










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.15 n.1 Bogotá jan./abr. 2010
EFECTO DE HONGOS MICORRIZICOS ARBUSCULARES EN PLÁNTULAS DE Elaeis guineensis (Palmaceae) CON ALTO NIVEL DE P EN EL SUELO
EFFECT OF ARBUSCULAR MYCORRHIZAL FUNGI IN Elaeis guineensis (Palmaceae) SEEDLINGS WITH HIGH PHOSPHORUS LEVEL IN THE SOIL
SILVIA EUGENIA BARRERA BERDUGO1, Bióloga; NELSON F. RODRIGUEZ LOPEZ2 M.Sc. Fisiología Vegetal. 1Escuela de Biología, Facultad de Ciencias, Universidad Industrial de Santander, Calle 9, Carrera 27, ciudad universitaria, Bucaramanga, Colombia. silviaebarrera@tux.uis.edu.co . 2Profesor Escuela de Biología, Universidad Industrial de Santander. Bucaramanga Colombia. Grupo Nacional de Investigación en Ecofisiología & Metabolismo Vegetal Tropical. Actualmente, Estudiante-Doctorado en Fisiología Vegetal, UFV, Viçosa, MG, Brasil. fisionel@uis.edu.co
Presentado 25 de enero de 2009, aceptado 30 de junio de 2009, correcciones 13 de julio de 2009
RESUMEN
Los hongos micorrízicos arbusculares (HMA) facilitan la absorción de nutrientes a las plantas hospederas, por esta razón estos microorganismos cumplen un rol fundamental en el funcionamiento de los agroecosistemas. El objetivo de este trabajo fue evaluar el efecto de la asociación simbiótica entre HMA nativos y comerciales y plántulas de Elaeis guineensis en condiciones de vivero con un nivel alto de fósforo (P) en el suelo.
Plantas de tres meses de edad fueron sometidas a cuatro tratamientos: Inóculo Nativo (IN), Inóculo Comercial (IC), Mixto (M) y Testigo absoluto (TA). Se evaluaron los parámetros: peso seco total de la planta, peso seco raíz, peso seco parte aérea de la planta, altura de la planta, tasa de crecimiento relativo, colonización micorrízica y número de esporas en el suelo. Se realizaron tres muestreos, uno inicial (día 0), uno a los 45 y a los 90 días después del trasplante de las plántulas (ddt). Los datos fueron analizados mediante un ANOVA o Kruskall-Wallis según el comportamiento de los datos, seguido de un test de Duncan para comparar las medias o un test modificado de Tuckey para datos no paramétricos. Se observaron diferencias significativas en el número de esporas entre los tratamientos IN, M y TA, a los 45 ddt. Para la variable colonización micorrízica se observaron diferencias significativas a los 45 ddt entre los tratamientos IC y TA, mientras que a los 90 ddt se presentaron diferencias entre los tratamientos IN, IC y M, con respecto al TA. A pesar de que el nivel de P en el suelo fue alto, el porcentaje de colonización micorrízica estuvo por encima del 50% en los muestreos realizados a los 45 y 90 días. El tratamiento IN funcionó mejor que los tratamientos IC y M, para las condiciones edáficas de este experimento.
Palabras clave: E. guineensis, hongos micorrízicos arbusculares, número de esporas, colonización micorrízica, fósforo en suelo.
ABSTRACT
The arbuscular mycorrhizal fungi (AMF) facilitate the absorption of nutrients to the host plants. These microorganisms therefore, fulfill a fundamental roll in the operation of agroecosytems. The aim of this work was to evaluate the effect between native and commercial arbuscular mycorrhizal fungi and Elaeis guineensis seedlings in greenhouse conditions with high P level. Plants of three months of the age were put under four treatments: Native inoculate (NI), Foreign inoculate (FI), Mixture (M) and control (TA). It was evaluated total dry weight of the plant, root and aerial part dry weight, height of the plant, relative growth rate (RGR), mycorrhizal colonization and spores number. At 45 and 90 days after the transplant these variable were evaluated. An initial sampling dry mass was taking by to determine RGR. The data were analyzed through the ANOVA or Kruskall-Wallis according to the behavior of the data, followed of a test of averages of Duncan or a modified test of Tuckey for nonparametric data. Significant differences in the spores number between the treatments NI, FI, M and TA and between NI and M to the 45 days appeared. In the colonization mycorrhizal differences between the treatments FA and TA were observed the 45 days. To the 90 days between all the treatments with respect to the TA. Although the P level in the soil was high, also it was it the mycorrhizal colonization at the two evaluated times. Through time the treatment NI worked better in the soil edaphic conditions used in this experiment.
Key words: E. guineensis, arbuscular mycorrhizal fungi, spores number, mycorrhizal colonization, phosphorus in the soil.
INTRODUCCIÓN
Las micorrizas arbusculares son asociaciones simbióticas formadas entre hongos micorrízicos arbusculares (HMA) y la mayoría de las plantas vasculares, para actuar como un complemento de la raíz de la planta en la toma de nutrientes (Colozzi Filho y Cardoso, 2000), especialmente en la absorción de P (Smith y Read, 1997; Requena et ál., 2007).
Este complemento se da gracias a la extensión de un micelio extraradical que llega más allá de la zona de depleción de la raíz (Blancof y Salas, 1997), con el fin de absorber agua, además de nutrientes del suelo y formar puentes hifales con las raíces de las plantas cercanas, como mecanismo de transferencia de nutrientes entre hospederos (Peterson et ál., 2004).
La importancia ecológica de los HMA radica en los beneficios que aportan a las plantas que colonizan (Rodriguez et ál., 2004) y al grado de adaptación que los HMA presentan a determinadas condiciones edafoclimáticas, lo cual le permite mayores beneficios a un determinado hospedero con respecto a otros (Cuenca et ál., 2004). En estudios ecológicos se ha evaluado el efecto de HMA nativos y comerciales sobre las especies vegetales, observándose respuestas tanto positivas como negativas en el crecimiento de las plantas, por lo que también dependen del genotipo de la planta con la cual se asocian (Klironomos, 2003).
Elaeis guineensis Jacq. (Palma de aceite) es una planta tropical perenne, con un ciclo de vida que puede sobrepasar los 100 años y un promedio de vida productiva de 25 a 30 años (Revelo, 2002). Su importancia radica en la producción de aceite, por lo que se requiere obtener un alto rendimiento de los cultivos debido a la gran demanda que genera en el sector comercial el procesamiento del fruto para la fabricación de productos como aceite para freír, margarinas, aceite para motores diesel, alcoholes ácidos, etc. (CEA y J.E.Austin Associates, 2002).
La palma de aceite forma asociación simbiótica con HMA nativos en su hábitat natural (Sieverding, 1991). Chu, (1997) y Corley y Tinker, (2003) han planteado que es funcionalmente dependiente de estos hongos ya que plántulas crecidas en suelo estéril no se desarrollan adecuadamente al no estar colonizadas por este tipo de microorganismos. En condiciones de vivero, han sido reportados resultados favorables de los HMA sobre el crecimiento de la palma de aceite, en intervalos de tiempo cortos y largos, así como en la absorción de nutrientes (Blal et ál., 1990; Chu, 1997), aún con concentraciones altas de fósforo en el suelo (Motta y Munévar, 2005). Teniendo como base esos datos, el objetivo de este trabajo fue evaluar el efecto de la asociación simbiótica entre HMA nativos y comerciales y plántulas de E. guineensis en condiciones de vivero con un nivel alto de fósforo (P) en el suelo.
MATERIALES Y METODOS
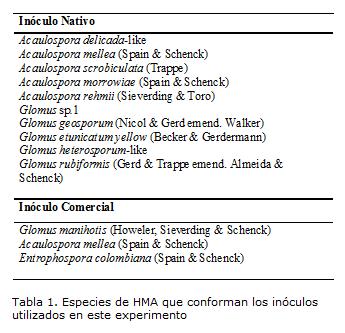
El experimento se realizó en la Granja Experimental Guatiguará (Piedecuesta-Santander), en condiciones de vivero. Se utilizaron plantas de 3 meses de edad (provenían de fase de vivero). La temperatura mínima promedio fue de 19ºC y la temperatura máxima 35ºC. Las plántulas de E. guineensis cultivar Tenera fueron trasplantadas a bolsas de polietileno de 12 kg que contenían una mezcla de suelo y arena de río en una proporción 3:1. El suelo mostró una textura franco-arenosa con las siguientes características químicas: pH=6.8, MO=1.9%, N=0.095%, P=59 ppm y Ca=11.7, K=0.32 y Mg=1.2 meq/100 g de suelo y fue esterilizado previamente con Basamid (Dazomet: 3,5-dimetil-(2H)-tetrahidro-1,3,5 tiadazina-2-tiona) a la dosis recomendada (60 g/m2). Los inóculos utilizados fueron: inóculo nativo (IN), aislado de las zonas palmicultoras de Santander con 32 esporas/g de suelo y el inóculo Comercial (IC), producto SUPPRAcon 34 esporas/g de suelo; los cuales se aplicaron a razón de 25g por planta y 12.5g de cada inóculo para el inóculo mixto (M), mezcla de IN con IC; así como un testigo absoluto (TA)La identificación de las especies del inóculo nativo fue realizada en el laboratorio de microbiología de suelos de la Universidad de Blumenau SC Brasil; tanto estas como las especies presentes en el inóculo comercial se muestran en la Tabla 1. Cada 10 días se aplicaron 100 ml de solución de nutrientes de Hoagland baja en P (0,11 g/L NH4H2PO4).
PARAMETROS DE CRECIMIENTO VEGETAL
Los parámetros de crecimiento evaluados en la planta fueron el peso seco total de la planta (PST), peso seco raíz (PSR), peso seco parte aérea de la planta (PSPA), altura de la planta (AP), y se calculó la tasa de crecimiento relativa (TCR). El área foliar se estimó usando el programa CompuEye, Leaf & Symptom Area Versión. 11.001 (Bakr, 2004).
FASE DE LABORATORIO
Para determinar el porcentaje de colonización micorrízica se tomaron 10 raicillas al azar por planta de un centímetro de largo y se tiñeron con azul de tripán siguiendo la metodología de Phillips y Hayman, (1970) con algunas modificaciones hechas por Sieverding (1991), apropiadas para suelos tropicales. El porcentaje de colonización se obtuvo a partir del número de campos positivos (presencia de arbúsculos, vesículas o hifas) sobre el número de campos totales multiplicado X 100. Esto se hizo con ayuda de un microscopio óptico (LW Scientific). Para el aislamiento de esporas se tomaron 10 g de suelo de la rizosfera de la planta al momento del muestreo, siguiendo el método descrito por Gerdemann y Nicolson, (1963) de tamizado húmedo y centrifugación con sacarosa teniendo en cuenta las modificaciones hechas realizadas por Sieverding, (1991) apropiadas para suelos tropicales. La cuantificación se realizó en un estereoscopio (Olimpus) utilizando placa de conteo.
DISEÑO EXPERIMENTAL Y ANÁLISIS ESTADISTICO
Se utilizó un diseño completamente aleatorizado, con los tratamientos: IN; IC; Mixto y TA. En cada uno de los tratamientos se emplearon 15 unidades experimentales y se tomaron cinco plantas por muestreo. Se realizaron tres muestreos, uno inicial de masa seca al comienzo del experimento, el segundo a los 45 días después del trasplante de las plantas (ddt) y el tercero a los 90 ddt.
Los datos obtenidos fueron analizados por muestreos de forma independiente y se realizaron pruebas de normalidad, homogeneidad de varianza y posteriormente un ANOVA o Kruskal Wallis según se comportaran los datos como paramétricos o no paramétricos. Se hizo una prueba de comparación de medias de Duncan (p≤0.05) o una prueba de Tuckey modificada para los datos no paramétricos donde se presentaron diferencias (Zar, 1999). El análisis se realizó en el programa SPSS versión 12.0 para Windows (SPSS Inc., 2003). Para calcular las medias de la TCR en cada intervalo de tiempo se utilizó el método de la aproximación clásica mediante la hoja de Excel propuesta por Hunt., et ál., (2002) para cada tratamiento, seguida de una prueba de comparación de medias de Duncan (p≤0.05) para determinar las diferencias significativas entre estos.
RESULTADOS
RESPUESTAS DE CRECIMIENTO DE LAS PLANTAS
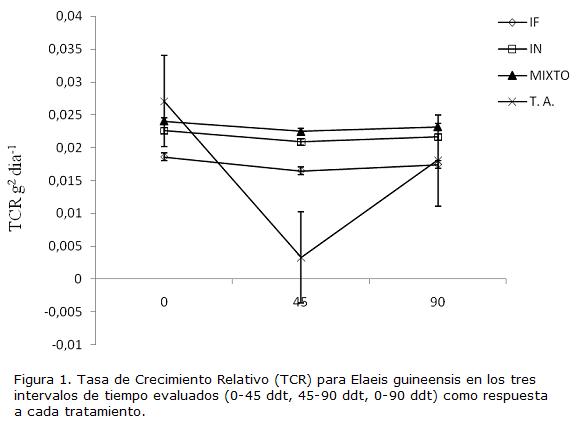
No se presentaron diferencias significativas a los 45 ddt y a los 90 ddt para las respuestas de crecimiento de las plantas (Tabla 2). En los tres intervalos de tiempo evaluados no se presentaron diferencias significativas para la TCR, sin embargo, se puede apreciar que hay una disminución de esta variable para el segundo intervalo de tiempo, mientras que al final del experimento se empieza a notar un leve aumento (Fig 1). Para el tratamiento TA la disminución en el segundo intervalo de tiempo fue mayor que en el resto de los tratamientos (Fig 1).
COLONIZACIÓN MICORRÍZICA Y NÚMERO DE ESPORAS
A los 45 ddt el porcentaje de colonización micorrízica fue alto a pesar de que el nivel de P en el suelo también fue alto (Tabla 2). El tratamiento IC presentó la colonización micorrízica más alta con 69.90% (± 8.08). Se observaron diferencias significativas para esta variable entre los tratamientos IC y TA.
Se observaron diferencias en el porcentaje de colonización micorrízica a los 90 días entre los tratamientos IN, IC y M con respecto al TA. En el tratamiento TA se incrementó la colonización micorrízica de 19.94 % a 38.04 % (Tabla 2) a través del tiempo.
Se observaron diferencias significativas en el número de esporas entre los tratamientos M-IN y M-TA a los 45 ddt (Tabla 2). Fueron aisladas 81 esporas/gramo (± 11.4) de suelo del tratamiento M, siendo este el de mayor número de esporas aislado. A los 90 ddt no se presentaron diferencias significativas en el número de esporas.
DISCUSIÓN
Debido a que el tratamiento TA presentó colonización micorrízica, no se observaron diferencias entre los tratamientos para las variables de crecimiento evaluadas en las plantas, no obstante, se tiene evidencia de que plántulas de palma de aceite en suelo estéril no se desarrollaron sin la presencia de HMA, aún cuando el nivel de P total en el suelo es de 66 ppm (Blal et ál., 1990).
Efectos significativos en el peso seco de plantas micorrizadas de E. guineensis con respecto a plantas no micorrizadas, han sido encontrados después de tres meses de estudio (Blal et ál., 1990), así como diferencias significativas en variables de crecimiento de la planta se han presentado en etapa de previvero (Chu, 1997). A pesar de que en nuestro experimento la contaminación de las plantas del TA no permitió observar esas diferencias, tampoco se pudieron apreciar efectos entre los otros tratamientos. Esto indica que es necesario escoger las especies fúngicas adecuadas para determinada planta y para características edáficas específicas.
Es importante tener en cuenta que la formación de la simbiosis micorrízica genera un alto costo para la planta hospedera (Schroeder y Janos, 2004), ya que se estima que del carbono fijado en la fotosíntesis, del 4% al 20% es movilizado hacia la raíz para la formación de la simbiosis (Bago et ál., 2000; Sánchez de Prager, 2007). De esta forma, el costo de los carbohidratos que van a ser transferidos a la raíz es más alto en las plantas micorrizadas que en las no micorrizadas (Bryla y Eissenstat, 2005).
La variedad de la planta es otro aspecto o factor a tener en cuenta en estudios posteriores, ya que el tiempo en vivero no es el mismo para todas las variedades y el efecto que los HMA tienen sobre diferentes variedades de una misma especie no es el mismo, razón por la cual puede estar regulada esta interacción por el genoma tanto de la planta como del hospedero.
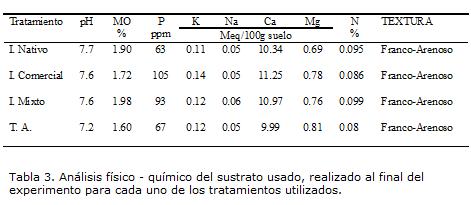
Aunque el nivel de P en el suelo fue alto al comienzo y al final del experimento (Tabla 3), el porcentaje de colonización micorrízica estuvo por encima del 50% a lo largo del experimento, indicando esto que la palma de aceite es una planta que forma micorrización en esta condición. Esto podría deberse a que la palma de aceite es dependiente de la micorrización para su desarrollo, en este caso sería micotrófica obligada y eso explicaría el porqué la planta responde a la colonización, aun cuando el nivel de P en el suelo es alto (Saggin-Junior y Lovato, 1999). En este experimento las plantas del tratamiento TA presentaron colonización micorrizica, aunque fue el porcentaje más bajo entre los tratamientos durante el experimento. Se ha mencionado que cuando el nivel de P en el suelo es alto y hay colonización de HMA en la raíz, se puede dar una reducción en el crecimiento de la planta (Kahiluoto et ál., 2001), esto nos muestra que la relación entre planta y HMA puede pasar de mutualismo a parasitismo y viceversa (Klironomos, 2003; Jones y Smith, 2004).
BIBLIOGRAFÍA
BAGO B, PFEFFER P, SHACHAR-HILL Y. Carbon Metabolism and Transport in Arbuscular Mycorrhizas. J Plant Physiol. 2000;124: 949-95.
[ Links ]CompuEye, Leaf & Symptom Area. [Programa de ordenador]. Version 11.001. Cairo (Egipto); 2005.
[ Links ]BLAL B, MOREL C, GIANINAZZI-PEARSON V, FARDEAU JC, GIANINAZZI S. Influence of Vesicular-arbuscular Mycorrhizae on Phosphate Fertilizar Efficiency in two Tropical Acid Soils Planted with Micropropagated Oil Palm (Elaeis guineensis jacq.). Biol Fertil Soils. 1990;9: 43-48.
[ Links ]BLANCOF F, SALAS E. Micorrizas en la Agricultura: Contexto Mundial e Investigación Realizada en Costa Rica. Agronomía Costarricense. 1997;21(1): 55-67.
[ Links ]BRYLA D, EISSENSTAT D. Respiratory Costs of Mycorrhizal Associations. En: Lambers H, Ribas-Carbo M, editores. Plant Respiration from Cell to Ecosystem. Urbana, Illinois: University Illinois; 2005. p. 207-225.
[ Links ]COLOZZI FILHO A, CARDOSO E. Detecção de Fungos Micorrízicos Arbusculares em Raízes de Cafeeiro e de Crotolária Cultivada na Entrelinha. Pesq Agrop Bras. 2000; 35(10): 2033-2042.
[ Links ]CORLEY RHV, TINKER PB. The Oil Palm. 4 ed. Iowa: Blackwell Science; 2003.
[ Links ]CORPORACIÓN CEA & J.E.AUSTIN ASSOCIATES. Estudio del Mercado Mundial para Aceite de Palma Africana. Bogotá; 2002.
[ Links ]CHU EY. Influência dos Fungos Micorrízicos Arbusculares e Níveis de Adubação do no Crescimento Inicial de Mudas de Dendê, Boletim de Pesquisa. Embrapa. 1997;176: 1-20.
[ Links ]CUENCA G, DE ANDRADE Z, LOVERA M, FAJARDO L, MENESES E. The Effect of two Arbuscular Mycorrhizal Inocula of Contrasting Richness and the same Mycorrhizal Potential on the Growth an Survival of Wild Plant Species from La Gran Sabana, Venezuela. Can J Bot. 2004;82: 582-589.
[ Links ]GERDEMANN JW, NICOLSON TH. Spores of Mycorrhizal Engogone Species Extracted by Wet Sieving and Decanting. Trans Brit Mycol Soc. Cambridge. 1963;46: 235-244.
[ Links ]JONES M, SMITH S. Exploring Functional Definitions Mycorrhizas: Are Mycorrhizas always Mutualism? Can J Bot. 2004;82: 1089-1109.
[ Links ]HUNT R, CAUSTON DR, SHIPLEY B, ASKEW A. A Modern Tool for Classical Plant Growth Analysis. Ann Bot. 2002;90: 485-488.
[ Links ]KAHILUOTO H, KETOJA E, VESTBERG M, SAARELA I. Promotion of AM Utilization through Reduced P Fertilization. 2. Field studies. Plant Soil. 2001;231: 65-79.
[ Links ]KLIRONOMOS K. Variation in Plant Response to Native and Exotic Arbuscular Mycorrhizal Fungi. Ecology. 2003;84(9): 2292-2301.
[ Links ]MOTTA D, MUNÉVAR F. Respuesta de Plántulas de Palma de Aceite a la Micorrización. Palmas. 2005;26(3): 11-20.
[ Links ]PETERSON RL, MASSICOTE H, MELVILLE L. Mycorrhizas: Anatomy and Cell Biology. Ottawa: NRC Research press; 2004.
[ Links ]PHILLIPS JM, HAYMAN DS. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-arbuscular Micorrhizal Fungi for Rapid Assessment of Infection. Trans Brit Mycol Soc, Cambridge. 1970;55: 158-161.
[ Links ]REQUENA N, SERRANO E, OCÓN A, BREUNINGER M. Plant Signals and Fungal Perceptions during Arbuscular Mycorrhizal Establishment. Phytochemistry. 2007; 68:33-4.
[ Links ]REVELO M. Generalidades sobre Botánica Morfología y Fisiología. En: Revelo, M. editores. Palmicultura Moderna Volumen I. Bogotá: Sociedad las Palmas Ltda; 2002. p. 9-18.
[ Links ]RODRÍGUEZ Y, DE LA NOVAL B, FERNÁNDEZ F, RODRÍGUEZ P. Estudio Comparativo de Seis Cepas de Hongos Micorrízicos Arbusculares en su Interacción con el Tomate (Lycopersicum esculentum m. var "amalia"). Ecología Aplicada. 2004; l3(1y2): 162-171.
[ Links ]SAGGIN-JUNIOR OJ, LOVATO PE. Aplicação de Micorrizas Arbusculares na Produção de Mudas e Plantas Micropropagadas. En: Siqueira JO, Moreira FMS, Lopes AS, Guilherme LRG, Faquin V, Furtini Neta AE, Carvalho Jg, editores. Inter-Relação Fertilidade, Biologia do Solo e Nutrição de Plantas. Lavras: Universidade Federal de Lavras. Departamento de Ciência do Solo. Sociedade Brasileira de Ciência do Solo; 1999. p. 687-703.
[ Links ]SANCHEZ M. Las Micorrizas: Estrategia Compartida para Colonizar el Suelo. En Sanchez, M. editores. Las endomicorrizas: Expresión bioedáfica de Importancia en el Trópico. Palmira. Universidad Nacional de Colombia; 2007. p. 115-175.
[ Links ]SMITH SE, READ DJ. Mycorrhizal Symbiosis. 2 ed. Londres: Academic Press Limited; 1997.
[ Links ]SIEVERDING E. Vesicular-arbuscular Mycorrhiza Management in Tropical Agrosystems. Eschborn: Deutsche Gesellschaft für Technische Zusammenarbeit GTZ; 1991.
[ Links ]SCHROEDER MS, JANOS D. Phosphorus and Intraspecific Density Alter Plant Responses to Arbuscular Mycorrhizas. Plant Soil. 2004;264: 335-348.
[ Links ]ZAR JH. Biostatistical Analysis. 4 ed. New Jersey: Prentice Hall; 1999.
[ Links ]
