America. En: GEPTS P, editor. Genetic Resources of Phaseolus beans. Dordrecht, Holland: Kluwer Academic Publishers; 1988. p. 185-214.
[ Links ]BURGER JC, CHAPMAN MA, BURKE JM. Molecular insights into the evolution of crop plants. Am J Bot. 2008;95:113-122.
[ Links ]BURKE JM, TANG S, KNAPP SJ, RIESEBERG LH. Genetic analysis of sunflower domestication. Genetics. 2002;161:1257-1267.
[ Links ]CAI HW, MORISHIMA H. Genomic regions affecting seed shattering and seed dormancy in rice. Theor Appl Genet. 2000;100:840-846.
[ Links ]CHACÓN MI, PICKERSGILL B, DEBOUCK DG. Domestication patterns in common bean (Phaseolus vulgaris L.) and the origin of the Mesoamerican and Andean cultivated races. Theor Appl Genet. 2005;110:432-444.
[ Links ]CHACÓN MI, PICKERSGILL B, DEBOUCK DG, ARIAS JS. Phylogeographic analysis of the chloroplast DNA variation in wild common bean (Phaseolus vulgaris L.) in the Americas. Plant Syst Evol. 2007;266:175-195.
[ Links ]DARWIN C. The Origin of Species, 1 ed. Wordsworth Editions Limited, London; 1859.
[ Links ]DARWIN CR. The variation of animals and plants under domestication, 1 ed, London: John Murray; 1868(1).
[ Links ]DE CANDOLLE A. Origin of Cultivated Plants, 1 ed. New York: Kessinger Publishing, LLC; 1883.
[ Links ]DEBOUCK DG. Notes sur les différents taxons de Phaseolus à partir des Herbiers - Section Paniculati. 2008. p. 233. Disponible en: URL: http://www.ciat.cgiar.org/urg.
[ Links ]DELGADO SALINAS A, BONET A, GEPTS P. The wild relative of Phaseolus vulgaris in Middle America. En: GEPTS P, editor. Genetic Resources of Phaseolus beans. Dordrecht, Holland: Kluwer Academic Publishers; 1988. p. 163-184.
[ Links ]DIAMOND J. Evolution, consequences and future of plant and animal domestication. Nature. 2002;418:700-707.
[ Links ]DOEBLEY JF, GAUT BS, SMITH BD. The molecular genetics of crop domestication. Cell. 2006;127:1309-1320.
[ Links ]FULLER DQ. Contrasting Patterns in Crop Domestication and Domestication Rates: Recent Archaeobotanical Insights from the Old World. Ann Bot. 2007;100:903-924.
[ Links ]GEPTS P. Genetic Resources of Phaseolus beans Current Plant Science and Biotechnology in Agriculture. Dordrecht: Kluwer Academis Publishers; 1988.
[ Links ]GEPTS P, OSBORN TC, RASHKA K, BLISS FA. Phaseolin-protein variability in wild forms and landraces of the common bean (Phaseolus vulgaris): evidence for multiple centers of domestication. Econ Bot. 1986;40:451-468.
[ Links ]HARLAN JR. Geographic patterns of variation in some cultivated plants. J Hered. 1975;66:184-191.
[ Links ]HARLAN JR. Crops & man. Second ed. American Society of Agronomy, Inc. Madison: Crop Science Society of America, Inc.; 1992.
[ Links ]HARLAN JR. The living fields: our agricultural heritage. Cambridge: Cambrige University Press; 1995.
[ Links ]HASTORF CA. Rio Balsas most likely region for maize domestication. Proc Natl Acad Sci U S A 2009;106:4957-4958.
[ Links ]KAPLAN L, LYNCH TF. Phaseolus (Fabaceae) in archaeology: AMS radiocarbon dates and their significance for pre-Columbian agriculture. Econ Bot. 1999;53:261-272.
[ Links ]KHAIRALLAH MM, SEARS BB, ADAMS MW. Mitochondrial restriction fragment length polymorphisms in wild Phaseolus vulgaris L.: insights on the domestication of the common bean. Theor Appl Genet. 1992;84:915-922.
[ Links ]KWAK M, GEPTS P. Structure of genetic diversity in the two major gene pools of common bean ( Phaseolus vulgaris L., Fabaceae). Theor Appl Genet. 2009;118:979-992.
[ Links ]KWAK M, KAMI JA, GEPTS P. The Putative Mesoamerican Domestication Center of Phaseolus vulgaris Is Located in the Lerma-Santiago Basin of Mexico. Crop Sci. 2009;49:554-563.
[ Links ]LADIZINSKY G. How many tough-rachis mutants gave rise to domesticated barley? Genet Resour Crop Evol. 1998a;45:411-414.
[ Links ]LADIZINSKY G. Plant evolution under domestication. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1998b.
[ Links ]LUIKART G, ENGLAND PR, TALLMON D, JORDAN S, TABERLET P. The power and promise of population genomics: from genotyping to genome typing. Nat Rev Genet. 2003;4:981-994.
[ Links ]MACNEISH RS. Ancient Mesoamerican Civilization. Science 1964;143:531-537.
[ Links ]MATSUOKA Y, VIGOUROUX Y, GOODMAN MM, SANCHEZ GJ, BUCKLER E, DOEBLEY J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc Natl Acad Sci U S A. 2002;99:6080-6084.
[ Links ]OLSEN KM, SCHAAL BA. Evidence on the origin of cassava: Phylogeography of Manihot esculenta. Proc Natl Acad Sci U S A. 1999;96:5586-5591.
[ Links ]PENG J, RONIN Y, FAHIMA T, RODER MS, LI Y, NEVO E, KOROL A. Domestication quantitative trait loci in Triticum dicoccoides, the progenitor of wheat. roc Natl Acad Sci U S A. 2003;100:2489-2494.
[ Links ]PICKERSGILL B. Domestication of Plants in the Americas: Insights from Mendelian and Molecular Genetics. Ann Bot. 2007;100:925-940.
[ Links ]PICKERSGILL B, HEISER CB. Origins and distribution of plants domesticated in the New World tropics. En: Reed CA, editor. Origins of agriculture. Paris: Mouton Publishers; 1977. p. 803-835.
[ Links ]PIPERNO DR, FLANNERY KV. The earliest archaeological maize (Zea mays L.) from highland Mexico: New accelerator mass spectrometry dates and their implications. Proc. Natl. Acad. Sci. USA. 2001:98.
[ Links ]PIPERNO DR, RANERE AJ, HOLST I, IRIARTE J, DICKAU R. Starch grain and phytolith evidence for early ninth millennium B.P. maize from the Central Balsas River Valley, Mexico. Proc Natl Acad Sci U S A. 2009;106:5019-5024.
[ Links ]SERRANO-SERRANO ML, HERNÁNDEZ-TORRES J, CASTILLO-VILLAMIZAR G, DEBOUCK DG, CHACÓN S. MI. Gene pools in wild Lima bean (Phaseoluslunatus L.) from the Americas: Evidences for an Andean origin and past migrations. Mol Phylogenet Evol. 2010;54:76-87.
[ Links ]TORO O, TOHME J, DEBOUCK DG. Wild bean (Phaseolus vulgaris L.): description and distribution. Cali, Colombia: Centro Internacional de Agricultura Tropical; 1990.
[ Links ]VAUGHAN DA, BALAZS E, HESLOP-HARRISON JS. From Crop Domestication to Super-domestication. Ann Bot. 2007;100:893-901.
[ Links ]VAVILOV NI. Origin and Geography of Cultivated Plants. Cambrige: Cambrige University Press; 1992.
[ Links ]WANG RL, STEC A, HEY J, LUKENS L, DOEBLEY J. The limits of selection during maize domestication. Nature. 1999;398:236-239.
[ Links ]WEEDEN NF. Genetic Changes Accompanying the Domestication of Pisum sativum: Is there a Common Genetic Basis to the -Domestication Syndrome for Legumes? Ann Bot. 2007;100:1017-1025.
[ Links ]XIONG LX, LIU KD, DAI XK, XU CG, ZHANG QF. Identification of genetic factors controlling domestication-related traits of rice using an F-2 population of a cross between Oryza sativa and O-rufipogon. Theor Appl Genet. 1999:98,243-251.
[ Links ]YAMASAKI M, SCHROEDER SG, SANCHEZ-VILLEDA H, GAUT BS, MCMULLEN MD. Empirical Analysis of Selection Screens for Domestication and Improvement Loci in Maize by Extended DNA Sequencing. Plant Genome. 2008;1:33-43.
[ Links ]ZOHARY D, HOPF M. Domestication of plants in the Old World, Third ed. New York: Oxford University Press; 2000.
[ Links ]
