










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.15 n.1 Bogotá jan./abr. 2010
ESTATUS DE LA POBLACIÓN DE MONOS AULLADORES ROJOS EN EL CAÑÓN DEL BARBAS, FRAGMENTO MONTANO, CORDILLERA CENTRAL, Colombia
Status of the red howler monkeys population in the Barbas canion, montane fragment, central mountains, Colombia
Jorge Mario Londoño, Ecólogo1; Carolina Gómez-Posada, MSc2. 1WCS Programa Colombia / Fundación EcoAndina. Filiación actual: Departamento de Postgrados, Universidad de Manizales. Manizales, Colombia aulladoresdelocaso@yahoo.com 2WCS Programa Colombia / Fundación EcoAndina. Filiación actual: Deparment of Biology, University of Washington. Seattle, cgomez@u.washington.edu
Presentado 2 de octubre 2008, aceptado 3 de junio de 2009, correcciones 27 julio 2009
RESUMEN
Los bosques de montaña en Colombia han sufrido fuerte degradación por causas antrópicas y en la actualidad la mayoría son remanentes aislados e inmersos en hábitat antropogénicos. Los aulladores rojos han persistido en estas condiciones pero poco se conoce sobre su biología y respuesta a la alteración de su ambiente. Durante cinco meses en el 2005, evaluamos densidad y estatus poblacional de esta especie en la cuenca del río Barbas (Departamentos de Quindío y Risaralda), fragmento aislado de bosque montano bajo de 790 ha. Estimamos la densidad empleando censos por transectos lineales repetidos. Adicionalmente seguimos tropas de aulladores y evaluamos variables de vegetación teniendo en cuenta los requerimientos ecológicos de la especie. Estimamos una densidad de 11.7±6 grupos y 132.1±68 individuos/km2, superior a la usualmente reportada en bosques continuos. Esta alta densidad probablemente se deba al aislamiento del fragmento, que limita las posibilidades de dispersión de los aulladores. El tamaño promedio por tropa fue 11.3±2.5 individuos, con más de cinco machos y siete hembras adultas, y de uno a cuatro inmaduros. En general el bosque ofrece buena disponibilidad de recursos estructurales y alimenticios para los aulladores. Esta población no se encuentra amenazada en la actualidad por actividades antrópicas. El fragmento ha sido recientemente protegido bajo la figura de Parque Regional Natural y se adelantan corredores de conexión con otros remanentes cercanos. Sin embargo, esta población de aulladores se encuentra aislada de otras poblaciones y por tanto es urgente desarrollar medidas de manejo del paisaje para atenuar los efectos del aislamiento.
Palabras claves: Alouatta seniculus, fragmento, densidad, estatus poblacional, bosque montano
ABSTRACT
The montane forests in Colombia have suffered strong degradation due to human activities; currently, most of them are remnants isolated and immersed in anthropogenic habitats. The red howler monkeys have persisted in these conditions but little is known on their biology and response to the alteration of their environment. During five months in 2005, we evaluated the population density and status of this species in the basin of the Barbas River (Quindío and Risaralda departments), a low montane isolated forest fragment of 790 ha. We estimated density by censuses for linear repeated transects. Additionally we followed troops of howlers and evaluated vegetation variables according to howler ecological requirements. We estimated a density of 11.7±6 groups and 132.1±68 individuals per km2, higher than what is usually reported for this species in continuous forests. This high density might be caused due to the isolation of the fragment that reduces the possibilities of dispersion of the howlers. The average troop size was 11±2.5 individuals, with more than five males and seven females, and one to four immature individuals. In general, the forest offers to the howlers, good availability of food and structural resources. Currently, this population is not threatened by anthropogenic activities. Recently, this fragment has been protected as a Regional Park, and it exists a project to develop corridors of connection with others remnants. Nevertheless, this howler population is isolated of other populations, and therefore urgent measures of landscape management are needed to attenuate the effects of the isolation.
Key words: Alouatta seniculus, fragment, density, population status, montane forest
INTRODUCCIÓN
Las actividades antrópicas han trasformado el bosque tropical en paisajes mixtos con predominio de monocultivos y extensos pastizales. La zona andina en Colombia es tal vez una de las más afectadas (se considera que solamente permanece el 30 % de la cobertura boscosa original), puesto que alberga gran parte de la población humana del país (Etter y Villa, 2000; Kattan y Álvarez-López, 1996). Los remanentes de bosque presentan tamaños y formas variables inmersos en una matriz de hábitat transformados. La mayoría de las áreas boscosas y zonas protegidas en las cordilleras colombianas se localizan por encima de los 2500 m de altitud, por lo que los bosques de alturas intermedias se encuentran en alguna medida en peligro (Cháves y Arango, 1997). Esta situación ha afectado la fauna de primates y actualmente el mono aullador rojo, Alouatta seniculus (Atelidae) es una de las pocas especies que aun persiste en la zona andina colombiana (por ejemplo, en el Eje Cafetero, además de aulladores rojos, solamente se han encontrado poblaciones de Aotus lemurinus y Cebus capucinus).
El mono aullador rojo es uno de los primates de mayor tamaño corporal en el neotrópico (~7 kg), es ampliamente reconocido por su sonoro aullido y presenta una amplia distribución en Sur América. A. seniculus se distribuye desde el norte de los Andes colombianos, hasta Venezuela, Trinidad, las Guyanas, y Brasil al norte del Amazonas, y en Ecuador, Perú, Bolivia y Brasil al occidente del Purus. El 20% del área de la distribución de esta especie está en Colombia (Defler, 2003). Los aulladores rojos habitan diferentes tipos de hábitat y se puede encontrar en bosques aislados e intervenidos. En Colombia se distribuyen por casi todo el territorio nacional, desde el nivel de mar hasta los 2400 m de altitud, con registros ocasionales a 3200 m (Defler, 2003; Hernández-Camacho y Cooper, 1976). Los aulladores viven en pequeños grupos familiares (6 a 9 individuos), son herbívoros (consumen frutos y hojas principalmente) y son animales de poco desplazamiento y baja actividad (como estrategia de ahorro de energía) (Defler, 2003; Milton, 1980). Dentro de los hábitat donde se encuentran los aulladores rojos en Colombia, los bosques de la cordillera de los Andes y en particular los de la región cafetera (departamentos de Quindío, Risaralda y Caldas principalmente), son los más alterados por la intervención humana.
A pesar de ser una de las especies más estudiadas en el neotrópico, la mayoría de la información de este mono proviene de selvas de tierras bajas y bosques continuos (Defler, 2003). Aunque la especie no es considerada en peligro de extinción, las poblaciones de montaña en Colombia podrían encontrarse amenazadas por el aislamiento y la fuerte intervención antrópica (Gómez-Posada, 2006; Defler, 2003). En este trabajo describimos la densidad y estatus poblacional de aulladores rojos en el Cañón del río Barbas, ubicado entre los Departamentos de Quindío y Risaralda, Colombia. Este remanente es uno de los fragmentos más grandes e importantes de bosque subandino en el eje cafetero del país. El Cañón del Barbas alberga la población más conocida y carismática de aulladores en la región, cuya presencia ha servido para generar oportunidades de conservación del bosque y de otras especies de fauna que lo habitan. Este trabajo aporta información sobre el estado actual de poblaciones de aulladores rojos en zonas montañosas y hace parte de un estudio a largo plazo que está documentando la respuesta de esta especie a la fragmentación y destrucción de sus hábitat (Gómez-Posada et ál., 2007; Gómez-Posada et ál., 2005; Londoño y Gómez-Posada, 2007; Gómez-Posada, 2006).
MÉTODOS
ÁREA DE ESTUDIO
La cuenca media del río Barbas, conocida como Cañón del Río Barbas se localiza entre los 4° 42´ N y 75° 38´ W entre los 1.500 y 2.100 m de altitud, sobre la vertiente occidental de la cordillera Central de los Andes Colombianos y es el límite entre los departamentos de Quindío y Risaralda. El Cañón del Barbas es un fragmento angosto y alargado (Fig. 1), de 790 ha de bosque que fueron protegidas históricamente por su alta pendiente (Franco y Bravo, 2005). Según los registros por fotografías aéreas, el fragmento lleva totalmente aislado al menos 60 años.
Estos bosques corresponden a la categoría de bosque muy húmedo montano bajo según Holdridge (Acero, 1985). La zona presenta una temperatura media entre los 17 y 24ºC y un promedio anual de lluvias de 1000 a 3000 mm (Acero, 1985). El área presenta bosques maduros y secundarios en diferentes estados de conservación y está rodeado por un mosaico de sistemas productivos: zonas de potrero para ganadería principalmente, grandes plantaciones forestales comerciales (predominantemente Eucalyptus spp. y Pinus patula) y cultivos pequeños de granadilla, café y plátano. En algunas áreas, se encuentra bajo presión, debido a la extracción de madera, semillas y bejucos, para elaboración de artesanías, y como materia prima para diversas actividades, principalmente de ebanistería.
El Cañón del Barbas es una de las manchas de bosque más grandes que se encuentran en esta zona altitudinal en la cordillera Central. Se han registrado cerca de 231 especies de aves, entre ellas, algunas endémicas y en peligro, como la pava caucana (Penelope perspicax) (Franco y Bravo, 2005). La mayoría del área es de carácter privado y recientemente ha sido nombrado Parque Regional (comunicación personal, Carlos Alberto Franco Cano, Director CRQ, septiembre 29 de 2008).
La región del Cañón del río Barbas ha sufrido de intervención antrópica desde tiempos pre-hispánicos. Sin embargo, el establecimiento de ciudades y la deforestación se presentaron hasta mediados del siglo XIX y principios del XX (CINEP, 2004). Para la región del río Barbas, solamente quedaron con cobertura boscosa las zonas con elevada pendientes en la cuenca (Franco y Bravo, 2005). La presión sobre el bosque ha ido disminuyendo paulatinamente en los últimos años, gracias una naciente toma de conciencia ambiental y a las actividades de educación desarrollados en la zona por instituciones locales, apoyadas por el Instituto von Humboldt. En la actualidad la frontera agrícola parece estar estable y por el contrario se han dejado abandonadas áreas que actualmente se aprecian como bosques en diferentes estados de sucesión. La caza de animales ha sido prohibida en los últimos años y los monos aulladores gozan de aprecio y admiración entre la comunidad.
Alrededor del Cañón del Barbas, se encuentran otros fragmentos de bosque de diferentes tamaños, aislados entre si por potreros para ganadería, cultivos y/o carreteras (Fig. 1). Estos fragmentos se encuentran separados del Cañón del Barbas por distancias superiores a los 250 m, por lo que es poco probable la dispersión de aulladores desde el Barbas (Mandujano et ál., 2004; Rodríguez-Toledo et ál., 2003; Estrada y Coates Estrada, 1996). Al costado oriental del Cañón, se encuentran los bosques de la cuenca alta del Barbas, donde han sido reportados aulladores. La cuenca alta y la cuenca media se encuentran aisladas por la autopista del Café, de alto tráfico vehicular. Adicionalmente, los bosques cerca de la autopista, se encuentra altamente degradados, por lo que se considera baja la posibilidad de dispersión de aulladores entre las dos los cuencas (Gómez-Posada, 2006). En ningún otro fragmento de la zona se han reportado aulladores.
Un kilómetro al sur del Cañón de Barbas, se localiza la Reserva Forestal Bremen-La Popa, de 747 ha, establecida a principios de la década del 70 para la protección de cuencas hidrográficas (Franco y Bravo, 2005). Actualmente se desarrolla en la región un proyecto de construcción de corredores de conexión del bosque del Cañón del Barbas con este remanente. Los aulladores del Barbas han empezado a registrarse en dichos corredores y se espera que ha futuro colonicen los bosques de Bremen-La Popa (comunicación personal, Fabio Lozano, Instituto Humboldt, junio de 2007).
ESTIMACIÓN DE LA DENSIDAD DE AULLADORES
El trabajo se desarrolló desde agosto a diciembre de 2005. Dentro del fragmento, delimitamos un área de 179 ha para la evaluación de la población de aulladores (Fig. 1). El área evaluada fue delimitada de acuerdo a la factibilidad del terreno para desarrollar el trabajo (la zona es muy pendiente y presenta áreas de difícil acceso), y abarcó los tipos de bosque presentes en el fragmento. Para estimar la densidad se utilizaron censos por transectos lineales repetidos (Peres, 1999; NRC, 1981). Los censos se llevaron a cabo en seis transectos que atravesaron el área evaluada, los cuales sumaron 6.1 km. Los transectos fueron ubicados con orientación sur - norte, separados por al menos 500 m entre si. Su ubicación dependió de la factibilidad del terreno para realizarlos. En cada transecto fueron realizados en promedio 16 censos, distribuidos equitativamente a lo largo del estudio. Cada transecto fue recorrido a una velocidad promedio de 0.5 km/hora, a las 8:00 horas, con un recorrido inverso del mismo transecto a las 14:00 horas (en bosques montanos la presencia de neblina impide la realización de censos más temprano en la mañana).
Para cada transecto, fue calculada independientemente la densidad ecológica de grupos a partir de la formula D = n / (2*ESW*L) (Chiarello y Melo, 2001), donde n” es el número total de grupos observados en cada transecto a lo largo del estudio, ESW” la distancia de detección efectiva (estimada con el programa Distance 4.1) y L” es la longitud total recorrida para ese transecto durante todo el estudio. La densidad total de grupos fue obtenida como el promedio entre los seis transectos.
Durante los censos, de acuerdo a la metodología planteada, una vez ubicado un grupo, el investigador tiene poco tiempo para observar la tropa de monos. Este corto intervalo dificulta la determinación de la tropa completa de primates, lo cual conlleva a subestimar el tamaño de grupo (Pruetz y Leasor, 2002; Peres, 1999). Por consiguiente, se siguieron varios grupos fuera de los censos, con el objetivo de establecer su tamaño. La densidad de individuos fue estimada multiplicando la densidad de grupos, por el tamaño promedio de las tropas (Peres, 1999). Para estimar el tamaño de la población de aulladores en el Cañón del Barbas, utilizamos la densidad calculada de individuos y la extrapolamos al tamaño total del fragmento (790 ha).
ESTRUCTURA Y COMPOSICIÓN DE LOS GRUPOS DE AULLADORES
Adicionalmente a lo censos, se siguieron grupos de aulladores de la zona (estos grupos fueron detectados durante los censos), en los cuales se identificaron las tropas e individuos. En lo posible se especificó la edad y sexo (siguiendo las características morfológicas utilizadas por Soini (1992) y Defler (1981): 1) Adulto: individuo que ha alcanzado la madures sexual y tamaño definitivo. Macho con región hioidea muy desarrollada. 2) Subadulto: Tamaño casi adulto pero perceptiblemente menos corpulento. 3) Juvenil: Todavía de tamaño pequeño, llega a ser la mitad o un poco más del tamaño del adulto. Tiene locomoción propia, pero se asocia frecuentemente con la madre. 4) Infante: Cría pequeña, cargada constantemente por la madre.
DETERMINACIÓN DE LA COMPOSICIÓN Y ESTRUCTURA VEGETAL
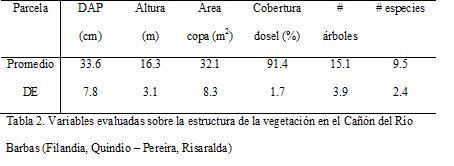
Para evaluar la disponibilidad de recursos alimenticios y estructurales para los aulladores en este bosque, fueron medidas algunas variables de composición y estructura vegetal según los requerimientos de este género de primates (Stevenson et ál., 2000; Estrada y Coates-Estrada, 1996; Soini, 1992; Yoneda, 1990; Schwarzkopf y Rylands, 1989). Con este fin, se realizaron 15 parcelas de área fija de 50 m de largo por 4 m de ancho. Estas se distribuyeron aleatoriamente dentro del área evaluada. En cada parcela fueron estimadas las siguientes variables: porcentaje de cobertura de dosel, DAP (de árboles mayores a 10 cm de DAP), altura, área de copa y especie de los árboles presentes.
RESULTADOS
ESTIMACIÓN DE LA DENSIDAD DE AULLADORES
A lo largo del estudio, se acumularon un total de 99.1 km recorridos durante los censos; y en cada transecto, se acumularon un promedio de 16.5 ± 9.1 km recorridos. Durante todo el muestreo, fueron registradas 41 observaciones de grupos de mono aullador, distribuidas en los seis transectos. A lo largo del estudio, fueron registrados 6.8 ± 4.4 grupos por transecto; y el promedio de observaciones fue 0.42 grupos por censo (desde 0 hasta 2). Esporádicamente fueron observados machos solitarios; éstos no fueron incluidos en los análisis. Los solitarios generalmente están pretendiendo unirse a algún grupo y su condición de solitarios es temporal.
La distancia perpendicular de registro de grupos de aulladores durante los censos, varió desde 0 hasta 55 m. La distribución de frecuencias de dichas distancias perpendiculares, calculada con el programa Distance 4.1, estimó una distancia de detección efectiva ESW = 11.9 m (CV=11%).
Fuera de los censos, se estableció el tamaño de seis tropas de aulladores, con un promedio de 11.3 ± 2.5 individuos. Reemplazando todos los datos anteriores en la fórmula explicada en métodos, fue calculada una densidad de 11.7 ± 6 grupos/km2 y de 132.1 ± 68 indv/km2. Extrapolando esta densidad al tamaño total del fragmento, se calculó la abundancia promedio del bosque en 92 tropas (intervalo de confianza 95%: 79 a 105 grupos), con 1044 individuos (intervalo de confianza 95%: 895 a 1102 individuos).
ESTRUCTURA Y COMPOSICIÓN DE LOS GRUPOS DE AULLADORES
Fue identificada la estructura y composición de seis tropas, con un tamaño promedio de 11.3, el cual varió desde 3 hasta 15 individuos (Tabla 1). Los grupos estuvieron conformados por tres a cinco machos adultos y subadultos, tres a siete hembras adultas y subadultas y de uno a cuatro individuos inmaduros (los cuales sumaron el 27 % de la población). La relación hembras a machos adultos fue de 1 : 0.75 y de hembras adultas a inmaduros de 1 : 0.82.
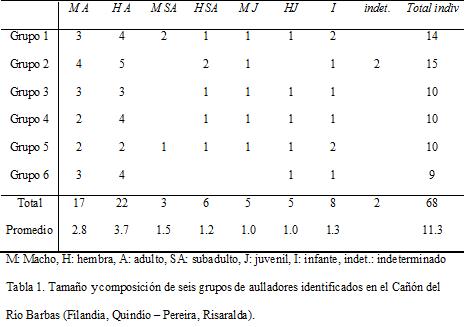
DETERMINACIÓN DE LA COMPOSICIÓN Y ESTRUCTURA VEGETAL
En la Tabla 2 se encuentra el resumen de las variables de vegetación evaluadas. La mayoría de los árboles presentaron gran porte, con DAP mayores a 25 cm, copa mayor a 24 m2 y alrededor de los 16 m de altura. La conectividad de dosel fue superior al 90 %.
En las parcelas fueron encontrados 210 árboles, distribuidos en 69 especies y 21 familias. Las familias más abundantes fueron Laureceae (con cuatro géneros) y Moraceae (con tres géneros). Las especies más comunes fueron Otoba lhemanni (7% de los individuos) y Cecropia telealba (5%). Del total de árboles registrados en las parcelas, el 32 % pertenecieron a 18 especies consumidas por los aulladores: Morus insignis, Miconia sp., C. telealba, Inga sp., O. Lhemanni, Garcinia sp., Allophylus mollis, Nectandra sp., Pouteria lucuma, Pseuldolmedia rigida y ocho especies de Ficus. Los aulladores utilizaron las plantaciones forestales como corredores de conexión entre parches de bosque natural y algunos grupos han establecido parte de su área de actividad en las plantaciones de mayor edad (superior a 10 años).
DISCUSIÓN
A. seniculus presenta densidades en tierras bajas generalmente entre 25 y 55 indv/km2, pero se encuentran reportes desde 4 hasta más de 100 indv/km2 (Defler, 2003; Palacios y Rodríguez, 2001; Chapman y Balcomb, 1998; Crockett y Eisenberg 1987). La densidad de esta especie puede variar dependiendo de algunos factores como las características intrínsecas del hábitat, heterogeneidad y estacionalidad de los bosques, competencia, aislamiento de remanentes boscosos, alteración del hábitat, cacería, enfermedad, entre otros (Palacios y Rodríguez, 2001; Chapman y Balcomb, 1998; Crockett, 1998; Crockett, 1996; Schwarzkopf y Rylands, 1989; Rylands y Keuroghlian 1988; Crockett y Eisenberg, 1987; Defler, 1981).
A pesar de ser una especie ampliamente estudiada en tierras bajas, se encuentran pocos estimativos de densidad poblacional de aulladores rojos en bosques montanos. En Colombia, en el Huila, cordillera Central (2300 m de altitud) se calculó una densidad de ca. 15 indv/km2 en un bosque no protegido y poco intervenido (Gaulin y Gaulin, 1982). En Risaralda, en la misma cordillera, a ~1900 m, se reportó una densidad de 73 indv/km2 en un bosque con plantaciones forestales no comerciales y vegetación natural de unos 60 años de recuperación y protección (Gómez-Posada et ál., 2007). La densidad que estimamos en el Cañón del Barbas (132 indv/km2) es superior a estos registros, así como a la mayoría de los reportes de tierras bajas y bosques continuos (Chapman y Balcomb, 1998; Crockett y Eisenberg, 1987) y similar a la reportada en condiciones de aislamiento. Así por ejemplo, en fragmentos aislados de bosque premontano en las cordilleras Central y Occidental de Colombia, fueron estimadas densidades de 377.7 y de 192 indv/km2 respectivamente (Londoño y Gómez-Posada, 2007; Gómez-Posada et ál., 2005). En una pequeña isla en el Lago Gurí en Venezuela fue estimada una densidad para aulladores rojos entre 800 a 1000 indv/km2 (Orihuela-López et ál., 2005). Esta alta densidad de aulladores encontrada en fragmentos aislados como el del Cañón del Barbas podría deberse al aislamiento del bosque y a la reducción del hábitat circundante, que limita sus capacidades de dispersión y los confina en el fragmento (Pruetz y Leasor, 2002; Defler, 1981). Para el género Alouatta, han sido reportadas densidades mayores en bosques aislados comparados con bosques continuos, e incluso se han encontrado fragmentos sobrepoblados (Orihuela-López et ál., 2005; Van Belle y Estrada, 2005; Rodríguez-Toledo et ál., 2003; Estrada y Coates-Estrada, 1996). Gracias a su estrategia de ahorro de energía, sus bajos requerimientos de espacio y su dieta folívora facultativa, los aulladores son reconocidos por su capacidad para sobrevivir en ambientes fragmentados e intervenidos (Mandujano et ál., 2004; Bicca-Marques, 2003; Rodríguez-Toledo et ál., 2003; Estrada et ál., 2002; Estrada y Coates-Estrada, 1996; Rylands y Keuroghlian, 1988; Lovejoy et ál., 1986; Crockett, 1998; Crockett, 1996; Milton, 1980). Así por ejemplo, en México, los aulladores de manto (Alouatta palliata) han persistido en mayor número de fragmentos y con mayor número de individuos, en comparación con monos araña Ateles geoffroyi (Estrada y Coates-Estrada, 1996). En diferentes estudios en dicho país, se han reportado a los aulladores en remanentes boscosos de diferentes tamaños y formas, e incluso en cultivos y bosques perturbados. En general, en los fragmentos la densidad de aulladores fue mayor que en bosques continuos (Mandujano et ál., 2004; Rodríguez-Toledo et ál., 2003; Estrada et ál., 2002; Estrada y Coates-Estrada, 1996).
En el presente trabajo se estimó por extrapolación una abundancia de aulladores de alrededor de 1000 individuos para el Cañón del Barbas. Un número poblacional alto podría mantener la variabilidad genética en el largo plazo y atenuar los efectos negativos de la endogamia y falta de flujo genético por el aislamiento (Robinson y Ramírez, 1982). Sin embargo, una alta densidad de individuos en un bosque aislado puede permitir la rápida propagación de epidemias, estrés, cambios en la estructura demográfica de las poblaciones, entre otros (Van Belle y Estrada, 2005; Crockett, 1998; Estrada y Coates-Estrada, 1996;). El promedio del tamaño de grupo en el Cañón del Barbas tiende a ser alto (11.3 ±2) y se encontraron grupos hasta con siete hembras y cinco machos adultos y subadultos. Este alto número de individuos en capacidad de reproducirse en cada tropa no es común para el tipo de sociedad del aullador rojo (Chapman y Balcomb, 1998; Crockett y Eisenberg, 1987; Crockett, 1996; Soini, 1992; Defler, 1981) y podría indicar indicios de alteraciones de la estructura social como resultado del aislamiento de la población, tal como ha sido reportado para aulladores de manto A. palliata (Van Belle y Estrada, 2005). La reciente figura legal para proteger el bosque del Cañón del Barbas y la creación de corredores de conexión con la reserva Bremen-La Popa, seguramente favorecerán a los aulladores, al ofrecer más hábitat disponible protegido y además opciones de dispersión. Es importante iniciar monitoreos continuos para determinar el efecto de la protección y conexión de los bosques sobre la densidad y composición de grupos de los aulladores.
La cuenca media del Barbas presenta zonas con bosques maduros y secundarios, por tanto en las parcelas de vegetación se encontró un amplio rango de valores para las variables evaluadas. Los promedios encontrados demostraron que en este bosque son comunes los árboles de gran porte, con gran DAP, grandes copas y alturas considerables. El DAP y la altura han sido relacionados como importantes al momento de evaluar el hábitat de los aulladores, ya que este género de animales son principalmente cuadrúpedos de dosel, que utilizan árboles de gran porte y prefieren los estratos superiores del bosque, con buena continuidad de dosel (Mandujano et ál., 2004; Defler, 2003; Stevenson et ál., 2000; Soini, 1992, Yoneda, 1990, Schwarzkopf y Rylands, 1989). Adicionalmente, el Cañón del Barbas presentó abundancia de las principales especies consumidas por los aulladores, particularmente aquellas de reconocida importancia para esta especie en bosques montanos, como las moráceas y las cecropiaceas (Giraldo et ál., 2007; Londoño, 2004; Gaulin y Gaulin 1982). Durante seis meses de seguimiento a dos tropas de aulladores en este bosque, se registraron 37 especies de plantas consumidas (De la Cruz, 2006). Al parecer, en este fragmento relativamente grande y diverso, los aulladores no se encuentran sometidos a estrés dietario, como puede suceder en fragmentos más pequeños (Orihuela et ál., 2005; Neves y Rylands, 1991). Se puede sugerir que el bosque del Cañón del Barbas, ofrece buena disponibilidad de recursos estructurales y alimenticios para los aulladores.
La población de aulladores del Barbas, probablemente fue diezmada durante el siglo XIX y por lo menos ¾ partes del siglo XX durante la fuerte reducción y entresaca de los bosques. Es posible que esta población de monos se encuentre en recuperación tras la veda de la caza y la disminución de la presión antrópica sobre el bosque. Desafortunadamente no se encuentran registros antiguos que permitan determinar las tendencias poblacionales de esta especie en este bosque. En un estudio realizado en 120 ha dentro de este mismo fragmento, se estimó por conteo directo, una densidad de aulladores de 37 indv/km2 (De la Cruz et ál., 2006), menor a la estimada en el presente estudio. La diferencia puede deberse a los diferentes métodos empleados y no permite sugerir que se haya presentado un incremento poblacional.
Los aulladores del Cañón del Barbas se encuentran aislados de otras poblaciones de aulladores. Es importante iniciar procesos de manejo a nivel de paisaje que incrementen la posibilidad de conexión con otras poblaciones cercanas de aulladores; como por ejemplo con la cuenca alta del Barbas. La flexibilidad conductual de estos monos les permite usar variedad de opciones de conexión entre bosques, como cercas vivas, cultivos con sombrío, guaduales, plantaciones forestales, entre otros (Mandujano et ál., 2004; Rodríguez-Toledo et ál., 2003; Estrada y Coates-Estrada, 1996)
AGRADECIMIENTOS
Este trabajo contó con el apoyo de John D. and Catherine T. MacArthur Foundation, Fundación para la Promoción de la Investigación y la Tecnología del Banco de la República (proyecto 1.758) e Idea Wild. Agradecemos a Smurfit Kappa Cartón de Colombia por el apoyo logístico y el permiso para trabajar en sus predios, así como a otros propietarios del bosque. A Marisol González y familia por la hospitalidad, a Felipe Castaño por la identificación del material vegetal, a Julián Andrés Sánchez Pachón por la colaboración en campo, a Leonor Valenzuela por la ayuda con el mapa y la georeferenciación de las fotografías aéreas. Al Doctor Gustavo Kattan por su continuo apoyo y orientación.
BIBLIOGRAFÍA
ACERO L. Árboles de la zona cafetera colombiana. Armenia: Ediciones Fondo Cultural Cafetero; 1985.
[ Links ]BICCA-MARQUES JC. How do the howlers cope with habitat fragmentation? En: Marsh LK, editor. Primates in fragments: ecology and conservation. New York: Kluwer Academic / Plenum Publisher; 2003. p. 283-304.
[ Links ]CHAPMAN C, BALCOMB S. Population characteristics of howlers: ecological conditions or group history. Int J Primatol. 1998; 19: 385-403.
[ Links ]CHAVES M, ARANGO N, editores. Informe nacional sobre el estado de la biodiversidad, Tomo II. Bogotá: Instituto de investigación de Recursos Biológicos Alexander von Humboldt; 1997.
[ Links ]CHIARELLO AG, DE MELO FR. Primate population densities and sizes in Atlantic forest remnants of northern Espirito Santo, Brazil. Int J Primatol. 2001; 22: 379-396.
[ Links ]CINEP [Internet]. Colombia país de regiones. Tomo I. Bogotá DC: Biblioteca Virtual Luís Ángel Arango; 2004. Consultado en febrero de 2006. Disponible en URL: http://www.banrepcultural.org/blaavirtual/geografia/region1/indice.htm.
[ Links ]CROCKETT C. The relation between red howler monkey (Alouatta seniculus) troop size and population growth in two habitats. En: Norconk MA, Rosenberg AL, Garber PA, editors. Adaptative Radiations of Neotropical Primates. New York: Plenum Press; 1996. p. 489-510.
[ Links ]CROCKETT C. Conservation biology of the Genus Alouatta. Int J Primatol. 1998; 9: 549-579.
[ Links ]CROCKETT C, EISENBERG J. Howlers: variations in group size and demography. En: Smuts B, Cheney D, Seyfarth R, Wrangham R, Struhsaker T, editors. Primate Societies. Chicago: The University of Chicago Press; 1987. p. 54-68.
[ Links ]DE LA CRUZ JC. Dispersión de semillas y patrones de defecación del mono aullador rojo (Alouatta seniculus Linneaus 1766) en un relicto de bosque subandino en el Cañón del río Barbas - Quindío [tesis de pregrado]. Bogotá: Carrera de Ecología, Facultad de Estudios ambientales y rurales, Universidad Javeriana; 2006.
[ Links ]DE LA CRUZ JC, HERNÁNDEZ-CHAPARRO RF, QUIMBAYO-RUIZ G. Densidad de Alouatta seniculus en dos relictos de bosque subandino en el Quindío. Ecotono. 2006; 2: 13-17.
[ Links ]DEFLER T. The density of Alouatta seniculus in the eastern llanos of Colombia. Primates. 1981; 22: 564-569.
[ Links ]DEFLER T. Primates de Colombia. Serie de Guías Tropicales de Colombia 4. Bogotá: Conservación Internacional; 2003.
[ Links ]ESTRADA A, COATES-ESTRADA R. Tropical rain forest fragmentation and wild populations of primates at Los Tuxtlas, Mexico. Int J Primatol. 1996; 17: 759-783.
[ Links ]ESTRADA A, MENDOZA A, CASTELLANOS L, PACHECO R, VAN BELLE S, et ál. Population of the black howler monkey (Alouatta pigra) in a fragmented landscape in Palenque, Chiapas, Mexico. Am J Primatol. 2002; 58: 45-55.
[ Links ]ETTER A,VILLA A. Andean forest and farming systems in part of the eastern cordillera (Colombia). Mt Res Dev 2000; 20: 236-243.
[ Links ]FRANCO AM, BRAVO G. Áreas importantes para la conservación de las aves en Colombia. En: BirdLife International and Conservation International, editores. Áreas Importantes para la conservación de las aves en los Andes Tropicales: sitios prioritarios para la conservación de la biodiversidad, Serie de Conservación de Birdlife N° 14. Quito: Birdlife Internacional y Conservation Internacional; 2005. p. 117-281.
[ Links ]GAULIN S, GAULIN C. Behavioral ecology of Alouatta seniculus in Andean cloud forest, Colombia. Int J Primatol. 1982; 3: 1-32.
[ Links ]GIRALDO P, GÓMEZ-POSADA C, MARTÍNEZ J, KATTAN G. Resource use and seed dispersal by red howler monkey (Alouatta seniculus) in Andean Forest. Neotrop Primates 2007; 14: 55-64.
[ Links ]GÓMEZ-POSADA C. Biología y estado de conservación del mono aullador rojo. En: Kattan G, Valderrama C, editores. Plan de conservación del mono aullador rojo (Alouatta seniculus) en la región del Sirap Eje Cafetero y Valle del Cauca. Bogotá DC: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt y WCS Programa Colombia / Fundación EcoAndina; 2006. p. 13-40.
[ Links ]GÓMEZ-POSADA C, MARTÍNEZ J, GIRALDO P, KATTAN G. Density, habitat use and ranging patterns of red howler monkey in Andean forest. Neotrop Primates. 2007; 14(1): 2-10.
[ Links ]GÓMEZ POSADA C, RONCANCIO N, HINCAPIÉ P. Evaluación de las poblaciones de mono aullador rojo (Alouatta seniculus) en fragmentos de bosque en el Valle del Cauca. Informe presentado a la Corporación Regional del Valle del Cauca CVC. Cali: WCS Colombia / Fundación EcoAndina; 2005.
[ Links ]HERNÁNDEZ-CAMACHO J, COOPER R. The nonhuman primates of Colombia. En: Thorington R, Heltne PH, editors. Neotropical primates: field studies and conservation. Washington DC: National Academy of Sciences; 1976. p. 35-69.
[ Links ]KATTAN G, ÁLVAREZ-LÓPEZ H. Preservation and management of biodiversity in fragmented landscape in the Colombian Andes. En: Schelhas J, Greenberg R, editors. Forest patches in tropical landscape. Washington DC: Island Press; 1996. p. 3-18.
[ Links ]LONDOÑO JM. Dieta del mono aullador (Alouatta seniculus) en la reserva la Montaña del Ocaso, Quimbaya, Quindío [tesis de pregrado]. Popayán: Programa de Ecología, Facultad de Ciencias, Fundación Universitaria de Popayán; 2004.
[ Links ]LONDOÑO JM, GÓMEZ-POSADA C. Evaluación del estado de la población del mono aullador rojo (Alouatta seniculus) en la reserva La Montaña del Ocaso. Informe presentado al CIBUQ, Universidad del Quindío. Cali: Fundación EcoAndina / WCS Colombia; 2007.
[ Links ]LOVEJOY TE, BIERREGAARD Jr RO, RYLANDS A, MALCOLM JR, QUINTELA CE, et ál. Edge and other effects of isolation on Amazon forest fragments. En: Soulé ME, editor. Conservation Biology: The science of scarcity and diversity. Suderland: Sinauer Associates; 1986. p. 257-285.
[ Links ]MANDUJANO S, ESCOBEDO-MORALES LA, PALACIOS-SILVA R. Movements of Alouatta palliata among forest fragments in Los Tuxtlas, México. Neotrop Primates. 2004; 12 (3): 126-131.
[ Links ]MILTON K. The foraging strategy of howler monkeys. A study in primate economics. New York: Columbia University Press; 1980.
[ Links ]NATIONAL RESEARCH COUNCIL. Techniques for the study of primate population ecology. Washington DC: National Academy Press; 1981.
[ Links ]NEVES AM, RYLANDS A. Diet of a group of howling monkeys, Alouatta seniculus, in an isolated forest patch in Central Amazonia. Acta Primatologica. 1991; 3: 263-274.
[ Links ]ORIHUELA-LÓPEZ G, TERBORGH J, CEBALLOS N. Food selection by a hyperdense population of red howler monkeys (Alouatta seniculus). J Trop Ecol. 2005; 21: 445-450.
[ Links ]PALACIOS E, RODRIGUEZ A. Ranging pattern and use of space in a group of red howler monkeys (Alouatta seniculus) in a southeastern Colombian rainforest. Am J Primatol. 2001; 55: 233-251.
[ Links ]PERES C. General guidelines for standardizing line-transect surveys of tropical forest primates. Neotrop Primates. 1999; 7: 11-16.
[ Links ]PRUETZ JD, LEASOR HC. Survey of three primate species in forest fragments at La Suerte Biological Field Station, Costa Rica. Neotrop Primates. 2002; 10: 4-9.
[ Links ]ROBINSON J, RAMÍREZ J. Conservation biology of Neotropical primates. En: Mares MA, Genoways HH, editors. Mammalian Biology in South America. Pittsburgh: University of Pittsburgh Press; 1982. p. 329-344.
[ Links ]RODRÍGUEZ-TOLEDO EM, MANDUJANO S, GARCÍA-ORDUÑA F. Relationships between forest fragments and howler monkeys (Alouatta palliata mexicana) in southern Veracruz, México. En: Marsh LK, editor. Primates in fragments: ecology and conservation. New York: Kluwer Academic / Plenum Publisher; 2003. p. 79-97.
[ Links ]RYLANDS A, KEUROGHLIAN A. Primate populations in continuous forest and forest fragments in Central Amazonia. Acta Amazonica. 1988; 18 (3-4): 291-307.
[ Links ]SCHWARZKOPF L, RYLANDS A. Primate species richness in relation to habitat structure in Amazonian rainforest fragments. Biol Conserv. 1989; 48: 1-12.
[ Links ]SOINI P. Ecología del coto mono (Alouatta seniculus, CEBIDAE) en el río Pacaya, reserva Pacaya - Saimiria, Perú. Folia Amazonica. 1992; 4: 103-118.
[ Links ]STEVENSON P, QUIÑONES M, AHUMADA J. Influence of fruit availability on ecological overlap among four neotropical primates at Tinigua National Park, Colombia. Biotropica. 2000; 32 (3): 533-544.
[ Links ]VAN BELLE S, ESTRADA A. Cambios demográficos en poblaciones del mono aullador negro (Alouatta pigra) como consecuencia de la fragmentación del hábitat. Universidad y Ciencia 2005; Número especial 002: 1-9.
[ Links ]YONEDA M. The difference of tree size used by five cebid monkeys in Macarena, Colombia. Field Stud. New World Monkeys, La Macarena Colombia. 1990; 3: 13-18.
[ Links ]
