










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.15 no.1 Bogotá Jan./Apr. 2010
RELACIÓN ENTRE ASOCIACIÓN MICORRÍCICA CON EL ESTADO FITOSANITARIO EN EL ARBOLADO URBANO DE BOGOTÁ D.C.
RELATIONSHIP BETWEEN MYCORRHIZAL ASSOCIATION WITH THE STATE WOODY PLANT HEALTH IN THE CITY OF BOGOTA DC
CAROLINA RAMOS MONTAÑO1, M.Sc. Ecología y Biología Evolutiva; RAÚL H. POSADA ALMANZA2, M.Sc. Microbiología; MIGUEL A. RONDEROS FIGUEROA3, Ing. Agroecólogo; GUSTAVO A. PENAGOS CAÑON4, Ing. Agroecólogo. 1Subdirección Científica, Jardín Botánico José Celestino Mutis. Correspondencia: Kra 28B No 64A-25 Aptos 302-402 Bogotá-Colombia. E-mail: charo.ramon@gmail.com . 1 Instituto de Ecología A.C. Xalapa - Veracruz- México. 3 Subdirección Científica, Jardín Botánico José Celestino Mutis. 4 Programa de Ingeniería Agroecológica, UNIMINUTO.
Presentado 23 de Diciembre 2008 Aceptado 20 marzo 2009 Correcciones 30 junio 2009
RESUMEN
En parques y zonas verdes de Bogotá DC., se evaluó la densidad de esporas y la colonización radical en la rizosfera de tres especies comunes del arbolado urbano: Eugenia myrtifolia, Ficus soatensis y Croton bogotensis, con el fin de determinar si el grado de micorrización está relacionado con el estado fitosanitario y si existe diferencias en el grado de micorrización entre dos zonas climáticas distintas. Se comprobó que la especie y el clima son fuentes significativas de variabilidad en el estado general de micorrización. Eugenia myrtifolia presentó la mayor colonización radical promedio pero el más bajo promedio de densidad de esporas, mientras C. bogotensis tuvo la respuesta contraria. En general los ambientes secos y semisecos favorecieron la micorrización. Considerando los datos de todas especies, se comprobó una relación positiva entre el estado fitosanitario general y la colonización radicular, y al evaluar la relación con la afectación de síntomas específicos se encontró que la clorosis, la quemazón y la herbivoría mantienen una relación negativa con la micorrización en E. myrtifolia y C. bogotensis. Los resultados sugieren que la asociación con hongos de micorriza arbuscular ayuda de algún modo a reducir esos síntomas, presentando un potencial importante para mejorar la sanidad y vida media de las especies arbóreas en la ciudad.
Palabras clave.. Micorriza arbuscular, Eugenia myrtifolia, Ficus soatensis, Croton bogotensis, densidad de esporas, colonización radicular, síntomas foliares, zonas climáticas urbanas.
ABSTRACT
Spore number and root infection by arbuscular mycorrhizal fungi were evaluated in Eugenia myrtifolia, Ficus soatensis and Croton bogotensis, in parks and green zones of urban woodlands of Bogotá DC. The aim was to investigate relations between mycorrhizal associations and tree phyto-sanitary conditions, and effects of two distinct climatic zones. It was demonstrated that plant species and climate are significant sources of variations in the general mycorrhizal state. Eugenia myrtifolia showed the highest degree of root colonization but the lowest number of spores, while C. bogotensis had the opposite response. In general, dry environments favored the mycorrhizal infection levels. By considering overall data, there was a positive relation between the general phytosanitary status of the urban trees and the mycorrhizal colonization. The evaluation of the relationship with the incidence of specific foliar symptoms showed that chlorosis, bight and herbivory maintained a negative relation with the mycorrhization in E. myrtifolia and C. bogotensis. Results suggest that association with AM fungi helps in any way for reducing those symptoms, showing an important strategy to improve the sanity of urban trees.
Key words. Arbuscular mycorrhiza, Eugenia myrtifolia, Ficus soatensis, Croton bogotensis, spores density, root colonization, foliar symptoms, climatic urban zones.
INTRODUCCIÓN
Los hongos formadores de micorriza arbuscular (HMA) han sido objeto de un amplio interés científico y tecnológico, desde el descubrimiento de su capacidad de mejorar la nutrición, el estado fitosanitario y la resistencia a condiciones de estrés, estando presentes en más del 70% de las plantas superiores (Alexopoulos et al., 1996; Cairney, 2000; Smith y Read, 2008). La absorción de bioelementos escasamente disponibles en el suelo como el fósforo, y metales traza de baja movilidad como el cobre y el zinc, aumentan notablemente en plantas micorrizadas (Taiz y Zeiger, 1998; Plenchett et al., 2005; Audet y Charest, 2006; Smith y Read, 2008). Sin embargo, el grado de micorrización depende de las condiciones nutricionales del suelo, la materia orgánica, el pH, la humedad (Clark, 1997; Shahri y Abbasi, 2005; Khanam et al., 2006) y de la especies de plantas que hacen parte de la asociación (Janos, 2007).
Existe evidencia de que la presencia de micorrizas arbusculares ayuda a reducir la afectación por enfermedades foliares y radicales (Azcón-Aguilar y Barea, 1997; Herrea et al., 2007; Khaosaad et al., 2007). Mediante un complejo sistema de señales bioquímicas, la raíz permite la colonización por HMA y éstos a su vez estimulan un sistema de defensa que reduce la invasión por hongos fitopatógenos latentes en el suelo (Zhao y Qi, 2008), controlándolos además mediante la ocupación de espacio colonizable (Bennett et al., 2006). En una revisión de estudios que evalúan la interacción entre patógenos y HMA, Borowicz (2001) determinó que si bien en un 16% de los casos los hongos patógenos afectan negativamente el desarrollo de HMA, éstos reducen a su vez en un 50% de los casos el crecimiento de hongos patógenos.
La densidad de esporas y el grado de colonización de HMA son afectados por las condiciones climáticas. Generalmente en ambientes secos se tiende a encontrar promedios de conteo de esporas mayores a los que presenta el mismo sitio en época de lluvias, mientras los ambientes más húmedos parecen favorecer la germinación de esporas y mantener un sistema de infección alta por mayor actividad fotositética de la planta (Guadarrama y Álvarez-Sánchez, 1999; Lugo y González-Maza, 2003; Escudero y Mendoza, 2005). La heterogeneidad ambiental a escalas de paisaje determina gradientes importantes en las condiciones del suelo, capaces de generar patrones diferenciales en la caracterización micorrícica (Camargo-Ricalde y Esperon-Rodriguez, 2005; Gehring et al., 2006; Posada et al., 2008).
Lastimosamente los ambientes urbanos no han despertado el mismo interés de la investigación ecológica en bosques nativos y agro-ecosistemas. La gran mayoría de los estudios relacionados con la problemática sanitaria y estrategias de control en el arbolado de Bogotá DC. han sido realizados por el Jardín Botánico José Celestino Mutis. En la ciudad más del 36% del arbolado urbano presenta un estado sanitario deficiente, con afecciones foliares como quemazón, clorosis, necrosis, manchado, deformación y herbivoría, que al estar en alta incidencia reducen significativamente la vida media del arbolado (JBJCM, 2008; Ramos, 2008). Dado que los síntomas pueden estar relacionados tanto con deficiencias de nutrientes como con la afectación por patógenos, cobra importancia determinar si en el ambiente urbano estos efectos pueden ser reducidos mediante la simbiosis con HMA.
El arbolado urbano, considerado como un componente de importancia fundamental en los procesos ecológicos que se dan en las ciudades (Beckett et al., 2000; Nowak, 2006; Wu, 2008), requiere de tratamientos que mejoren su condición nutricional y sanitaria. En este estudio se evaluó el grado de micorrización que presentan las especies Ficus soatensis Dugant (Moraceae), Eugenia myrtifolia Sims (Myrtaceae) y Croton bogotensis Cuatrecasas (Euphorbiaceae) en parques y zonas verdes de Bogotá en dos tipos de ambiente de acuerdo a la precipitación media. Se puso a prueba la hipótesis de que la densidad de esporas y la colonización radical están relacionadas con el estado sanitario y que a su vez habrá diferencias de colonización y densidad de esporas entre ambientes.
METODOLOGÍA
El estudio abarcó diversos parques y zonas verdes de la ciudad de Bogotá D.C. Esta ciudad alberga alrededor de 8 millones de habitantes, presenta un régimen de precipitación que varía entre 400 y 1.300 mm de precipitación anual, y promedios de temperatura entre 11 y 16 °C (DAMA, 2005). Los sitios de muestreo fueron ubicados en dos tipos de ambiente, teniendo en cuenta el mapa de distribución de precipitación del DAMA (2007): Zonas secas y semisecas (precipitación media < 750 mm), y zonas húmedas y subhúmedas (precipitación media > 850 mm).
El muestreo incluyó parques y amplias zonas verdes en tres localidades de tendencia climática seca (Suba, Rafael Uribe Uribe, Kennedy) y cuatro localidades de tendencia climática húmeda (Barrios Unidos, Engativá, Teusaquillo y Usaquén), abarcando un importante cubrimiento de la ciudad de Bogotá (Figura 1). Se usó este tipo de sitios para trabajar con individuos cuya expansión radical no estuviera restringida por emplazamientos artificiales de materia dura. El trabajo de campo consistió en la valoración general (%) del estado sanitario mediante observación directa de la afección por diferentes síntomas en la fitomasa total de cada individuo: clorosis, necrosis, deformidad foliar, herbivoría, quemazón, puntos de succión, manchado y defoliación. El estimador de sanidad general (% San) fue obtenido restando a un 100% los respectivos porcentajes de afectación de los diversos síntomas evaluados. El estudio incluyó un total de 49 individuos.
Se tomó muestras de suelo a 5-10 cm de profundidad, en cuatro puntos ortogonalmente ubicados alrededor del árbol, a máximo 1,5 m de distancia del tronco. Las muestras fueron completamente homogenizadas en una sola muestra compuesta de suelo por individuo. La fase de laboratorio tuvo lugar en el laboratorio de Fisiología Vegetal del Jardín Botánico José Celestino Mutis. Se empleó una muestra de 15 g de suelo por individuo para extracción de esporas mediante el método de Gerdemann y Nicolson (1963), en un gradiente de sacarosa al 72% y el resultado se expresó como número de esporas por gramo de suelo seco. La humedad de la muestra fue estimada a partir de sub-muestras secadas hasta peso constante.
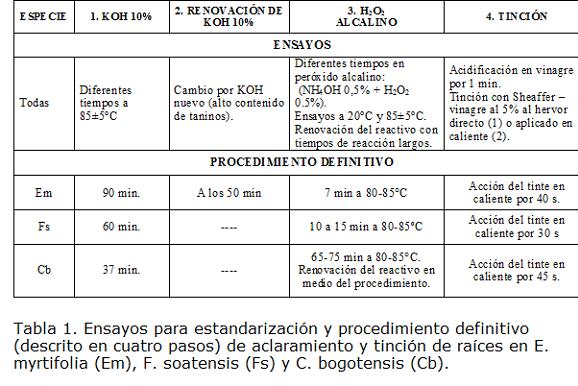
Para el proceso de aclaramiento y tinción de las raíces extraídas en las muestras de suelo, se recurrió a las propuestas de Kormanik y McGraw (1982) para aclaramiento y de Vierheilig et al. (1998) para la tinción. En términos generales, el método implementa el uso de peróxido alcalino para hacer más eficiente el proceso de aclaramiento de las raíces, y el uso de tinción vinagre - tinta Sheaffer (tinta de estilógrafo) en reemplazo del azul de tripano. Para determinar los tiempos óptimos de tinción para la observación óptima de estructuras de micorriza fue necesario realizar múltiples ensayos, modificando el tiempo de exposición a los reactivos para cada especie.
El carácter de los ensayos realizados y los protocolos finales estandarizados se presentan en la Tabla 1. En todos los casos fue preferible aplicar el tinte recién hervido sobre las raíces en lugar de hervirlas directamente en vinagre - Sheaffer. Una vez se obtuvo raíces micorrizadas de alto contraste (máximo aclaramiento del tejido radicular y máxima tinción de estructuras de HMA), los excesos de coloración fueron lavados con una mezcla de vinagre-agua microfiltrada al 50%. Las raíces fueron puestas en paralelo en portaobjetos y se procedió a la cuantificación microscópica por campos perpendiculares a las raíces; para reportar colonización se consideraron positivos campos con presencia ya sea de hifas, vesículas o arbúsculos. Se evaluaron 80 campos de observación por muestra, y se calcularon los porcentajes de colonización correspondientes.
Los datos de densidad de esporas (No. esporas / g) requirieron una transformación del tipo lnx + cosx para normalización. La densidad y la colonización (%) fueron analizadas mediante un ANDEVA multivariado a dos vías para determinar el efecto general del ambiente y la especie, y ANDEVAS a una vía para determinar los efectos sobre cada variable por separado. Un test Holm-Sidak determinó la diferencia de medias entre pares de comparación. Posteriormente se calculó coeficientes de correlación de Spearman (R) para evaluar la existencia de relación entre las variables de micorrización y el estado sanitario general (% San), y entre el porcentaje de humedad en el suelo y las variables de micorrización. Los análisis fueron hechos mediante los programas Statistica 6.0 (StatSoft Inc., 2001) y Sigma Stat 3.1 (Systat Software Inc., 2004).
RESULTADOS
El ANDEVA multivariado mostró que existe un efecto significativo de la especie (F4,84=13,134; P< 0,001) y el ambiente (F2,42=4,588; P=0,016) sobre la micorrización. Los efectos específicos sobre la densidad de esporas y la colonización se describen en la Tabla 2 y se observan en la Fig. 2.
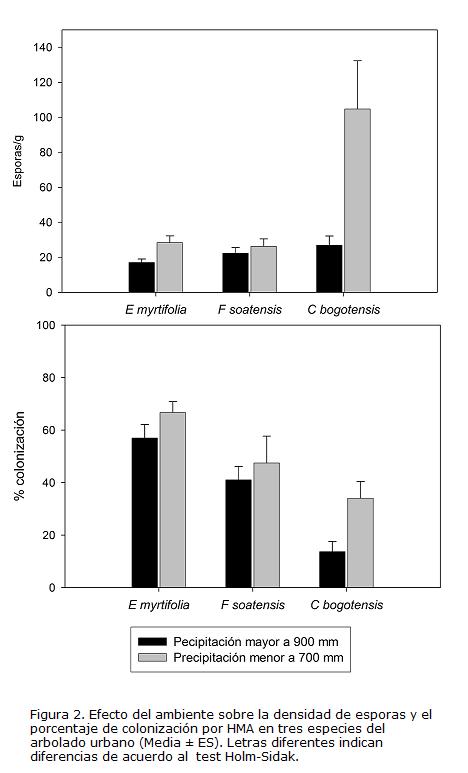
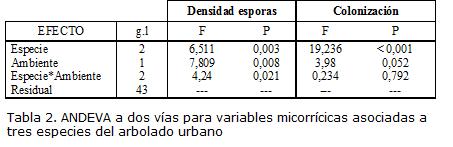
La colonización promedio fue mayor en E. myrtifolia (62% con variación entre 54,5 - 69,2 %) que en F. soatensis (44% variación entre 32,4 - 56,2 %) y C. bogotensis (23% con variación entre 14,0 - 32,6 %). En todas las especies los porcentajes de colonización tendieron a ser mayores en ambientes secos (28,6 - 75,4 %) que en ambientes húmedos de la ciudad (17,1 - 27,2 %), con una diferencia de medias marginalmente significativa (F1,43=3,99; p=0,052. Fig. 2). La densidad promedio de esporas fue mayor en C. bogotensis (64 esporas con variación entre 29 - 98 esporas/g) que en E. myrtifolia (23 con variación entre 17 - 28 esporas/g) y F. soatensis (24 esporas con variación 19 - 30 esporas/g). La diferencia entre ambientes en la densidad de esporas, fue significativa para C. bogotensis y E. myrtifolia (Fig. 2). La especie C. bogotensis presentó la mayor variabilidad en la micorrización: aunque hubo muestras sin colonización radical, también se encontró casos hasta del 63%, de colonización, particularmente pertenecientes al parque San Andrés en zona semiseca de la ciudad
Al correlacionar las variables micorrícicas (densidad de esporas y colonización radicular) con el estado fitosanitario, ninguna especie mostró una correlación clara entre la colonización radicular y % sanidad (Fig. 3); E. myrtifolia mostró una relación positiva, sólo marginalmente significativa. Indudablemente la colonización radicular, sin discriminar por especie, se correlacionó positiva y significativamente con el estado fitosanitario foliar general (R=0,343, P=0,016), a diferencia de la densidad de esporas, con la cual no hubo ninguna relación (R=0,106, P=0,466).
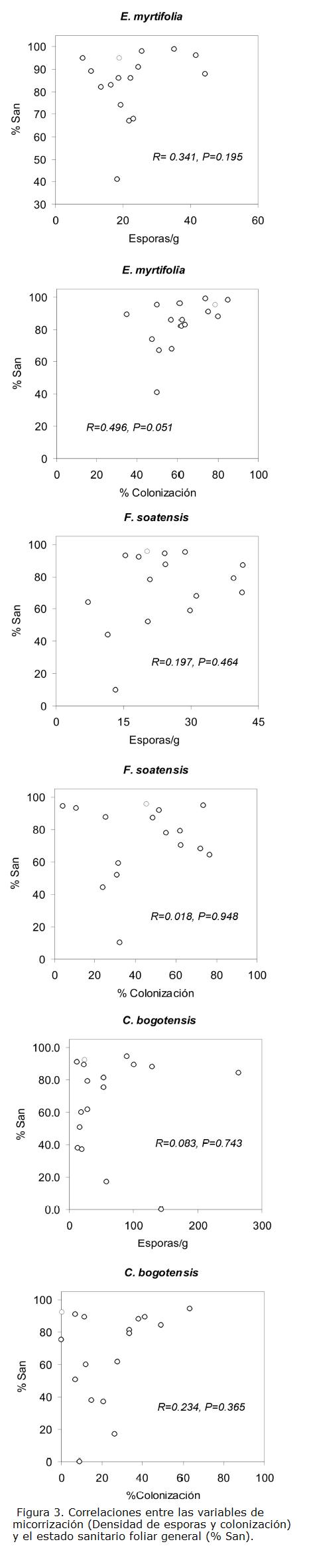
La densidad de esporas y la colonización radicular estuvieron positiva y significativamente relacionados en las especies E. myrtifolia y C. bogotensis. La humedad del suelo no mostró una relación importante con la micorrización, a excepción de una correlación positiva y significativa con la colonización radical en E. myrtifolia (Tabla 3).
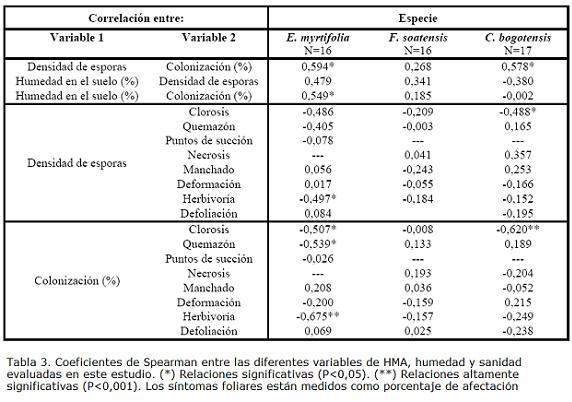
En cuanto a los síntomas sanitarios evaluados, clorosis y quemazón abarcaron en conjunto más del 40% de la afectación total en E. myrtifolia y F. soatensis, mientras la defoliación fue el síntoma más común en C. bogotensis (casi un 30% de la afectación). Otro síntoma común fue la herbivoría, que representó entre el 13 y el 21% de la problemática sanitaria de las especies. La lesión menos frecuente fue la necrosis, con un 5% de la afectación relativa (Fig. 4).
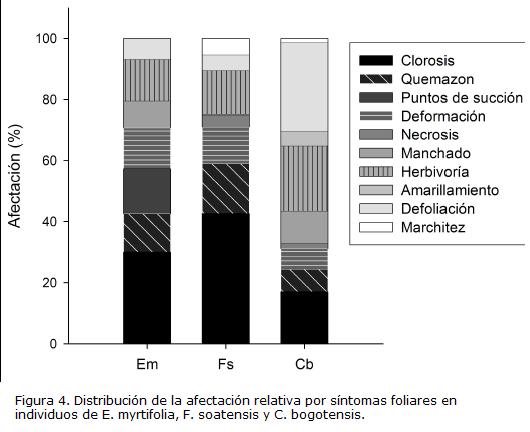
Los coeficientes de correlación calculados entre la micorrización y el porcentaje de afectación de cada síntoma sanitario se muestran en la Tabla 3. En la mayoría de los casos, el análisis dio lugar a valores negativos, y hubo una tendencia a que valores altos de correlación con la colonización también lo fueron con la densidad de esporas. En E. myrtifolia, clorosis, quemazón y herbivoría estuvieron negativamente relacionadas con el porcentaje de colonización; adicionalmente la herbivoría también estuvo negativamente correlacionada con la densidad de esporas. En C. bogotensis, las dos variables de HMA estuvieron negativamente relacionadas con la afectación por clorosis. La especie F. soatensis no presentó ninguna relación significativa.
DISCUSION
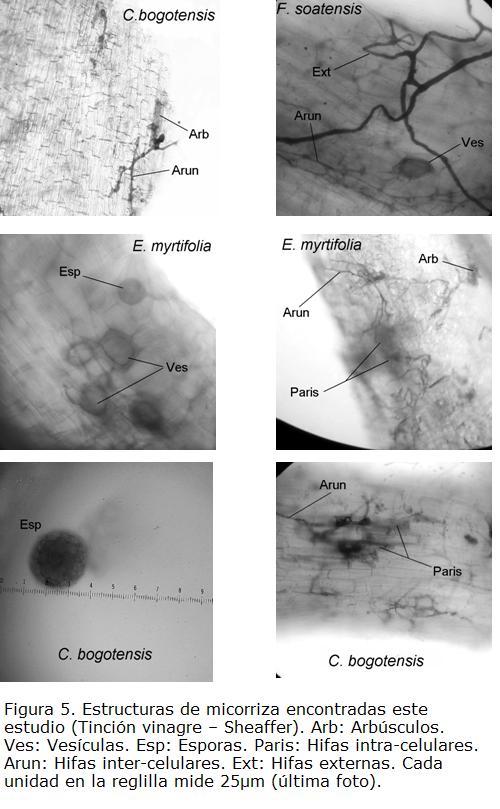
Tres de las especies más comunes en el arbolado urbano de Bogotá presentan respuestas de asociación micorrícica distinta (ver documentación fotográfica en Fig. 5). Las tres especies presentaron colonización por HMA en secuencia E. myrtifolia › F. soatensis › C. bogotensis. . Sin embargo, la densidad de esporas mostró un comportamiento diferente: C. bogotensis › E. myrtifolia = F. soatensis. Los resultados parecen seguir un patrón en función de la capacidad de colonizar las raíces y reprodución por esporas como mecanismo de supervivencia (Camargo-Ricalde, 2002). En el caso de E. myrtifolia habría una alta colonización radical, con una baja reproducción; en el caso de C. bogotensis, bajos niveles de colonización con la respectiva alta reproducción de la MA.
En el caso de C. bogotensis, la alta variabilidad de los conteos de esporas y el hecho de que esta especie no haya mostrado un efecto significativo del ambiente sobre la colonización, indica que son factores específicos del sitio los que favorecieron la esporulación (Anderson et al., 1983; Camargo-Ricalde y Esperón-Rodriguez, 2005). Por ejemplo, la adición de nitrógeno y fósforo a los suelos reduce la esporulación, mientras altas concentraciones de algunos bioelementos menores la estimulan (Carling et al., 1996; Smith y Read, 2008); esta respuesta puede variar dependiendo de la especie del hongo involucrada en la asociación micorrícica (Douds y Schenck, 1990). La alta densidad de esporas que se dio en C. bogotensis podría deberse al pico de esporulación de alguna especie hospedero-específica (Alexopoulos et al., 1996; Selvaraj et al., 2005). Es decir que en zonas urbanas de Bogotá tanto las caracteristicas especificas de los suelos locales como tambien la población especifica de los HMA nativos, además del posible manejo individual de cada árbol por la adminsitración urbana pueden resultar en una alta variabilidad, marcando dependencia en lugares especificas para algunas especies.
En las zonas secas y semisecas de la ciudad se presentaron los mayores registros de esporulación y colonización radicular. Existe un amplio soporte bibliográfico que indica que los HMA presentan una mayor producción de esporas cuando se requiere soportar los periodos de sequía o de baja precipitación (Guadarrama y Álvarez-Sánchez, 1999; Escudero y Mendoza, 2005; Shahri y Abbasi, 2005). Sin embargo, no existe igual consenso acerca de la colonización; ésta se ve estimulada por aumentos en la humedad del suelo (He et al., 2002; Camargo-Ricalde y Esperon-Rodríguez , 2005; Khanam et al., 2006) y decrece rápidamente al aumentar los días consecutivos de sequía (Braunberger et al., 1994). Otros argumentos indican que realmente los mayores porcentajes de colonización parecen darse en lugares de precipitación moderada, donde luego de la saturación del suelo acontecen unos pocos días de drenaje (Anderson et al., 1986). Este tipo de régimen hídrico es similar al que se da en las zonas secas y semisecas de Bogotá.
La hipótesis planteada, de una relación positiva entre el estado sanitario general y la micorrización, fue comprobada en forma general. Puede ser que gracias a una nutrición más balanceada como resultado de la HMA, las especies vegetales resistan mejor al medio ambiente urbano y así muestren menores sintomatologías asociadas a la deficiencia de nutrientes y la acción de fitopatógenos. Varios autores han reportado que plantas micorrizadas absorben con mayor eficiencia el magnesio (Wang et al., 2008), presentando por tanto mayores contenidos de clorofila, incluso en suelos con altas concentraciones de metales pesados (Paradise et al., 1995; Arriagada et al., 2007) como las que pueden darse en ambientes contaminados. Otros estudios indican que la asociación con HMA aumenta la tolerancia a la herbivoría y reduce la sobrevivencia de ciertos estadios larvales (Vicari et al., 2002; Bennett y Bever, 2007). La relación positiva, marginalmente significativa en E. myrtifolia podría interpretarse en forma conjunta con sus altos porcentajes de colonización radical como una relación de dependencia de la asociación con HMA, mientras que F. soatensis y C. bogotensis tendrían una baja dependencia.
Al evaluar en detalle las relaciones entre el tipo de lesiones foliares y las infecciones radiculares por los HMA, se encontró que la colonización y la esporulación mantienen una relación negativa con la clorosis Bavarescoet al., 2000) en E. myrtifolia y C. bogotensis, y una relación también negativa con la herbivoría y la quemazón en E. myrtifolia resultado que corresponde con el de O'Herlihy et al, (2003) y Siddiqui y Singh (2005). Parece que la micorriza de estas dos especies vegetales reacionan sensiblemente a la disminución del area foliar debido a actividades de patogenos y herbívoros. Una menor área foliar fotosintéticamente activa indica menores carbohidratos para la supervivencia de la micorriza (Smith y Read, 2008). No solo plantas con alto potencial de infeccion micorrízica, como E. myrtifolia, sino tambien plantas con relativamente baja colonización radicular, como C. bogotensis, pueden presentarlo. Las dos especies pueden ser representantes de plantas altamente dependientes y facultativamente dependientes de micorrizas (Janos, 2007). Desde el otro lado de la dependencia en la relación micorrícica, también se debe mencionar que en total, casi el 60% de los afecciones sanitarias de E. myrtifolia y el 17% en C. bogotensis serían controladas por una alta colonización radical.
En suma, los resultados de este estudio permitieron mostrar tres tipos de comportamiento de las especies arbóreas en relación con el establecimiento de asociaciones micorrícicas en el entorno urbano. Eugenia myrtifolia es una especie dependiente de la asociación, con un alto porcentaje de colonización radical, y que exhibe un mejor estado de micorrización con mejores condiciones sanitarias. C. bogotensis es una especie de baja dependencia micorrícica, sin embargo en algunos parques colonización viene acompañada con altos conteos de esporas, por lo tanto se presume que exista asociación específica con alguna especie altamente esporulante o que esta especie depende en ciertas condiciónes edaficas o condiciones de manejo del arbol de las micorrizas y en otras no. Por último, aunque F. soatensis presentó colonización y conteos promedios importantes de esporas en el suelo, no se encontró ningún patrón claro de respuesta, ni en relación con las condiciones ambientales, ni con el estado sanitario general; esto crea inquietud sobre su estado de mícotrofía y la existencia de otros factores que influyen en el grado de asociación de la especie con HMA.
CONCLUSIONES
La respuesta micorrícica en ambientes urbanos difiere de acuerdo a la especie del hospedero y depende del régimen climático - específicamente la precipitación. En general, los ambientes secos y semisecos de Bogotá favorecen tanto la colonización de las especies arbóreas estudiadas como la esporulación en la rizosfera de los HMA. Hay alta diferencia entre las especies arboreas con respecto a la interacción micorriza con el estado fitosanitario. Mientras especies como E. myrtifolia reaccionan sensiblemente en la colonización radical por HMA en relación con el ataques por patogenos y herbiforos foliares, otras plantas, como F. soatensis no muestran ningúna relación. Este puede significar que existen especies arbóreas urbanas que son más robustas en su micorrización durante cambios ambientales o ecologicas que otras. Indudablemente por el hecho de una relación positiva entre el nivel de micorrización y el estado fitosanitario general, se requiren más investigación bajo condiciones controladas para identificar hasta que punto la micorriza controla factores patogénicos y herbívoros en sitios urbanos.
RECOMENDACIONES
Se recomienda ampliamente la implementación de tratamientos en vivero con micorrizas y la adición de inóculos en el transplante definitivo a su lugar de destino, en todas especies arboreas urbanas que forman micorrizas arbusculares. También se recomienda investigar más especies de arbustos o arboles de la zona urbana en invernadero de su dependencia de HMA y la relación con su resistencia a patogenos y herbiveros. Se requiere investigar si las especies arboreas requieren una micorrización especifica de acuerdo a la zona climática de la ciudad.
AGRADECIMIENTOS
Este proyecto fue financiado por el Jardín Botánico José Celestino Mutis, e hizo parte de un convenio interinstitucional con la Fundación Educativa Minuto de Dios. Agradecemos además con especial afecto la colaboración técnica de Nubia Espinoza.
BIBLIOGRAFIA
ALEXOPOULOS CJ, MIMS CW, BLACKWELL M. Introductory Mycology. 4 ed. Nueva York: John Wiley & Sons, Inc.; 1996.
[ Links ]ANDERSON RC, EBBERS BC, LIBERTA AE. Soil Moisture Influences colonization of prairie cordgrass (Spartina Pec Tin Ata Lind.) by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1986; 102:523-527.
[ Links ]ANDERSON RC, LIBERTA AE, DICKMAN LA, KATZ AJ. Spatial Variation in Vesicular-arbuscular mycorrhiza spore density. Torreya 1983; 110:519-525.
[ Links ]ARRIAGADA CA, HERRERA MA, OCAMPO JA. Beneficial effect of saprobe and arbuscular mycorrhizal fungi on growth of Eucalyptus globulus co-cultured with Glycine max in soil contaminated with heavy metals. J. Environ. Manage. 2007; 84: 93-99.
[ Links ]AUDET P, CHAREST C. Effects of AM colonization on wild tobacco plants grown in zinc-contaminated soil. Mycorrhiza 2006; 16:277-283.
[ Links ]AZCÓN-AGUILAR C, BAREA JM. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens - an overview of the mechanisms envolved. Mycorrhiza 1997; 6: 457-464.
[ Links ]BAVARESCO L , CANTU E, TREVISAN M. Chlorosis occurrence, natural arbuscular-mycorrhizal infection and stilbene root concentration of ungrafted grapevine rootstocks growing on calcareous soil. J. Plant Nutr. 2000; 23:1685-1697.
[ Links ]BECKETT P, FREER-SMITH P, TAYLOR G. Effective Tree Species for local air-quality management. J. Arboric. 2000; 26: 12-19.
[ Links ]BENNETT AE, ALERS-GARCIA J, BEVER JD. Three-way interactions among mutualistic mycorrhizal fungi, plants, and plant enemies: Hypotheses and synthesis . Am. Nat. 2006; 167:141-152.
[ Links ]BENNETT A, BEVER, JD. Mycorrhizal species differentially alter plant growth and response to herbivore. Ecology 2007; 88:210-218.
[ Links ]BOROWICZ VA. Do arbuscular mycorrhizal fungi alter plant-pathogen relations? Ecology 2001; 82:3057-3068.
[ Links ]BRAUNBERGER PG, ABBOTT LK, ROBSON AD. The effect of rain in the dry-season on the formation of vesicular-arbuscular micorrizas in the growing season of annual clover-based pastures. New Phytol. 1994; 127:107-114.
[ Links ]CAMARGO-RICALDE SL. Dispersal, distribution and establishment of arbuscular mycorrhizal fungi: a review. Bol. Soc. Bot. México 2002; 71:33-44.
[ Links ]CAMARGO-RICALDE SL, ESPERON-RODRIGUEZ M. Efecto de la heterogeneidad espacial y estacional del suelo sobre la abundancia de esporas de hongos micorrizógenos arbusculares en el valle semiárido de Tehuacán-Cuicatlán, México. Rev. Biol. Trop. 2005; 53:339-352.
[ Links ]CARLING DE, RONCADORI RW, HUSSEY RS. Interactions of arbuscular mycorrhizae, Meloidogyne arenaria, and phosphorous fertilization on peanut. Mycorrhiza 1996; 6:9-13.
[ Links ]CAIRNEY JW. Evolution of mycorrhiza systems. Naturwissenschaften 2000; 87:467-475.
[ Links ]CLARK RB. Arbuscular mycorrhizal adaptation, spore germination, root colonization, and host plant growth and mineral acquisition at low pH. Plant Soil 1997; 192: 15-22.
[ Links ]DAMA (Departamento Técnico Administrativo de Medio Ambiente). Informe Anual de Calidad del Aire: Año 2005. Red de Monitoreo de Calidad del Aire de Bogotá D.C. 2005.
[ Links ]DAMA (Secretaría de Ambiente). Atlas ambiental de Santa Fe de Bogotá. Alcaldía Mayor de Santa Fe de Bogotá. 2007.
[ Links ]DOUDS DD, SCHENCK NC. Increased sporulation of vesicular-arbuscular mycorrhizal fungi by manipulation of nutrient regimens. Appl. Environ. Microbiol. 1990; 56: 413-418.
[ Links ]ESCUDERO V, MENDOZA R. Seasonal variation of arbuscular mycorrhizal fungi in temperate grasslands along a wide hydrologic gradient. Mycorrhiza 2005; 15:291-299.
[ Links ]GEHRING CA, MUELLER RC, WHITHAM TG. Environmental and genetic effects on the formation of ectomycorrhizal and arbuscular mycorrhizal associations in cottonwoods. Oecologia 2006; 149:158-164.
[ Links ]GERDEMANN JW, NICOLSON TH. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963; 46:235-244.
[ Links ]GUADARRAMA P, ÁLVAREZ-SÁNCHEZ FJ. Abundance of arbuscular mycorrhizal fungi spores in different environments in a tropical rain forest, Veracruz, Mexico. Mycorrhiza 1999; 8:267-270.
[ Links ]HE X, MOURATOV S, STEINBERGER Y. Temporal and spatial dynamics of vesicular-arbuscular mycorrhizal fungi under the canopy of Zygophyllum dumosum Boiss. in the Negev Desert. J. Arid Environ. 2002; 25:379-387.
[ Links ]HERREA EA, MEJÍA LC, KYLLO DA, ROJAS E, MAYNARD Z, BUTLER A, VAN BAEL SA. Ecological implications of anti-pathogen effects of tropical fungal endophytes and mycorrhizae. Ecology 2007; 88:550-558.
[ Links ]JANOS DP. Plant responsiveness to mycorrhizas differs from dependence upon mycorrhizas. Mycorrhiza 2007; 17:75-91.
[ Links ]JBJCM (Jardín Botánico José Celestino Mutis). Indicadores Censo del Arbolado: Mes de Septiembre (2008). Disponible en: URL: http://www.jbb.gov.co/jardinbotanico/censo/Avances%20CAU%20WEB%20Septiembre/Memorias%20II%20Encuentro%20Internacional%20de%20Arboricultura%20Urbana/MAPAS%20Censo%20Bogota/
[ Links ]KHANAM D, MIRIDHA MAU, SOLAIMAN ARM, HOSSAIN T. Effect of edaphic factor son root colonization and spore population of arbuscular mycorrhizal fungi. Bull. Ins. Trop. Agr., Kyushu Univ. 2006; 29:97-104.
[ Links ]KHAOSAAD T, GARCIA-GARRIDO JM, STEINKELLNER S, VIERHEILIG H. Take-all disease is systemically reduced in roots of mycorrhizal barley plants. Soil Biol. Biochem. 2007; 39: 727-734.
[ Links ]KORMANIK PP, MCGRAW AC. Quantification of vesicular-arbuscular mycorrhizae in plant roots. En: Schenck NC editor. Methods and Principles of Mycorrhizal Research. St.Paul: The American Phytopathological Society, 1982. p. 37-45.
[ Links ]LUGO MA, GONZÁLEZ-MAZA ME. Arbuscular mycorrhizal fungi in a mountain grassland II: Seasonal variation of colonization studied, along with its relation to grazing and metabolic host type. Mycologia 2003; 95: 407-415
[ Links ]NOWAK DJ. Institutionalizing urban forestry as a biotechnology to improve environmental quality. Urban For. Urban Green. 2006; 5:93-100.
[ Links ]O'HERLIHY EA , DUFFY EM, CASSELLS AC. The effects of arbuscular mycorrhizal fungi and chitosan sprays on yield and late blight resistance in potato crops from microplants. Folia Geobot. 2003; 38:201-207.
[ Links ]PARADISE R, DALPÉ Y, CHAREST C. The combined effect of arbuscular micorrizas and short-term cold exposure on wheat. New Phytol. 1995; 129: 637-642.
[ Links ]PLENCHETT C, CLERMONT-DAUPHIN C, MEYNARD JM, FORTIN JA. Managing arbuscular mycorrhizal fungi in cropping systems. Can. J. Plant Sci. 2005; 85:31-40.
[ Links ]POSADA RH, FRANCO LA, RAMOS C, PLAZAS LS, SUÁREZ JC, ÁLVAREZ F. Effect of physical, chemical and environmental characteristics on arbuscular mycorrhizal fungi in Brachiaria decumbens (Stapf) pastures. J Appl. Microbiol, 2008; 104: 132-140.
[ Links ]RAMOS C. Caracterización Fitosanitaria de Cuatro Áreas Bioclimáticas del Distrito Capital. Informe Técnico JBB463-2007. Bogotá: Jardín Botánico José Celestino Mutis, Subdirección Científica; 2008.
[ Links ]SELVARAJ T, CHELLAPPAN P, YU JIN J, HOON K. Occurrence and quantification of vesicular-arbuscular mycorrhizal (VAM) fungi in industrial polluted soils. J. Microbiol. Biotechnol. 2005; 15: 147-154.
[ Links ]SHAHRI MH, ABBASI M. Variation of spores vesicular-arbuscular mycorrhiza population in Pistachio Natural Forest soil in North of Khorassan. J. Sci. & Technol. Agric. & Natur Resour. 2005; 8: 86-97.
[ Links ]SIDDIQUI ZA , SINGH LP, Effects of fly ash and soil micro-organisms on plant growth, photosynthetic pigments and leaf blight of wheat. J. Plant Dis. Protect. 2005; 112:146-155.
[ Links ]SMITH SE, READ DJ. Mycorrhizal Symbiosis. 3 ed. London: Academic Press; 2008.
[ Links ]TAIZ L, ZEIGER E. Plant Physiology. Sunderland: Sinauer Associates, Inc. 1998.
[ Links ]VICARI M, HATCHER PE, AYRES PG. Combined effect of foliar and mycorrhizal endophytes on an insect herbivore. Ecology 2002; 83: 2452-2464.
[ Links ]VIERHEILIG H, COUGHLAN AP, WYSS U, PICHÉ Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998; 64:5004-5007.
[ Links ]WANG C X, LI XL, ZHOU JC, WANG GQ, DONG YY. Effects of arbuscular mycorrhizal fungi on growth and yield of cucumber plants. Commun. Soil Sci. Plant Anal. 2008. 39:499-509.
[ Links ]WU J. Toward a landscape ecology of cities: beyond buildings, trees and urban forest. En: Carreiro MM, Song Y, Wu J, editors. Ecology, Planning, and Management of Urban Forests: International Perspectives. Nueva York: Springer; 2008. p. 10-28.
[ Links ]ZHAO S, QI X. Signaling in plant disease resistance and symbiosis. J Integr Plant Biol 2008; 50:199-807.
[ Links ]
