










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.15 no.1 Bogotá Jan./Apr. 2010
ACEPTACIÓN DE UNA DIETA ARTIFICIAL POR LARVAS DE LA MARIPOSA BATTUS POLYDAMAS POLYDAMAS(LEPIDOPTERA: PAPILIONIDAE)
ACCEPTANCE OF AN ARTIFICIAL DIET FOR CATERPILLARS OF THE BUTTERFLY BATTUS POLYDAMAS POLYDAMAS (LEPIDOPTERA: PAPILIONIDAE)
Ricardo A. Claro 1, MsC; Natalia Ruiz1, PhD. 1Departamento de Biología, Universidad Nacional de Colombia AA 14490 Bogotá, Colombia. raclaroc@unal.edu.co ; nruizr@unal.edu.co
Presentado 18 de febrero de 2008, aceptado 1 de Julio de 2009, correcciones 1 de julio de 2009
RESUMEN
Se evalúo la aceptabilidad de dietas artificiales para la alimentación de larvas de la mariposa Battus polydamas polydamas (Lepidoptera: Papilionidae), bajo condiciones de laboratorio. A las larvas se les ofrecieron dietas en diferentes presentaciones y composiciones: en discos, vertida como suplemento sobre hojas del hospedero y plantas hospedero como control. La composición de la dieta se realizó de acuerdo con ensayos previos y con los análisis nutricionales desarrollados a larvas maduras y a hojas de la planta hospedera, Aristolochia maxima (Aristolochiaceae). La longevidad de las larvas, criadas en laboratorio, se vio afectada significativamente por los diferentes tratamientos de alimentación. Solo se finalizó el ciclo de desarrollo entre las larvas alimentadas con hojas jóvenes de la planta (control) y con dieta esparcida como suplemento sobre hojas del hospedero; las curvas de sobrevivencia no presentaron diferencias significativas entre las larvas sostenidas con estos dos tratamientos. En contraste, las tasas de crecimiento larval, fueron significativamente afectadas según el tratamiento, obteniéndose mayor biomasa en las alimentadas con hojas con suplemento. Se sustenta la cría de mariposas de la especie B. polydamas bajo una dieta artificial como suplemento esparcido sobre hojas del hospedero bajo condiciones de temperatura, humedad y luminosidad controlada, con propósitos de investigación o futuros planes de manejo y conservación.
Palabras claves: Aristolochia máxima, Oligofagos, Historias de Vida, Cría, Conservación.
ABSTRACT
We evaluated the acceptability of an artificial diet for the feeding larval stage of the butterfly Battus polydamas polydamas (Lepidoptera: Papilionidae) under control conditions. We offered to the larvae a diet in different forms and composition: disks, spilled like a supplement over the host sheet plant and host plant like a control. The diet composition was made according with preliminary essays and nutritional analyses to the larvae and the host plant, Aristolochia maxima. The longevity of the larvae, raised in the laboratory, was significantly affected for the different food treatments. Only finalized the life cycle of the larvae feed with young host plant sheet (control) and with diet spilled like a supplement over the host plant sheet. The surviving curves do not show significant differences between the larvae feeding with young leaves and the larvae feeding with spread diet. In contrast, the larva growth rates were significantly affected by the treatment. We found that the breeding of B. polydamas with an artificial diet is possible, under control conditions of temperature, humidity and light, for a research proposal or for future handling and conservation planes.
Key words: Aristolochia maxima, Oligophagous, Life History, Breeding, Conservation.
INTRODUCCIÓN
La herbivoría, en los lepidópteros está mediada por diferentes factores como la escogencia del sitio de oviposición por parte de la hembra, el comportamiento de la progenie, la distribución geográfica de las especies, los nutrientes de la planta y en su gran mayoría por la composición química de las plantas (Da Silva 2005; Jaenike 1990). En relación con la estructura química, los Papilionidae se estudian ampliamente, especialmente la tribu Troidini (Papilioninae) cuya interacción con las plantas del genero Aristolochia (Aristolochiaceae) se ha utilizado como ejemplo de coevolución entre insectos herbívoros y sus plantas hospederas (Da Silva 2005). La estrecha relación se le atribuye a características de los compuestos químicos de la planta, que sirven como fagoestimulantes para que las larvas de esta tribu de mariposas se alimenten específicamente de ese género de plantas (Da Silva 2005; Berenbaum 1995;). Estos compuestos químicos, son retenidos y mantenidos hasta la fase adulta, para protegerlas de los ataques de los depredadores y parasitoides (Sime 2000) y advierten el sabor desagradable de las mariposas (Fordyce 2001). Algunos compuestos responsables de esa actividad son los ácidos aristolóquicos, las aristolactamas, los alcaloides bencilisoquinolinos y los terpenoides (Silva-Brandâo et ál. 2006; Sime et ál. 2000; Klitzke y Brown 2000; Chai 1986; Mix et ál. 1983).
Adicionalmente, para los Troidini, la planta hospedero es su principal recurso y les aporta los compuestos nutricionales necesarios para el crecimiento, desarrollo y defensa en cada una de las etapas de su ciclo de desarrollo. Desde la formación del huevo, el almacenamiento e intercambio de nutrientes adquiridos y depositados en la alimentación adulta (O´brien 2004; Boggs 1997a, 1997b, 1993), sirven como primer recurso de nutrientes para la etapa larval (Barros 2006). En la fase larval se alimenta de plantas para almacenar y distribuir compuestos destinados al crecimiento, almacenamiento y sobrevivencia (Telang et ál. 2001; Scriber y Slansky 1981), que suministran la energía y nutrientes implicados en las actividades reproductivas, de forrajeo y sobrevivencia en la etapa adulta (Fischeret ál. 2004; Marshall 1982; Boggs 1981). De igual forma, estas mariposas hacen parte de los siguientes eslabones de la cadena alimenticia, convirtiéndose en un recurso para otros consumidores (Lee 1995; Chai 1986). En otras palabras, la manera como estos lepidópteros asignen los recursos en los componentes integrales de su historia de vida como el crecimiento, el desarrollo, el almacenamiento, la reproducción y la defensa, determinará su valor adaptativo y el tamaño de la población en condiciones naturales.
En contraposición, la extraordinaria belleza por sus coloridos y formas, desarrolló una industria entre coleccionistas y comercializadores (Parsons 1995; Garrawayet ál. 1993; Parsons 1992;), que sumado a la destrucción directa del hábitat, amenaza enormemente la sobrevivencia de estas mariposas. Por lo tanto, mecanismos de conservación in-situ, complementado con la cría en cautiverio, pueden contribuir con la preservación de estas mariposas y sus recursos (Newet ál. 1995). Aunque, para las mariposas de la familia Papilionidae se trazaron planes de conservación y poseen su propio libro rojo por su alto grado de amenaza (Tyler et ál. 1994), poco esfuerzo se dedica al desarrollo de medios de cría artificial (Siyuan 1999; Morton 1979; Singh 1977) a pesar de estar las técnicas bien desarrolladas para la cría en masa de polillas polífagas, utilizando su alimento natural o dietas artificiales, con propósitos económicos (Genc y Nation 2004; Mattoni et ál. 2003; Cohen 2001).
Ante la especificidad de la relación Troidini-Aristolochia, mediada por los ácidos aristolóquicos y la importancia de la dieta en los componentes de la historia de vida del organismo, nos planteamos las siguientes preguntas: 1) ¿Es posible criar estas mariposas oligófagas bajo una dieta artificial? y 2) ¿Cuál sería el efecto en los rasgos de historia de vida de la larva? Por consiguiente, en este trabajo evaluamos la aceptabilidad de una dieta artificial y su efecto en la sobrevivencia, el crecimiento y la longevidad de larvas de Battus polydamas polydamas (Linnaeus, 1758), para un mejor entendimiento de la ecología química de la interacción, que permita un avance en técnicas de cría de mariposas aplicables en trabajos de investigación y en programas de manejo y conservación de mariposas.
MATERIALES Y MÉTODOS
MARIPOSAS Y PLANTA HOSPEDERA
El género Battus está representado en Colombia por siete especies y ocho subespecies que habitan principalmente las zonas cálidas de bosque húmedo, montano y seco tropical del país (Le Crom et ál 2002). Se caracterizan por ser mariposas de tamaño mediano a grande, de coloración negra con bandas y manchas amarillas, los adultos presentan un vuelo rápido en sitios abiertos y orillas del bosque (Le Crom et ál 2002). Las larvas son de color café y negro con una serie de tubérculos en el cuerpo. Las mariposas de la especie B.polydamas en estado adulto se caracterizan por el color negro intenso de sus alas anteriores y posteriores, marcadas con una banda vertical angosta submarginal de color amarillo (Le Crom et ál 2002). Los huevos son de color amarillo con diez estrías corrugadas longitudinalmente y colocados en grupos pequeños de hasta dieciséis (Fagua et ál. 1998); en este estado de huevo se mantienen aproximadamente nueve días. Las larvas son poligófagas, especficamente del género de plantas Aristolochia, de hábitos gregarios con una duración total del estado de 20,66 días (Fagua & Ruiz 1996 y Fagua et ál. 1998). Hasta el tercer estadio, presentan polimorfismo y ya maduras tienen cuerpo de color café claro con amarillo, moteado con finas estrías obscuras, tubérculos carnosos de color marrón con un par de tubérculos largos móviles en el protórax. Al igual que toda la familia de Papilionidae, se reconocen también por la presencia del osmeterium, de coloración amarillo-anaranjado intenso, que actúa como mecanismo de defensa para disuadir el ataque de los depredadores. La pupa, que presenta dos coloraciones, café con dorso anaranjado o verde con dorso amarillo, contiene un cuerno toráxico de longitud mediana proyectado hacia atrás y permanece por lo menos 18 días en este estado (Le Crom et ál. 2002; Constantino 1997; Fagua y Ruiz 1996).
Los huevos de B. polydamas, utilizados en el estudio fueron encontrados sobre Aristolochia maxima (Jacq, 1760), se coleccionaron en el municipio de Nilo, departamento de Cundinamarca, Colombia, a una altitud media de 336 m, con una temperatura media anual de 28 ºC, una humedad relativa promedio de 72%, una precipitación media anual de 1380 mm y un valor de brillo solar de 2169 horas promedio anual (IDEAM 2006), en una formación vegetal, claramente definida según el sistema de zonas de vida, cómo Bosque Seco Tropical.
ENSAYOS PRELIMINARES
Para los tratamientos con dietas artificiales se ensayó primero una mezcla de la dieta reportada por Morton (1979), que ha servido para la cría exitosa de varias especies de mariposas (Genc y Nation 2004; Mattoni et ál. 2003; Siyuan 1999; Ellis 1998; Singh 1977;). Sin embargo, no hubo evidencia alguna de que las larvas probaran la dieta. Posteriormente, se aumentó la proporción de material de la planta seca y pulverizada dentro de la dieta a un 2%, 5%, 10% y el resultado fue el mismo, sobrevivencia larval entre siete y diez días y ninguna muestra de dieta consumida.
EXPERIMENTOS DE CRÍA
Los tratamientos utilizados en el desarrollo de los experimentos, fueron:
1. Tratamiento uno (control); larvas alimentadas con alimento natural: en el interior de bolsas plásticas de 20 X 25 cm con sellado hermético de la parte superior, infladas con aire de compresor, fueron colocadas hojas aún prendidas al tallo de la parte apical de las plantas de A. maxima. Sobre las hojas se puso la larva que se alimento at livitum y se peso cada vez que las hojas se marchitaban y era necesario el cambio de alimento.
2. Tratamiento dos; larvas alimentadas con discos de dieta artificial: material de la dieta (Tabla 1), previamente preparada y refrigerada, fue perforado con un sacabocados de corcho, de tal manera que se formaban discos de diámetro de 8 mm y un ancho de 2 mm, estos discos se colocaron dentro de recipientes plásticos de 2 onzas cuyo fondo contenía una circunferencia del mismo diámetro del recipiente en papel filtro. Al interior fue colocada una larva recién eclosionada y el recipiente se cubrió con una tapa plástica de pequeños orificios en la superficie elaborados con alfiler, y fue dejado en observación dentro de la cámara de cría. Los discos se cambiaron a medida que mostraban resequedad por evaporación.
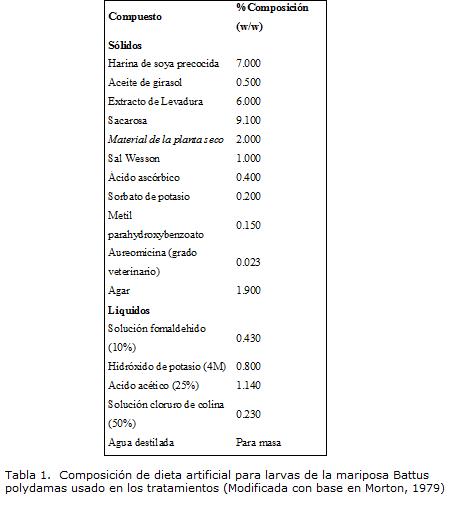
3. Tratamiento tres; larvas alimentadas con dieta artificial vertida: mezcla de la dieta (Tabla 1), previo a su endurecimiento, es decir, en caliente, fue vertida en el fondo de vasos plásticos de 2 onzas, que se dejaron secar por unas horas y posteriormente se deposito la larva, para seguimiento en la cámara de cría, el alimento se cambio de acuerdo al secado que presentara la dieta al pasar de los días y permaneció cerrada con tapas plásticas iguales al tratamiento anterior.
4. Tratamiento cuatro; larvas alimentadas con dieta artificial esparcida en hoja: se esparce mezcla de la dieta artificial, sin material de la planta, con un pincel, sobre el envés de hojas frescas de A. maxima, y, colocándolas en potes plásticos de 18 onzas, protegidos con tapas plásticas, se colocaron larvas de B. polydamas. Las hojas fueron cambiadas de acuerdo al consumo por parte de las larvas y al deterioro de la misma.
Los experimentos se realizaron en un cuarto de cría con condiciones controladas de temperatura 27,7ºC ± 1,6, humedad 59,6%± 4,7 y fotoperíodo de 12 horas luz, donde se mantenían los huevos hasta su eclosión para posteriormente trasladar las larvas a los tratamientos. Por otro lado, en el interior de la misma cámara, para aquellos animales que llegaron a estado adulto, se construyó una jaula de 1 metro cúbico, cubierta y sellada con tela tull para toldillo, dejando una abertura para la manipulación de animales y plantas. En su interior se mantenían dos masetas cada una con una planta joven de A. maxima, con la intención de, si se lograba apareamiento, que las hembras tuvieran lugar de postura.
Para la evaluación de cada uno de los tratamientos, se midieron variables de sobrevivencia, longevidad y crecimiento en etapa larval de los individuos de B. polydamas, colectados desde el estado de huevo, de la siguiente forma:
- El seguimiento en el tiempo del grupo de individuos en etapa larval se desarrollo con la construcción de tablas de vida de cohorte, empleando el programa Excel (Microsoft® Excel, XP).
- La longevidad se determinó por el número de días que duró el ciclo, que se consignó previamente en la tabla de vida.
- La tasa de crecimiento, se evaluó como el cambio de peso en un tiempo específico, establecido por la masa final menos la masa inicial dividido el número de días que duró la etapa de alimentación larval.
DIETA ARTIFICIAL
Después de los ensayos realizados, se decidió utilizar una dieta con aceite de girasol en una proporción del 0.5%, para una mejor y pronta disponibilidad de ácidos grasos para la larva y aumentar al 9.1% la sacarosa por aporte de carbohidratos solubles (Tabla 1).
Para la mezcla y elaboración de la dieta, los materiales sólidos finamente granulados junto al material de la planta, sometido a secado durante 48 horas en un horno de aire forzado a una temperatura de 60º C y después molido finamente en un molino de cuchillas con una criba de 1mm, fueron mezclados y homogenizados. Posteriormente, junto a una mixtura de agar con agua destilada en caliente se vierte rápidamente el material solidó y a continuación se adicionan los componentes líquidos de la dieta. Esta preparación se esparció en cajas petri o en recipientes plásticos de 2 onzas dejando secar a temperatura ambiente para su consecutiva utilización.
Para la preparación de la dieta utilizada en el tratamiento cuatro no se incluía material de la planta una vez que la forma de utilizar la dieta fue esparciéndola sobre la superficie de las hojas que se colocaron como alimento para las larvas.
CONTENIDOS NUTRICIONALES
Para las larvas y la planta, se desarrollaron análisis químicos para determinar contenidos nutricionales de materia seca, proteína cruda (Nitrogeno x 6.25), extracto etéreo que estima el contenido de aceites y grasas presentes, cenizas que indica la proporción de los minerales en forma global y Extracto Libre de Nitrogeno que estima los glúcidos como almidones, azúcares solubles, pectinas y ácidos orgánicos, fácilmente digeribles por un consumidor (A.O.A.C. 1984). Para las hojas de la planta, adicional a los análisis anteriores, se estimó el contenido de los constituyentes de la pared celular, que son la mayor fracción de la planta y, la menor y más lentamente digerida en el tracto digestivo de los herbívoros: fibra cruda la cual estima la celulosa y lignina, generalmente indigeribles por algunos animales (A.O.A.C. 1984), fibra en detergente acido (FDA) que evalúa contenidos de lignina, celulosa y minerales insolubles y fibra en detergente neutro (FDN) que es una medida más precisa de los contenidos de lignina, celulosa y hemicelulosa. El 100% menos el valor de la FDN representa los nutrientes más aprovechables que se encuentran encerrados por la pared celular (contenido celular), esta fracción incluye proteína y carbohidratos solubles, minerales solubles y lípidos, por lo tanto un alto porcentaje de contenido celular es indicativo de alto valor nutritivo de la planta (Haufler y Servello 1994; Nagy 1987; Van Soest 1982). Suplementariamente, para determinar y corroborar la presencia de los compuestos químicos responsables de la interacción entre A. maxima y B. polydamas, se corrieron análisis de Cromatografía Líquida de Alta Eficiencia, CLAE (Claro et ál.2007).
ANÁLISIS ESTADÍSTICO
Con STATGRAPHICS Plus 5.0, se analizaron:
- La longevidad, expresada como el número de días que permanecieron los individuos en estado larval hasta alcanzar el estado de pupa en los diferentes tratamientos, utilizando la prueba no paramétrica Kruskal-Wallis, donde cada larva correspondió a una repetición, utilizando 23 larvas para el tratamiento uno, 11 para el tratamiento dos, 11 para el tratamiento tres y 15 para el tratamiento cuatro, para un tamaño total de muestra de 60 larvas.
- La sobrevivencia, utilizando la prueba estadística no paramétrica Longrank, para comparar si las curvas de sobrevivencia en estado larval de B. polydamas obtenidas de las tablas de vida para cada tratamiento son la misma, en el que cada larva correspondió a una repetición, disponiendo 23 larvas para el tratamiento uno, 11 para el tratamiento dos, 11 para el tratamiento tres y 15 para el tratamiento cuatro, obteniendo un tamaño total de muestra de 60 larvas.
- Las tasas de crecimiento (g/día), en etapa larval de los individuos de B. polydamas para los tratamientos que presentaron sobrevivencia, con un análisis de varianza de una vía, en el cual cada larva correspondió a una repetición, 16 larvas para el tratamiento uno y 12 para el tratamiento cuatro, con un tamaño total de muestra de 28 larvas correspondientes.
RESULTADOS Y DISCUSIÓN
OBSERVACIONES Y COLECTAS EN CAMPO
Las posturas, en su mayoría contuvieron grupos de más de cinco huevos, ubicados preferiblemente en la parte apical y predominantemente en el tallo y pecíolo, de las plantas de Aristolochia maxima. Alrededor del 50% del total de huevos colectados presentaron parasitismo por parte de una especie de Hymenoptera de la familia Braconidae, y aproximadamente el 20% del total de los huevos presentó infertilidad. Así mismo, en condiciones naturales la preferencia alimenticia de los estados larvales, fue por hojas jóvenes de la parte apical de la planta, sin embargo, algunas larvas maduras fueron observadas alimentándose de hojas con mayor grado de dureza, consecuencia de su tolerancia a la asimilación de tejido esclerótico con el aumento del tamaño de la oruga. Estos comportamientos de oviposición y alimentación larval son predominantes en este género de papilios (Hall y Buttler 1998; Fagua y Ruiz 1996; Brown et ál. 1980).
Ése aglomerado larval puede ser una consecuencia de la estrategia de oviposición, ya que la postura de huevos en masa puede reducir el efecto del parasitismo y depredación, además de hacer los huevos menos vulnerables a la deshidratación,; al mismo tiempo, es ventajoso por que incrementa la habilidad de termorregulación de las larvas, disminuye el riesgo de depredación e incrementa la eficiencia de forrajeo, sin embargo, el costo de este comportamiento puede verse expresado en una competencia intraespecíficas cuando el material de la hoja es limitado (Hochuli 2001).
CONTENIDOS NUTRICIONALES
Los análisis de laboratorio para estimar el contenido nutricional de la planta, se desarrollaron para hojas jóvenes y también para hojas tomadas al azar (Tabla 2).
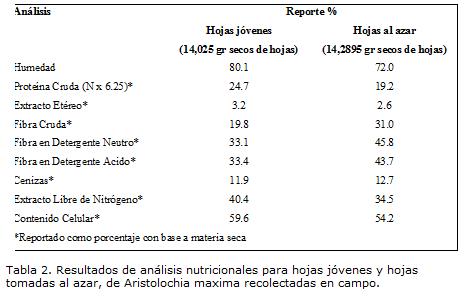
El contenido de agua en hojas jóvenes, fue superior que en hojas tomadas al azar, al igual que la diferencia en la proporción de nutrientes en materia seca, como proteína, grasa, carbohidratos y contenidos celulares, 5.5%, 0.6%, 5.9% y 5.4%, respectivamente, evidenciando una mejor calidad nutricional aportada por hojas jóvenes a los animales en sus primeros instares, además de ser un material 11.2% menos fibroso haciéndolo más digerible y aprovechable por parte de las larvas.
Si se tienen en cuenta los resultados obtenidos, es claro que el comportamiento larval de B. polydamas observado en campo, responde a las diferencias en contenidos nutricionales entre unas hojas y otras. Además, el crecimiento larval, implica el cambio morfológico de la cabeza y la mandíbula en cada muda, ofreciéndole a la larva estrategias anatómicas en su vida alimenticia. La consecuencia de este cambio es la reducida eficiencia de asimilación de recursos en los últimos instares, probablemente como resultado de ingestión de alta proporción de comida de baja calidad, mientras que larvas jóvenes consumen selectivamente proporciones de la hoja que son bajas en fibra, fácilmente digeribles y con una baja proporción de materia seca, ofreciéndole un mejor recurso en crecimiento y desarrollo (Avila et ál 2003; Hochuli 2001; Scriber y Feeny 1979).
En el caso de las larvas, el contenido de nutrientes fue estimado para individuos con un peso entre 1.00 y 1.30 g en los últimos instares, criadas en cautiverio bajo las mismas condiciones enunciadas para el tratamiento uno (Tabla 3).
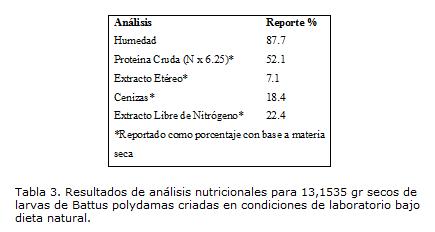
Los resultados dejan ver compuestos nutricionales acumulados en el periodo larval que estarán a disposición para los rasgos de la historia de vida, como la reproducción, sobrevivencia y crecimiento, recursos que provienen del aporte de las hojas jóvenes consumidas en el estado larval, entre los que se encuentran: el nitrógeno como importante recurso para la producción de huevos, constituyente para el tejido de crecimiento y aporte de nitrógeno para los procesos metabólicos; grasas y aceites para el aporte energético, para la síntesis de hormonas, producción de huevos y espermatozoides en estado adulto, esclerotización de la cutícula y como papel metabólico y antinfeccioso; y minerales, que aunque en cantidades traza, son necesarios para el crecimiento y desenvolvimiento así como para el intercambio iónico de las membranas (Panizzi et ál. 1991). En cuanto a los carbohidratos, que al contrario de los compuestos anteriores, no presentan un acumulo dentro de la larva y cumplen funciones como fuente de energía, fagoestimulantes y en muchos casos son necesarios para el crecimiento en este estado, son fácilmente tomados y asimilados del néctar como fuente nutricia para los adultos, donde juegan un papel importante en la longevidad y fecundidad y quizás por esa razón en estado larval la acumulación no es tan alta como con los otros compuestos (Fischer et ál.2004).
DIETA ARTIFICIAL
Sabiendo que la cantidad y la disponibilidad de nutrientes afectan las reservas útiles para el desarrollo y crecimiento de un organismo y a su vez la fecundidad, el número y el éxito de la progenie (Fischeret ál. 2004; Marshall 1982; Boggs 1981, 1992), las dietas elaboradas para los tratamientos dos y tres (Tabla 2), aparentemente aportan las proporciones necesarias para el buen desempeño larval. Sin embargo, no fue del gusto de los animales, posiblemente por factores como los enunciados por Singh (1977) que actúan por si solos o en conjunto, como:
- La textura de la dieta, que probablemente no permitió que las larvas pudieran presionar sus partes bucales por falta de una superficie firme.
- El contenido de agua de la dieta, que de acuerdo a la cantidad en la mezcla para preparación, algunas larvas recién eclosionadas no se podían movilizar fácilmente y se ahogaron.
- La forma de la dieta, que al quedar una superficie lisa al momento de la preparación y observando que las hojas poseen un plano que definitivamente no es uniforme, se convierte en un aspecto físico de relevancia en la movilidad de la larva y en la aceptación de la dieta.
- La deficiencia en fagoestimulantes esenciales, que a pesar de haber determinado la presencia de ácidos aristolóquicos I y II en las hojas jóvenes de A. maxima, cabe la posibilidad de que la planta contenga compuestos volátiles que se añadan a la inducción de estimulación alimenticia.
SOBREVIVENCIA LARVAL
No se encontraron diferencias significativas en el patrón de sobrevivencia entre las larvas de los tratamientos uno y cuatro(X2= 1.62; gl=1; p= 0,2024; Fig. 1).
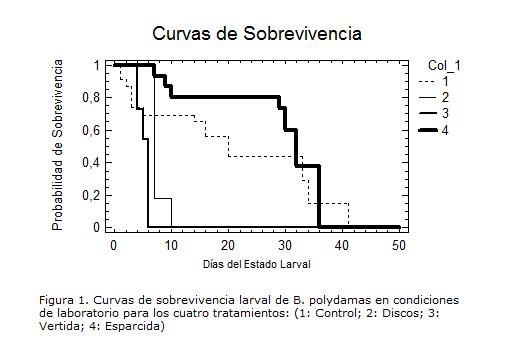
No obstante, las larvas inmaduras del tratamiento dos y tres no probaron alimento artificial, sobrevivieron máximo entre seis y diez días, quizás por el aporte de nutrientes adquirido y almacenado del huevo como primer alimento, pues el corion y el vitelo, capa exterior e interior del huevo, respectivamente, son un importante reservorio de proteínas, lipoproteínas, compuestos carbonados y otros nutrientes (Barros 2006; O´brien et ál 2004.).
LONGEVIDAD DEL ESTADO LARVAL
El número de días que permanecieron los individuos de B. polydamas criados en laboratorio, en estado larval hasta alcanzar el estado de pupa, se vio afectado por los diferentes tratamientos de alimentación aplicados (Kruskall-Wallis=26.0951; gl=3; p<0.05). Las larvas que más demoraron en alcanzar el estado de pupa fueron aquellas que se alimentaron con dieta artificial, esparcida en el envés de hojas jóvenes de A. maxima, con una diferencia en promedio de diez días, comparada con las que se alimentaron de solamente hojas jóvenes de la planta (Tabla 4). Sin embargo, estas larvas presentaron mayor variabilidad entre los días empleados para alcanzar el estado de pupa, mientras que entre larvas del tratamiento cuatro, el tiempo promedio para llegar a ese estado fue más uniforme, sugiriendo que los días necesarios para alcanzar el estado de pupa dependen del aporte de los compuestos nutricionales del alimento, una vez que la dieta es más homogénea en su contenido, contrastada con las hojas que individualmente pueden variar en contenido nutricional, prolongado el tiempo hasta satisfacer las demandas de recursos para almacenamiento y sobrevivencia.
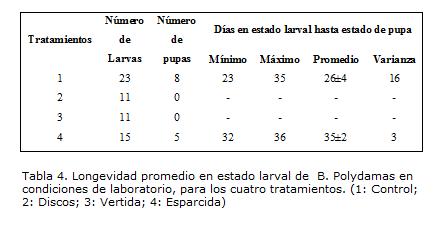
La longevidad de los individuos en estado larval presentó una duración en días más prolongado que los registrados por Fagua et ál. (1998), quienes afirman que la duración como larva es de 20,66 días y para el estado de pupa de 21,66 días. Situación que adicionalmente a lo explicado anteriormente, quizás también este determinada por la diferencia en la humedad mantenida en condiciones de laboratorio, que para el caso de estos ensayos fue de 59,6%± 4,7, mientras que Fagua et ál. (1998) utilizaron una humedad relativa del 75%. Para el estado adulto se considera que el tipo de evaluación debería someterse a condiciones de alojamiento adecuadas para el animal, ya que aunque la temperatura, humedad y luminosidad fueron aceptadas por las mariposas que duraron seis días, no tenían el suficiente espacio para comportamientos etológicos propios de este estado donde reportes como el de Brown et ál (1980) muestran una longevidad en cautiverio de hasta dos semanas.
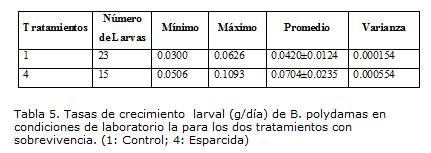
TASAS DE CRECIMIENTO LARVAL
Las tasas de crecimiento larval en Battus polydamas, fueron significativamente afectadas por la dieta (F0.05(1,11)=8.29; p=0.015). En larvas alimentadas con dieta esparcida sobre el envés de la hoja la ganancia de peso (g/día) fue 1.7 veces más alta que las alimentadas con hojas de A. maxima, aunque más variables en la ganancia de peso diaria (Tabla 5). Este resultado puede estar reflejando un mayor almacenamiento de compuestos nutricionales, expresado en el crecimiento, como consecuencia del aumento en contenidos nutricionales aportados por la dieta artificial, que se suman a los recursos propios de las hojas jóvenes de A. maxima.
Adicionalmente, se obtuvieron las curvas de peso en periodo larval para los animales que completaron el ciclo hasta el estado de pupa en los tratamientos uno y cuatro (Fig. 2A-2B).
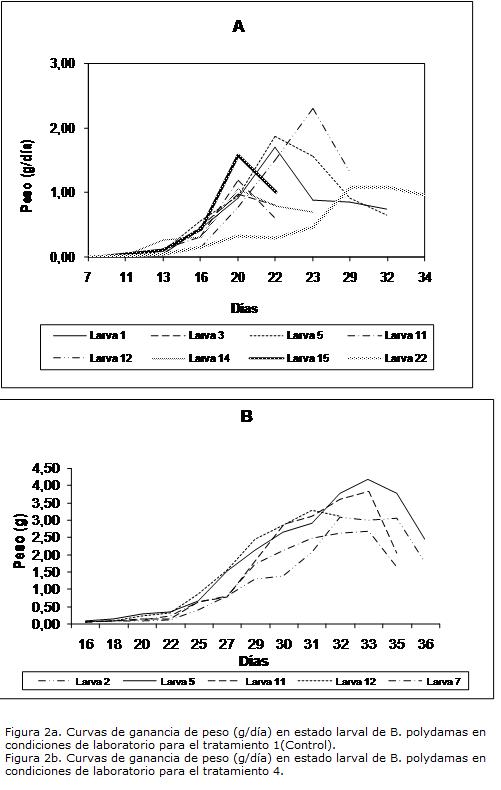
Las curvas de peso dejan ver que, como consecuencia de los cambios morfológicos por crecimiento larva, en los últimos días de la etapa, hay un declive abrupto en la curva después de su etapa máxima de crecimiento. La larva no es lo suficientemente selectiva por su alimento y tiene más tolerancia a tejido esclerótico que ofrecen menos nutrientes, razón que explica el mayor acúmulo de nutrientes en los primeros instar y posterior disminución de peso por menos ingesta de material nutritivo, pues el consumo en los últimos instares incluye más fibra que hace un alto constituyente del material excretado. El punto donde está al 100% de su peso, representa, de acuerdo a Raubenheimer y Simpson (1998), una alta cantidad de nutrientes disponibles y de transferencia para el estado adulto, mientras que las reservas adquiridas de su primer alimento, como es el corion del huevo, son utilizados para sobrevivencia y crecimiento. El punto final de la curva, muestra el peso donde está la máxima eficiencia con la cual los nutrientes son convertidos en una forma química, en la cual son ofrecidos para entrar al estado de pupa. Sin embargo, el peso final de las larvas está significativamente afectado por el tratamiento alimenticio aplicado (F0.05(1,11)=35.13; p<0.05), con un peso promedio para el tratamiento uno de 0.85±0.24 y para el tratamiento cuatro de 2.21±0.6. con una alta variabilidad en larva alimentadas con dieta artificial esparcida sobre hojas jóvenes. Lógicamente, estos pesos son determinados por las tasas de crecimiento larvales, a mayor ganancia de peso mayor será el peso final para entrar al siguiente estado.
Las curvas de peso, sugieren un comportamiento de maximización en las tasas de crecimiento larval de B. polydamas para el tratamiento cuatro, demorándose más tiempo en ser eficientes en el aprovechamiento de los recursos ofrecidos en la dieta. Las larvas con el tratamiento 4 no necesitan ser tan eficientes, ya que cuentan con el suplemento de nutrientes de la dieta artificial, más el aporte natural de hojas jóvenes de A. maximainsinuando que existe, un punto mínimo y un punto óptimo en la eficiencia de toma de recursos. El mínimo, está representado por aquellas larvas que se alimentan de hojas jóvenes de la planta, que en menos tiempo deberían ser más eficientes en la utilización de los recursos disminuyendo la tasa de crecimiento, mientras que las larvas que se alimentan suplementariamente con dieta artificial se demoran más días y aumentan la tasa de crecimiento sin necesidad de aumentar la eficiencia en la utilización del recurso. Ese comportamiento se ve expresado en un mayor tamaño del estado adulto, donde se observó que al comparar, una hembra capturada del medio natural, otra alimentada con hojas de la planta y una tercera alimentada con dieta artificial esparcida sobre la superficie de la hoja, estas dos criadas bajo condiciones controladas, la último supero en un centímetro el tamaño de las otras dos.
CONCLUSIONES
La utilización de una dieta artificial para larvas de B. polydamas bajo condiciones de luminosidad, temperatura y humedad controladas, fue recibida por las larvas únicamente esparcida sobre hojas de A. maxima como suplemento alimenticio, mostrando efectos en la sobrevivencia, el crecimiento y la longevidad de larvas de B. polydama,s hasta llevarlas al estado adulto, lo cual convierte este tratamiento en una opción para la cría de mariposas de esta especie con propósitos de investigación o en programas de manejo y conservación.
Por otro lado, los diferentes tratamientos utilizados probaron que la historia de vida de las mariposas de B. polydamas, está mediada por la especificidad de la relación Troidini-Aristolochia y que el comportamiento de selección de la planta, en este caso de A. máxima, es indispensable para su desarrollo. Igualmente descarta la opción de suministrar solamente dieta artificial, puesto que afecta los rasgos de historia de vida de los individuos causando hasta la muerte desde los primeros instar.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos a la Asociación Parque Natural Mana Dulce, al profesor Juan Carulla y al personal del Laboratorio de Nutrición de la UN, a los estudiantes del Laboratorio de Aromas y Productos Marinos de la UN y al profesor Jairo Cerón del Laboratorio de Entomología del IBUN. La investigación y experimentación, fuente de este articulo, se logro gracias al apoyo financiero de la Vicerrectoría de. Investigaciones de la Universidad Nacional de Colombia.
BIBLIOGRAFIA
A.O.A.C., Oficial Methods of Analysis of the Association of Analytical Chemists. (14th ed); 1984.
[ Links ]AVILA TO, WOODS HA, RAGUSTO RA. Effects of dietary variation on growth, composition, and maturation of Manduca sexta (Sphingidae: Lepidoptera). J. Insect. Physiol. 2003;49: 293-306.
[ Links ]BARROS EG. Within and between species scaling in the weight, water, carbon and nitrogen contents of eggs and neonate larvae of twelve satyrine butterflies (Lepidoptera: Nymphalidae). Eur. J. Entomol. 2006;103: 559-568.
[ Links ]BERENBAUM MR. Chemistry and oligophagy in the papilionidae. En: Scriber J., M.; Tsubaki, Y.; Lederhouse R., C. (eds). Swallowtail Butterflies: Their ecology and evolutionary biology. Scientific Publishers. 1995: 457 pp.
[ Links ]BOGGS LC. Nutritional and life-history determinants of resource allocation in holometabolous insects. Am. Nat. 1981;17: 692-709.
[ Links ]BOGGS LC. Resource allocation: Exploring connection between foraging and life history. Funct Ecol. 1992;6: 508 - 518.
[ Links ]BOGGS LC. The effect of adult food limitation on life history traits in Speyeria mormonia (Lepidoptera: Nymphalidae). Ecology. 1993;74(2):433-441.
[ Links ]BOGGS LC. Reproductive allocation from reserves and income in butterfly species with differing adult diets. Ecology. 1997a;78(1): 181-191.
[ Links ]BOGGS LC. Dynamics of reproductive allocation from juvenile and adult feeding: radiotracer studies. Ecology. 1997b;78(11): 192-202.
[ Links ]BROWN KS, DAMMAN AJ, FEENY P. Troidine Swallowatils (Lepidoptera: Papilionidae) in Southeastern Brazil: Natural History and Foodplant Relationships. J. Res. Lepid. 1980;19(4): 199-226.
[ Links ]CHAI P. Field observatios and feeding experiments on the responses of rofus-tailed jacamars (Galbula ruficauda) to free-flying butterflies in a tropical rainforest.Biol. J. Linn. Soc. 1986;29: 161-189.
[ Links ]CLARO RC, CORREA H, DUQUE C, RUIZ N. Ácido aristolóquico I y II en Aristolochia maxima (aristolochiaceae) y larvas de Battus polydamas polydamas y Parides panares erythrus (papilioninae).Acta biol Colomb. 2007;12 (2):59-68.
[ Links ]COHEN AC. Formalizing Insect Rearing and Artifitial Diet Technology. Am Entomol. 2001;47 (4): 198-206.
[ Links ]CONSTANTINO LM. Mariposas diurnas (Lepidoptera: rophalocera) del choco biogeográfico, Colombia: Diversidad, Sistemas de Cría, Manejo Sostenible de Poblaciones Silvestres y Estrategias para la Conservación. Informe final. Estudio de Factibilidad Técnica. Proyecto Biopácifico. Ministerio de Medio Ambiente. 1997: 125 p
[ Links ]DA SILVA LK. Interações evolutivas entre borboletas da tribo troidini (Papilionidae, Papilioninae) e suas plantas hospedeiras no genero Aristolochia (Aristolochiaceae). Tese apresentada ao Instituto de Biologia da Universidade Estadual de Campinas para obtenção do título de Doutor em Ecologia. 2005: 181 p
[ Links ]ELLIS A. Effects of hostplant species and artificial diet on growth of buckeye (Junonia coenia) and painted lady (Vanessa cardui) caterpillars (Nymphalidae). J Lepid Soc. 1998;52(1): 73-83.
[ Links ]FAGUA GG, RUIZ N. Relaciones de herbivoría entre papiliónidas (Lepidoptera) y especies de Aristolochia (Aristolochiaceae). En: Andrade C., G.; García A., G.; Fernández, F. (eds). Insectos de Colombia. Centro Editorial Javeriano (ceja), Academia Colombiana de Ciencias Exactas, Físicas y Naturales. 1996: 541.
[ Links ]FAGUA GG, RUIZ N, GONZALES F, ANDRADE MG. Calidad del hospedero en el ciclo de desarrollo de Battus polydamas (Lepidoptera: Papilionidae: Troidini). Revista Colombiana de Entomología. 1998; 24 (3 - 4): 131 - 140.
[ Links ]FISCHER K, O´BRIEN DM, BOGGS CL. Allocation of larval and adult resources to reproduction in a fruit-feeding butterfly. Funct Ecol. 2004;18: 656-663.
[ Links ]FORDYCE JA. The lethal plant defense paradox remains: inducible host-plant aristolochic acids and the growth and defense of the pipevine swallowtail. Entomol. Exp Appl. 2001;100: 339-346.
[ Links ]GARRAWAY E, BAILEY AJA, EMMEL C. Contribution to the ecology and conservation biology of the endangered Papilio homerus (Lepidoptera: Papipilionidae). Trop Lepid. 1993;4(2): 83-91
[ Links ]GENC H, NATION JL. An artifitial diet for the butterfly Phyciodes phaon (Lepidoptera: Nymphalidae). Fla Entomolo. 2004;87 (2): 194-198.
[ Links ]HALL DW, BUTTLER JF. Polydamas Swallowtail, Battus polydamas lucayus (Rothschild & Jordan). EENY-062. Institute of Food and Agricultural Sciences, Universidad de la Florida. 1998.
[ Links ]HAUFLER JB, SERVELLO FA. Techniques for wildlife nutritional analyses. En: T. A. Bookhout. Research and management techniques for wildlife and habitats. The Wildlife Society, Bethesda, Maryland, USA. 1994: 307- 323.
[ Links ]HOCHULI DF. Insect hervibory and ontogeny: how do growth and development influence feeding behaviour, morphology and host use?. Austral Ecol. 2001;26:563-570.
[ Links ]IDEAM. Instituto de Hidrológica, Meteorología y Estudios Ambientales. Sistema de Información Nacional Ambiental. 2006.
[ Links ]JAENIKE J. Host specialization on phytophagous insects. Annu Rev Ecol Syst. 1990. 21:243-73.
[ Links ]KLITZKE FC, BROWN KS. The occurrence of aristolochic acids in neotropical troidine swallowtails (Lepidoptera: Papilionidae). Chemoecology. 2000;10: 19-102.
[ Links ]LE CROM JF, CONTANTINO LM, SALAZAR JA. Mariposas de Colombia, Tomo I, Papilionidae. Carlec Ltda. 2002:112 pp.
[ Links ]LEE AD. Tasty generalists and nasty specialists? Antipredator mechanisms in tropical Lepidopteran larvae. Ecology. 1995;76(5): 1483-1496.
[ Links ]MARSHALL DL. Male nutrient investment in the lepdioptera: what nutrients should males invest?. Am Nat. 1982;120:273-279.
[ Links ]MATTONI R, LONGCORE T, KRENOVA Z, LIPMAN A. Mass rearing the endangered Palos Verdes blue butterfly (Glaucopsyche lygdamus palosverdesensis: Lycaenidae). J Res Lepid. 2003;37: 55-67.
[ Links ]MIX DB, GUINAUDEAU H, SHAMMA M. The aristolochic acids and aristolactams. J Nat Prod. 1983;45(6): 657-666.
[ Links ]MORTON AC. Rearing Butterflies on Artificial Diets. J Res Lepid. 1979;18 (4): 221-227.
[ Links ]NAGY JG, HAUFLER JB. Nutrición de los animales silvestres.. En: Rodríguez T., R. (ed.). Manual de técnicas de gestión de vida silvestre. The Wild Life Society. 1987: 135-149
[ Links ]NEW TR, PYLE R M, THOMAS JA, THOMAS CD, HAMMOND PC. Butterfly Conservation Management. Annu Rev Entomol. 1995;40: 57- 83.
[ Links ]O´BRIEN DM, BOGGS CL, FOGEL ML. Making eggs from néctar: the role of life history and dietary carbón turnover in butterfly reproductive resource allocation. Oikos. 2004;105 (2): 279 - 291.
[ Links ]PANIZZI AR, PARRA JRP. Ecologia nutricional de insetos e suas implcaçôes no manejo de pragas. Editora Manole LTDA. Sâo Paulo. Brasil. 1991:359
[ Links ]PARSONS JM. Butterfly farming and trading in the Indo- Australia region and its role in tropical conservation. Topic Lepid. 1992;3(1): 1-31.
[ Links ]PARSONS JM. Butterfly farming and trading in the Indo- Australia region and its benefits in the conservation of swallowtails and their tropical forest habitats.. En: Scriber J., M.; Tsubaki, Y.; Lederhouse R., C. (eds). Swallowtail Butterflies: Their Ecology and Evolution Biology, Scientific Publishers. 1995:371-391
[ Links ]RAUBENHEIMER D, SIMPSON S. Nutrient transfer functions: the site of integration between feeding behaviour and nutritional physiology. Chemoecology 1998; 8: 61-68.
[ Links ]SCRIBER JM, FEENY P. Growth of herbivorous caterpillars in relation to feeding specialization and to the growth form of their food plants. Ecology. 1979;60(4): 829-850.
[ Links ]SCRIBER JM, SLANSKY F. The nutritional ecology of immature insects. Ann Rev Entomol. 1981;26: 183-211.
[ Links ]SILVA-BRANDÂO LK, SOLFERINI VN, TRIGO JB. Chemical and phylogenetic relationships among Aristolochic L. (Aristolochiaceae) from southeastern Brazil. Biochem. Syst. Ecol. 2006;34: 291-302.
[ Links ]SIME RK, FEENY PP, HARIBAL MM. Sequestration of aristolochic acids by the pipevine swallowtail, Battus philenor (L.): evidence and ecological implications. Chemoecology. 2000;10: 169-178.
[ Links ]SINGH P. Artificial diets for insects, mites, and spiders. IF1/Plenum New York, Washington, London. 1977: 594 pp.
[ Links ]SIYUAN C, ZENG L, GUANGWEN I. Research in Artificial Diet for Multi-Generation Rearing of Papilio xuthus. Journal Of South China Agricltural University. 1999;1: 1-14.
[ Links ]TELANG A, BOOTON V, CHAPMAN RF, WHEELER DE. How female caterpillars accumulate their nutrient reserves. J Insect Physiol. 2001;47: 1055-1064.
[ Links ]TYLER HA, BROWN KS, WILSON KH. Swallowtail Butterflies of the Americas: A Study In Biological Dynamics, Ecological Diversity, Byiosystematics And Conservation. Scientific Publishers. Gainesville. United States of America. 1994: 375 p.
[ Links ]VAN SOEST PJ. Nutritional ecology of the ruminant. Cornell University Press, Ithaca, NY, USA. 1982.
[ Links ]
