










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.15 no.1 Bogotá Jan./Apr. 2010
Estructura espacial de Eucinostomus argenteus (Pisces: Gerreidae) en la zona norte del Caribe colombiano
Spatial structure of Eucinostomus argenteus (Pisces: Gerreidae) in the north zone of the Colombian Caribbean Sea.
Maria Pacheco1, Jorge Paramo1,2 Claudia Sánchez1 3 1Grupo de Investigación Ciencia y Tecnología Pesquera Tropical (CITEPT), Universidad del Magdalena, Cra. 32 No. 22-08 Avenida del Ferrocarril, Santa Marta, Colombia. ingmarypp@yahoo.es 2 Universität Bremen, Zentrum für Marine Tropenökologie (ZMT), Fahrenheit Strasse 6, Bremen, Alemania. jorge.paramo@zmt-bremen.de 3 Instituto de Investigaciones Marinas y Costeras (INVEMAR), Cerro Punta Betín, Apartado 1016, Santa Marta, Colombia, claudia_sanchez@invemar.org.co
Presentado 25 de junio de 2009, aceptado 19 de octubre de 2009, correcciones 4 de noviembre de 2009
RESUMEN
Esta investigación se llevó a cabo para determinar la distribución espacial de la densidad (captura por unidad de área) - CPUA y de la estructura de tallas de Eucinostomus argenteus y su relación con las variables ambientales en la zona norte del Caribe colombiano. Los datos fueron se tomaron durante un crucero de investigación realizado en la época seca (Febrero de 2006) entre Puerto Estrella (La Guajira) y Santa Marta (Magdalena). Las muestras biológicas se colectaron con una red de arrastre (en estratos de profundidad < 50 m y 50-100 m) siguiendo un diseño de muestreo sistemático. Los individuos maduros y de tallas mayores se encontraron distribuidos principalmente entre Manaure y Punta Gallinas, donde la plataforma continental es muy ancha y la oceanografía local esta modulada por la surgencia estacional. Los peces juveniles, se distribuyeron al sur del área de estudio, entre Boca Camarones y el Río Buritaca, cerca de la costa. En éste sector la plataforma es muy estrecha y con alta productividad biológica por la presencia de ríos, manglares y pastos marinos, sirviendo como zona de alimentación y refugio para E. argenteus.En general, los resultados sugieren que las variables ambientales son importantes para la distribución espacial de la abundancia y las tallas de E.argenteus en la zona norte del Caribe colombiano, siendo la temperatura y la profundidad las variables que predijeron mejor la distribución espacial de la especie.
Palabras clave: Distribución espacial, Eucinostomus argenteus, geostadística, GAM, Caribe colombiano.
ABSTRACT
This research was carried out to determine the spatial distribution of density (catch per unit area) - CPUA and size structure of Eucinostomus argenteus and their relationship with environmental variables in the north zone of the Colombian Caribbean sea. The data comes from a survey of fisheries research during the dry season (February of 2006) between Puerto Estrella (La Guajira) and Santa Marta (Magdalena). Biological samples were taken with a bottom trawl net (< 50 m and 50-100 m stratum of depth) following a systematic design survey. The mature fishes with higher sizes were mainly located in the sector between Manaure and Punta Gallinas, where the continental shelf is very wide and the local oceanography is modulated by the seasonal upwelling. The juvenile fishes were found toward the south of the study area between Boca Camarones and the Rio Buritaca, close to the coast. In this sector the continental shelf is very narrow with high biological productivity for the presence of rivers, mangroves and sea grasses, which serve as feeding grounds and nursery habitat for E. argenteus. In general, our results suggest that environmental conditions were important determinants of spatial distribution of abundance and sizes of E. argenteus in the north zone of the Colombian Caribbean Sea, with temperature and depth as variables that better predict the spatial distribution of this specie.
Key words: spatial distribution, Eucinostomus argenteus, geostatistic, GAM, Colombian Caribbean.
INTRODUCCIÓN
Eucinostomus argenteus hace parte importante en la estructura comunitaria de los ecosistemas costeros y marinos tropicales debido a su gran abundancia, (De Grado y Bashirullah, 2001, Godefroid et ál., 2001, Ordóñez-López y García-Hernández, 2005). Esta especie es considerada dominante en playas arenosas desprovistas de vegetación, y hace parte de la captura incidental de la pesca de arrastre camaronera (Vianna y Costa-Neves, 2004). En el Caribe colombiano, la mayoría de los estudios sobre la familia Gerreidae se han enfocado hacia los hábitos tróficos y biológicos (Arenas, 1990, Arenas y Acero, 1992, Navajas y García, 1999), pero muy pocos se han dirigido a conocer su distribución espacial y abundancia. Algunos autores han documentado la dinámica poblacional y distribución espacial de Eugerres plumieri y aspectos tróficos de la familia Gerreidae en la Ciénaga Grande de Santa Marta (CGSM) (Arenas y Acero, 1992, Rueda y Santos-Martínez, 1998, Rueda y Defeo, 2003), Ciénaga de la Virgen (Rodríguez, 1982) y Ciénaga de Tesca (Acosta, 1985). Se sabe que los peces tienden a estar organizados en estructuras en forma de agregaciones, de ahí que su distribución en el espacio o el tiempo no es al azar, sino que están bien organizados de acuerdo a factores físicos, químicos y biológicos que controlan su actividad, tal como la alimentación, evitar la depredación, migración, reproducción y selección del hábitat (Simard et ál., 1992). Además, durante su ciclo de vida, los peces experimentan desplazamientos o migraciones entre zonas geográficas, así, los individuos determinan de una manera u otra la regularidad y cronología de las actividades pesqueras. Por lo tanto, los administradores pesqueros requieren información sobre la abundancia, distribución espacial y estructura de tallas de un stock en particular, que conlleven a su manejo sostenido (Rivoirard et ál., 2000). La geoestadística da a la ecología una novedosa herramienta para la interpretación de los patrones espaciales de los organismos, de los numerosos componentes ambientales con los cuales estos interactúan, y la conexión de dependencia espacial entre organismos y su ambiente (Rossi et ál., 1992, Rivoirard et ál., 2000). Esta investigación se llevó a cabo para determinar la distribución espacial de la densidad (Captura Por Unidad de Área) - CPUA y de la estructura de tallas de Eucinostomus argenteus y su relación con las variables ambientales durante la época seca en la zona norte del Caribe colombiano. Con esta información se contribuye al conocimiento de la biología de la especie, considerando los procesos ecológicos y oceanográficos, bajo un enfoque ecosistémico para el manejo de las pesquerías (Bianchi y Skjoldal, 2009).
ÁREA DE ESTUDIO
El área de estudio comprendió la zona norte del Caribe colombiano a lo largo de la plataforma continental entre Puerto Estrella (12° 24' N - 71° 48' W) y Santa Marta (11° 16' N - 74° 14' W) (Figura.1). La plataforma continental frente a la península de La Guajira es muy estrecha, con la isobata de los 200 metros (m) a solo 10 millas náuticas (mn) de la costa en la parte mas ancha. Al oeste, la plataforma se ensancha desde el Cabo de la Vela hasta Riohacha hasta una distancia máxima de 25 mn y luego se reduce de nuevo hasta que casi desaparece frente al Parque Nacional Natural Tayrona (PNNT) (Fig. 1). La región desde La Guajira hasta Santa Marta se ve afectada por la corriente del Caribe y por la surgencia estacional de aguas subtropicales durante la época seca, de diciembre -abril y mediados de agosto (Pujos et ál., 1986, Bula-Meyer, 1990, Andrade et ál., 2003).
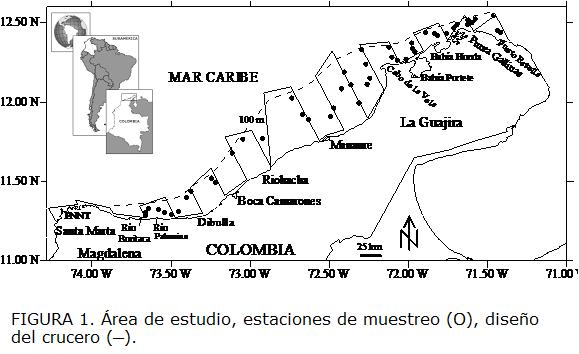
MATERIALES Y MÉTODOS
Se realizó un crucero de investigación pesquera entre el 6 y el 17 de febrero de 2006, usando un diseño de muestreo sistemático, con transectos paralelos entre sí y perpendiculares a la costa, espaciados regularmente cada 10 mn (Figura 1). Sin embargo, los sitios de muestreo se realizaron considerando solo las áreas arrastrables, lo cual involucró algo de aleatoriedad en la toma de muestras. Las muestras biológicas (1.655 individuos de E. argenteus) se obtuvieron mediante una red de arrastre demersal de 20,6 m de relinga superior, 25,6 m de relinga inferior, y 4,5 cm de tamaño de malla del copo. Se realizaron 48 arrastres con una duración promedio por arrastre de 15 minutos (Folmer y Pennington, 2000) y velocidad promedio de 3 nudos, en dos estratos de profundidad < 50 m y de 50 a 100 m. Se registró la información oceanográfica (temperatura en ºC y salinidad en ups - unidades practicas de salinidad) mediante un CTD Sea Bird Electronics SBE y se determinó la profundidad con una ecosonda científica SIMRAD EY500. En cada estación se tomaron muestras de sedimento con una draga Van Veen, con una cobertura de área de 0,07 m2. Se estableció el tipo de sustrato en cada estación, a través del valor phi medio (tamaño medio del grano) para cada una de las muestras. Los valores de phi medio de -1 a 0 equivalen a arena muy gruesa, de 0 a 1 arena gruesa, de 1 a 2 arena mediana, de 2 a 3 arena fina, de 3 a 4 arena muy fina, y los valores mayores de 4 a los lodos (Buchanan, 1984, Benavides, 2007). Se registró la posición geográfica, (Longitud y Latitud) de cada lance, longitud total de los peces (LT), estado gonadal (EG) peso total (PT) y sexo. Se utilizó el software MIX (Mixture analysis) (MacDonald y Pitcher, 1979) para analizar el numero de datos de frecuencia de tallas (LT) de E. argenteus. Este análisis considera los datos de frecuencia de tallas como una mezcla de funciones de densidad de probabilidad (pdf), cuyas modas dependen de una combinación de distancias entre las tallas medias, las magnitudes de las varianzas y la proporción del número de peces en cada moda y el tamaño de muestra total. Se utilizó el algoritmo quasi-Newton para ajustar los datos de frecuencia de tallas para obtener las medias asumiendo una distribución normal. El enfoque usado fue minimizar el chi-cuadrado y obtener un valor p en el cual las distribuciones observadas y estimadas no fueran significativamente diferentes. Para determinar si existen diferencias significativas entre las medias por sexo se utilizó un t-test de Student (Gotelli y Ellison, 2004, Manly, 2004).
Se establecieron cuatro estados de madurez macroscópicos I y II inmaduros, III y IV maduros. La talla a la madurez sexual (LT50%) se modeló ajustando la función logística con la proporción de hembras maduras con 5 mm de intervalo de longitud total (LT). La curva fue ajustada por mínimos cuadrados para obtener los parámetros del modelo logístico.
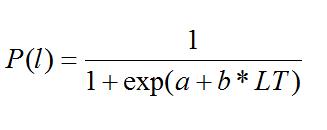
Donde P(l) es la proporción de hembras maduras, a y b son los parámetros estimados. La talla al 50% de madurez es LT50%= -(a/b) (King 2007).
Se calculó mediante el método de área barrida (King, 2007), el valor de la Captura por Unidad de Área (CPUA) en kg/km2 y los peces se separaron en juveniles y adultos de acuerdo a la talla media de madurez. La distribución espacial de la abundancia y estructura de tallas se modeló mediante geoestadística. Por lo tanto, se realizó una modelación espacial de la abundancia y estructura de tallas de Eucinostomus argenteus. Para analizar la autocorrelación se calculó un variograma experimental , donde h representa la distancia entre estaciones de muestreo (Cressie, 1993).
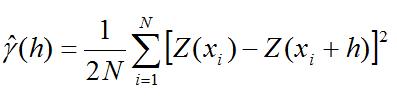
N es el número de pares de datos separados por una distancia h (Conan, 1985). El modelo de variograma que mejor se ajustó a los datos fue el esférico, una vez comprobado otros modelos (e.g. exponencial, gaussiano) mediante mínimos cuadrados ponderados. El modelo esférico con rango r y sill C, es igual a:
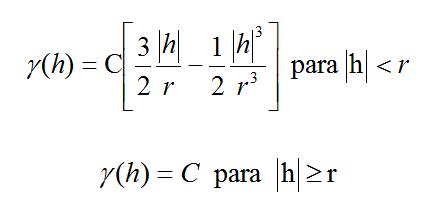
El modelo de variograma obtenido se utilizó junto con los datos del muestreo para calcular las ponderaciones óptimas atribuibles en cada estación y para estimar la densidad y estructura de tallas de E. argenteus (Z*) en localidades no muestreadas (interpolación mediante modelación espacial, Kriging).
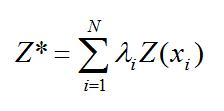
Donde N es el número de muestras, λi es la ponderación atribuida a la muestra xi, y Σλi = 1. Los N ponderadores λi se calculan para asegurar que el estimador sea insesgado y que la varianza de estimación sea mínima (Journel y Huijbregts, 1978, Petitgas, 1996).
La relación entre la abundancia y estructura de tallas con las variables ambientales fue evaluada con modelos GAM. Los Modelos Aditivos Generalizados (GAM, por sus siglas en ingles, Generalized Additive Modeling) (Hastie y Tibshirani, 1990) son una técnica moderna de estadística no paramétrica que libera al investigador del concepto limitante de una estricta forma paramétrica y permite el ajuste de modelos estadísticos que están de acuerdo con la teoría ecológica (Katsanevakis y Maravelias, 2009). Los GAM se están usando cada vez más para estudiar la distribución espacial de las especies marinas y su relación con el hábitat (Maravelias y Papaconstantinov, 2003, Katsanevakis, 2007, Maravelias et ál., 2007 a,b) y en la interpretación de estudios sobre ecología marina (Katsanevakis y Maravelias, 2009). En este estudio se hizo un análisis de las preferencias ambientales de la abundancia y estructura de tallas de E. argenteus y los predictores (temperatura, salinidad, tipo de sedimento, profundidad) mediante GAM (Hastie y Tibshirani, 1990). Un modelo aditivo es una extensión de los modelos lineales, pero permite que las funciones lineales de los predictores (temperatura, salinidad, tipo de sedimento y profundidad) sean reemplazados por funciones de suavizamiento (Agenbag et ál., 2003).
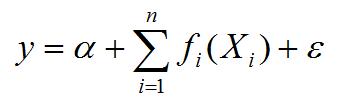
Donde y es la respuesta, Xi son los predictores,α es una constante y ε es el error. Los ƒi son estimados usando suavizadores. En este estudio se utilizó un suavizador spline (s) para estimar estas funciones no paramétricas y una familia Gausiana. El procedimiento de diagnóstico del GAM incluyó el valor de significancia (p) y el cálculo del porcentaje de devianza explicada por el modelo. La devianza es similar a la varianza y la devianza nula es similar a la varianza total. Por lo tanto, la devianza nula menos la devianza residual es la varianza explicada por el modelo (Stoner et ál., 2001).
También se utilizó el criterio de información Akaike (AIC), que es una medida de la devianza del modelo corregida para el número de predictores y se escoge el modelo con el AIC más bajo (Burnham y Anderson, 2002).
RESULTADOS
La Temperatura Superficial del Mar (TSM) (Fig. 2a) fue menor en el norte del área de estudio, entre Manaure y Puerto Estrella, con dos núcleos de surgencia: uno en el Cabo de la Vela y otro en Punta Gallinas. En el sur la TSM aumentó hacia la costa, (> 27ºC) con un pequeño núcleo de aguas más frías entre Dibulla y el río Palomino. La temperatura cercana al fondo (Fig. 2b) fue baja y homogénea en toda el área de estudio (21,4 - 23,0 °C). En el Cabo de la Vela los registros fueron menores comparados con los registros del sur, hasta Palomino. Sin embargo, tanto en la superficie como cerca al fondo, en boca Camarones se encontraron mayores temperaturas, debido a la presencia de una termoeléctrica que descarga las aguas de refrigeración (calientes) directamente al mar.
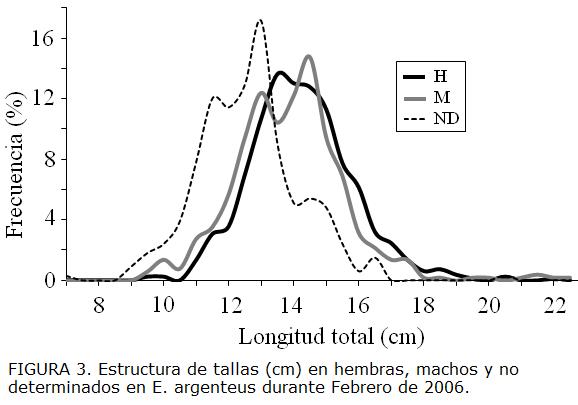
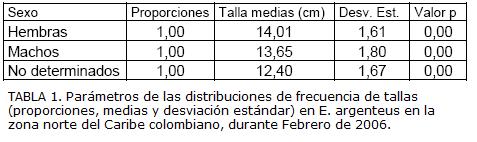
La estructura de tallas de E. argenteus estuvo comprendida entre 7,0 cm y 22,5cm de LT (Fig. 3) y se caracterizó por presentar una distribución normal para machos (n = 509 individuos), hembras (n = 814 individuos) y no determinados (n = 332 individuos) (Kolmogorov-Smirnov: p>0,05). La comparación de la estructura de tallas entre machos, hembras e indeterminados permitió comprobar que existen diferencias significativas (t-test de Student, p = 0,00), los individuos indeterminados presentaron las tallas menores, mientras las hembras presentaron las tallas mayores (Tabla 1).
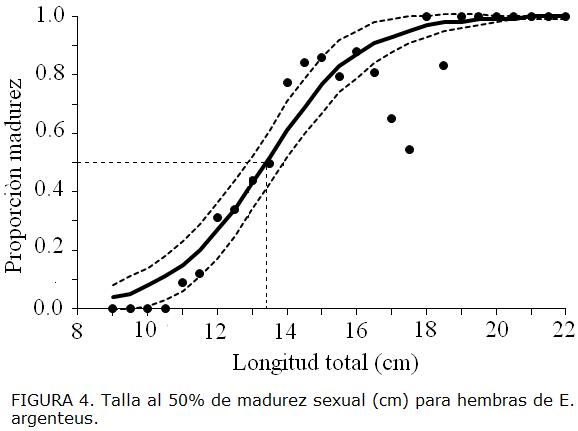
Las hembras mostraron un 35,51% de individuos inmaduros y 64,49% de maduros. La talla al 50% de madurez (TL50%) en hembras fue 13,38 (cm) TL (I.C. 95% inferior = 12,88, I.C. superior = 13,90), los parámetros a = 9,84 y b= -0,73 (Fig. 4).
La distribución espacial de la CPUA de E. argenteus, estudiado a través de un variograma isotrópico experimental y modelado por un modelo de variograma esférico (Fig. 5), mostró una clara estructura espacial, con un 31,1% de la varianza no explicada por el diseño de muestreo (nugget) y el rango de correlación fue de 58 km. La distribución espacial de E. argenteus mostró dos zonas con mayor agregación: una entre Riohacha y Punta Gallinas, la cual presentó dos núcleos de abundancia alta, uno frente a Manaure y el otro frente al Cabo de la Vela y la otra agregación se presentó cerca de la costa entre Boca Camarones y el río Buritaca (Fig. 6).

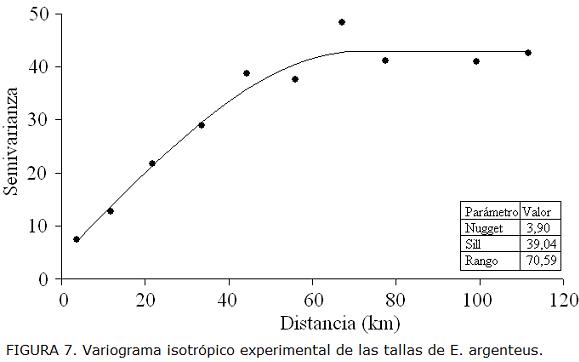
En el análisis espacial de las tallas, el rango de correlación fue de 70,59 km. A cortas distancias el variograma mostró una estructura no resuelta por el diseño de muestreo que consideró solo un 3,9% de la varianza total (Fig. 7). Se utilizó como criterio de peces juveniles y adultos la talla media de madurez sexual (juveniles < 13,38 cm > adultos). La distribución espacial de las tallas de E. argenteus mostró que los peces adultos (talla media 14,01 cm) se encontraron principalmente en la zona norte del área de estudio desde Manaure hasta Punta Gallinas, mientras que los peces juveniles (talla media 12,40 cm) se distribuyeron al sur del área de estudio entre Dibulla y el río Palomino (Fig. 8.
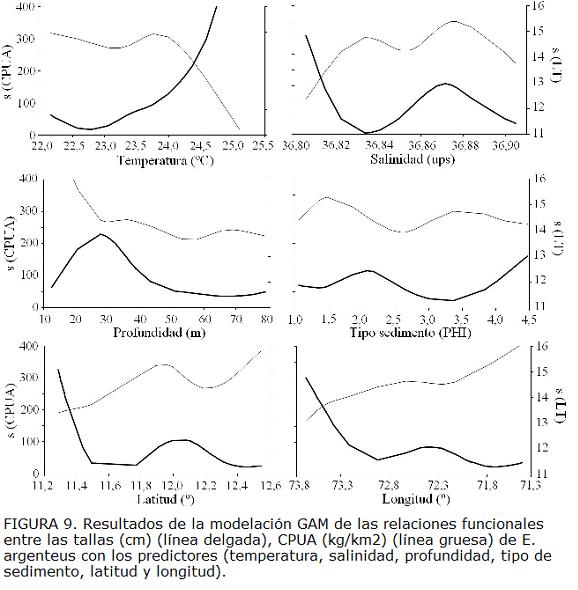
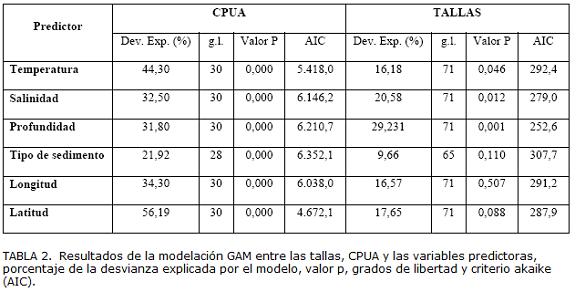
Los análisis de las relaciones entre la CPUA y las tallas con los predictores ambientales (temperatura, salinidad, tipo de sedimento y profundidad) mostraron asociaciones significativas. La temperatura y la profundidad fueron las variables que presentaron mayor porcentaje de la varianza explicada (Tabla 2). Las abundancias mayores estuvieron asociadas con temperatura mayor a 23,5 °C y en 30 m de profundidad (Fig. 9). La distribución espacial de las tallas estuvo asociada significativamente con temperatura, salinidad y profundidad. Se observó que las tallas menores se encuentran asociadas con temperaturas mayores de 24 ºC mientras que las tallas mayores (adultos) con temperaturas menores de 24 ºC (Fig. 9). Las tallas mayores se encontraron a profundidades menores de 30 m y, las tallas menores se encontraron asociados a profundidades mayores de 30 m. La temperatura y la profundidad fueron las variables que predijeron mejor la distribución espacial de la abundancia de E. argenteus (Fig. 9) y la profundidad fue la variable predictora que explicó mejor la distribución espacial de las tallas de E. argenteus (Tabla 2). Los valores de CPUA se encontraron relacionados (p<0,05) (Tabla 2) con la latitud y longitud, ya que las mayores abundancias se presentaron hacia el oeste y hacia el sur del área de estudio, representada por las tallas menores (Fig. 9).
DISCUSIÓN
Eucinostomus argenteus es una especie ampliamente distribuida en la zona norte del Caribe colombiano durante la época seca, apareciendo en 33 de las 48 estaciones establecidas en toda el área de estudio. Lo anterior confirma las observaciones de Manjarrés (1998), Manjarrés et ál. (2001) y Benavides (2007), quienes mencionan a E. argenteus como una especie dominante en número y biomasa disponible en el Caribe colombiano. En general, E. argenteus ha sido considerada como una especie dominante en el componente ecológico marino-eurihalino que conforma la estructura de la comunidad íctica en diferentes áreas tropicales estudiadas (Saul y Cunningham, 1995, Godefroid et ál., 2001, Ordóñez-López y García-Hernández, 2005, Parra et ál., 2007). Los individuos maduros y de tallas mayores que se encontraron distribuidos principalmente en el sector comprendido entre Manaure y Punta Gallinas, zona influenciada por el evento de surgencia que se presenta en La Guajira en la época seca y se caracteriza por aguas frías y de gran productividad. Estos factores ambientales están asociados a las distribución espacio - temporal de huevos y larvas de los peces, el estado de madurez gonadal, el desove y son importantes en la determinación de los patrones de crecimiento larval, jugando así un papel importante en el sustento de las pesquerías (Andrade y Barton, 2005). Los peces juveniles que se distribuyeron al sur del área de estudio, entre Boca Camarones y el Río Buritaca, sector caracterizado por temperaturas más altas, una zona de mezcla de las aguas provenientes de los ríos, ricas en nutrientes y presenta ecosistemas de manglar y pastos marinos (Díaz -Pulido, 1997, INVEMAR, 2000), los cuales, son apropiados para el desarrollo de las etapas iniciales del ciclo de vida de las especies. Lo anterior ha sido observado en otros sistemas costeros (Ordóñez-López y García-Hernández, 2005, Godefroid et ál., 2001), en los cuales, se asocia la presencia de peces juveniles en áreas de manglares y arrecifes coralinos a las condiciones de refugio y disponibilidad de alimento que ofrecen estos ecosistemas costeros. Las condiciones ambientales se consideran importantes en la distribución y abundancia de los peces del Caribe colombiano (Paramo et ál., 2003, Paramo y Roa, 2003). Producto del evento de surgencia que se presenta en el sector de La Guajira, la temperatura superficial del mar disminuye. Durante el periodo seco, la corriente del Caribe se desplaza hacia el oeste con velocidades que alcanzan más de un nudo en ciertos sectores (Pujos et ál., 1986).
Esta corriente, podría trasladar aguas frías y ricas en nutrientes provenientes de la zona de surgencia de La Guajira (norte del área de estudio) hacia el sur del área de estudio (Paramo, 2007). Es probable que la surgencia traslade los huevos y larvas hacia el área comprendida entre Boca Camarones y el Río Buritaca, donde se encontró un atrapamiento de aguas cálidas hacia la costa, donde se pueden encontrar las condiciones de enriquecimiento, concentración y retención favorables para el reclutamiento (“tríada de Bakun”, Bakun, 1996), que podría considerarse como un área de crianza para peces demersales y pelágicos pequeños (Paramo et ál., 2003, Paramo, 2007, Paramo et ál., 2009). Manjarrés et ál. (2001) describen a la profundidad como una de las variables que explican mejor la variación estacional de las asociaciones de peces demersales en el Caribe colombiano, siendo las agregaciones de E. argenteus una de las más persistentes inter-anualmente durante el periodo de estudio (1995 -1996). Debido a que E. argenteus se encontró asociado principalmente a profundidades entre los 20 y 40 m y se evidencia su proximidad a la costa, tanto en sus estados juveniles como adultos. Así, la abundancia en escala espacial (Ayala-Pérez et ál., 2001) y las diferencias en la estructura de talla, permiten argumentar las preferencias de uso del hábitat. Es evidente que existe una segregación espacial en E. argenteus que está relacionada con condiciones del hábitat tales como la temperatura del agua y la profundidad que caracterizan las diferentes condiciones del hábitat y que favorecen el desarrollo de cada una de los etapas del ciclo de vida de la especie.
AGRADECIMIENTOS
Los autores agradecen al Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología “Francisco José de Caldas” (COLCIENCIAS), a la Universidad del Magdalena, al Instituto Colombiano de Desarrollo Rural (INCODER), al Instituto de Investigaciones Marinas y Costeras “José Benito Vives de Andréis” (INVEMAR), al Institut de Recherche pour le Developpment, France (IRD), por el apoyo financiero, técnico y logístico para el desarrollo del proyecto “Las Áreas Marinas Protegidas (AMPs): Una Herramienta de Manejo para las Pesquerías Demersales en la Zona Norte del Caribe Colombiano” código 020309-16652. Este trabajo fue parte de la tesis del primer autor como requisito para obtener el grado de Ingeniera Pesquera de la Universidad del Magdalena (Colombia).
BIBLIOGRAFIA
ACOSTA O. Algunos aspectos de vida de la familia Gerreidae en le Ciénaga de Tesca, Cartagena. [Tesis de Biólogo Marino]. Bogotá: Facultad de Ciencias del Mar, Universidad Jorge Tadeo Lozano; 1985.
[ Links ]AGENBAG GJJ, RICHARDSON AJ, DEMARCQ H, FRÉON P, WEEKS S, SHILLINGTON FA. Estimating environmental preferences of South African pelagic fish species using catch size and remote sensing data. Prog. Oceanog. 2003; 59: 275 - 300
[ Links ]ANDRADE CA, BARTON ED, MOOERS CHNK. Evidence for an eastward flow along the Central and South American Caribbean Coast. J. Geoph. Res. 2003; 108: 1-11.
[ Links ]ANDRADE C, BARTON ED. The Guajira upwelling system. Continental Shelf Research. . 2005; 25: 1003 - 1022.
[ Links ]ARENAS P. Alimentación y relaciones tróficas de las mojarras de la Ciénaga Grande de Santa Marta, Caribe colombiano (Pisces: Perciformes, Gerreidae). [Tesis de Maestría]. Santa Marta: Departamento de Biología. Universidad Nacional de Colombia; 1990.
[ Links ]ARENAS P, ACERO A. Organización trófica de las mojarras (Pisces: Gerreidae) de la Ciénaga Grande de Santa Marta (Caribe colombiano). Rev. Biol. Trop. 1992; 40 (3): 287 - 302.
[ Links ]AYALA-PÉREZ L, GOMEZ-MONTEZ B, MIRANDA-RAMOS J. Distribución, abundancia y parámetros poblacionales de la mojarra Diapterus rhombeus (Pisces: Gerreidae) en la Laguna de Términos, Campeche, México. Rev. Biol. Trop. 2001; 49 (2): 635-642.
[ Links ]BAKUN A. Patterns in the ocean: ocean processes and marine population dynamics. University of California Sea Grant, UCSD, San Diego, CA y Centro de Investigaciones Biológicas de Noroeste, La Paz, Baja California. 1996.
[ Links ]BENAVIDES S. Ensamblaje de peces demersales en la costa norte del caribe colombiano durante la época seca: un enfoque espacial para el manejo de los recursos pesqueros. [Tesis Ingeniería Pesquera]. Santa Marta: Facultad de ingeniera,Universidad del Magdalena; 2007.
[ Links ]BIANCHI G, SKJOLDAL H.R, editores. The ecosystem approach to fisheries. Rome: CABI FAO; 2009.
[ Links ]BUCHANAN J. Sediment analysis, En: Holme NA, McIntyre AD, editors. Methods for the study of marine benthos. Blackwell. London; 1984. p. 41-45.
Bula-Meyer G. Oceanografía. En: Reichel-Dolmatoff G. (Editor). Caribe Colombia. FONDO FEN-COLOMBIA, Bogotá. 1990; p.101-113. [ Links ] [ Links ]BURNHAM K, ANDERSON D. Model selection and multimodel inference: a practical information-theoretic approch. 2 ed. New York: Springer-Verlag.; 2002.
[ Links ]CONAN G. Assessment of shellfish stocks by geostatistical techniques. ICES CM 1985/K: 30.
[ Links ]CRESSIE N. Statistics for spatial data. New York: John Wiley and Sons. 1993.
[ Links ]DE GRADO A, BASHIRULLAH A. Algunos atributos de la estructura comunitaria de la ictiofauna de la laguna grande de obispo, golfo De Cariaco. Venezuela. Instituto Oceanográfico de Venezuela, Universidad de Oriente. Biología Marina; 2001.
[ Links ]DÍAZ-PULIDO G. Ecosistemas Marinos y Costeros. En: Chávez ME, Arango N, editores. Informe nacional sobre el estado de la Biodiversidad Colombia. Tomo I, Diversidad Biologica. Instiuto de Invetigaciones de Recusrsos Biologicos Alexander von Humboldt; 1997. p. 228-314.
[ Links ]FOLMER O, PENNINGTON M A. Statistical evaluation of the design and precision of the shrimp trawl survey off West Greeland. Fish. Res. 2000; 49: 165 - 178.
[ Links ]GODEFROID R, SANTOS C, HOFSTAETTER M, SPACH L. Occurrence of Larvae and Juveniles of Eucinostomusargenteus, Eucinostomus gula, Menticirrhus americanus,Menticirrhus littoralis, Umbrina coroides and Micropogoniasfurnieri at Pontal do Sul beach. Paraná Braz. Arch. Biol. Tech. 2001; 44 (4): 411-418.
[ Links ]GOTELLI NJ, ELLISON AM. A Primer of Ecological statistics. USA. Sinauer Associates.; 2004.
[ Links ]HASTIE T, TIBSHIRANI R. Generalized Additive Models. 1 ed. London: Chapman and Hall.; 1990.
[ Links ]INVEMAR. Programa nacional de investigación en biodiversidad marina. Ecorregiones naturales marinas y costeras. INVEMAR; 2000.
[ Links ]JOURNEL A, HUIJBREGTS C. Mining geostatistics. New York: Academic Press.; 1978.
[ Links ]KATSANEVAKIS S. Density surface modeling with line transect sampling as a tool for abundance estimation of marine benthic species: the Pinna nobilis example in a marine lake. Mar. Biol. 2007; 152: 77-85.
[ Links ]KATSANEVAKIS S, MARAVELIAS CH D. Bathymetric distribution of demersal fish in the Aegean and Ionian Seas based on generalized additive modelling. Fish. Sci. 2009; 75: 13-23.
[ Links ]KING M. Fisheries biology, assessment and management. 2 ed. London: Wiley-Blackwell.; 2007.
[ Links ]MACDONALD PD, PITCHER TJ. Age-groups from size-frecuency data: a versatile and efficient method of analyzing distribution mixtures. J. Fish. Res. Board Can. 1979; 36:987-1001.
[ Links ]MANJARRÉS L. Caracterización ambiental, ecológica y biológico-pesquera de las asociaciones de peces demersales del Caribe colombiano norte, con énfasis en pargos (Lutjanidae). [Tesis de Maestría]. Santa Marta: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 1998.
[ Links ]MANJARRÉS L, GARCÍA C, ACERO A. Caracterización ecológica de las asociaciones de peces demersales del Caribe colombiano norte, con énfasis en los pargos (Lutjanidae). Bol. Inv. Mar. Cost. 2001; 30: 77 - 107.
[ Links ]MANLY BFJ. Multivariate Statistical Methods: A Primer. Third Edition. London. Chapman & Hall.; 2004.
MARAVELIAS CD, PAPACONSTANTINOU C. Size-related habitat use, aggregation patterns and abundance of anglerfish (Lophius budegassa) in the Mediterranean Sea determined by generalizad additive modelling. J. Mar. Biol. Assoc. UK. 2003; 83:1171-1178 [ Links ] [ Links ]MARAVELIAS CD, TSITSIKA EV, PAPACONSTANTINOU C. Seasonal dynamics, environmental preferences and habitat selection of John Dory (Zeus faber). Est. Coast. Shelf. Sci. 2007a; 72(4):703-710.
[ Links ]MARAVELIAS CD, TSITSIKA EV, PAPACONSTANTINOU C. Evidence of Morocco dentex (Dentex maroccanus) distribution in the NE Mediterranean and relationships with environmental factors determined by generalized additive modelling. Fish.Ocean. 2007b; 16: 294-302
[ Links ]NAVAJAS P, GARCÍA C. Aspectos tróficos de cinco especies de peces de la familia Gerreidae (Pisces) en el Golfo de Salamanca, Caribe colombiano. En: Proyecto Estudio ecológico pesquero de los recursos demersales del Golfo de Salamanca, Caribe colombiano. Estimación de la variabilidad de los componentes biológicos del sistema; 1999.
[ Links ]ORDOÑEZ-LÓPEZ U, GARCÍA-HERNÁNDEZ V. Ictiofauna juvenil asociada a Thalassia testudinum en Laguna Yalahau, Quintana Roo. Hidrobiologia. 2005; 15 (2): 195-204.
[ Links ]PARAMO J, ROA R. Acoustic-geostatistical assessment and habitat-abundance relations of small pelagic fish from the Colombian Caribbean. Fish. Res. 2003; 60: 309-319.
[ Links ]PARAMO J, QUIÑONES R, RAMIREZ A, WIFF R. Relationship between abundance of small pelagic fishes and environmental factors in the Colombian Caribbean Sea: an analysis based on hydroacustic information. Aquat. Liv. Res. 2003; 16(3): 239 - 245.
[ Links ]PARAMO J. AMCP: herramienta de manejo para las pesquerías en el norte del Caribe colombiano. En: de Andrade R, editor. Áreas Marinas y Costeras Protegidas de Múltiples Usos. Ocho Libros editores. GEF Marino - PNUD; 2007. p. 197 - 199.
[ Links ]PARAMO J, GUILLOT L, BENAVIDES S, RODRÍGUEZ A, SÁNCHEZ C. Aspectos poblacionales y ecológicos de peces demersales de la zona norte del Caribe colombiano en relación con el hábitat: una herramienta para identificar Áreas Marinas Protegidas (AMPs) para el manejo pesquero. CALDASIA. 2009; 31(1): 123-144.
[ Links ]PARRA B, RUIZ LJ, PRIETO A. Índices ecológicos y parámetros biométricos de Haemulidae (Pisces: Perciformes) en la zona costera de la Isla de Cubagua, Venezuela. Zootecnia Tropical. 2007; 25(1): 51-61.
[ Links ]PETITGAS P. Geostatistics and their applications to fisheries survey data. En: Megrey BA, Moskness E, editors. Computers in fisheries research. London: Chapman and Hall; 1996. p. 113-142.
[ Links ]PUJOS M, PLAGLIARDINI J, STEER R, VERNETTE G, WEVER O. Influencia de la contracorriente norte colombiana para la circulación de las aguas en la plataforma continental: su acción sobre la dispersión de los efluentes en suspensión del río Magdalena. Bol. Cient. CIOH. 1986; 6: 3-15.
[ Links ]RIVOIRARD J, SIMMONDS J, FOOTE KG, FERNANDES P, BEZ N. Geostatistics for Estimating Fish Abundance. 1 ed. London: Blackwell Science.; 2000.
[ Links ]RODRIGUEZ V. Contribución al conocimiento biológico de las mojarras (Pisces: Gerreidae) de la Ciénaga de la Virgen. Tesis de Biólogo Marino. Universidad Jorge Tadeo Lozano. Facultad de Ciencias del Mar; 1982.
[ Links ]ROSSI R, MULLA D, JOURNEL A, FRANZ E. Geostatistical tools for modeling and interpreting ecological spatial dependence. Ecol. Monog. 1992; 62(2): 277-314.
[ Links ]RUEDA M, SANTOS-MARTINEZ A. Evaluación de los principales recursos pesqueros de la Ciénaga Grande de Santa Marta, costa Caribe colombiana. INVEMAR. 1998.
[ Links ]RUEDA M, DEFEO O. Linking fishery management and conservation in a tropical estuarine lagoon: biological and physical effects of an artisanal fishing gear. Estuar. Coast. Shelf Sci. 2003; 56: 935-942.
[ Links ]SAUL A, CUNNINGHAM P. Comunidade Ictiofaunística da Ilha do Bom Abrigo, Cananéia, São Paulo, Brasil. 2- Lanço. Arq. Biol. Tecnol. 1995; 38(4): 1053-1069.
[ Links ]SIMARD Y, MARCOTTE D, BOURGAULT G. Exploratión of geostatistical methods for mapping and estimating acoustic biomass of pelagic fish in the Gulf of St. Lawrence: size of echo-integration unit and auxiliary environmental variables. Aquat. Liv. Res. 1992; 6: 185-199.
[ Links ]STONER A, MANDERSON J, PESSUTII J. Spatially explicit analysis of stuarine habitat for juvenile winter flounder: combing generalized additive models and geographic information system. Mar. Ecol. Prog. Ser. 2001; 213: 253 - 271
[ Links ]VIANNA M, COSTA-NEVES F. Length-weight relationship of fish caught as by-catch by shrimp fishery in the southeastern coast of Brazil. B. Inst. Pesca, São Paulo. 2004; 30(1): 81 - 85.
[ Links ]
