










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.16 no.1 Bogotá Jan./Apr. 2011
EFECTO INDUCTOR DEL AGUA DE COCO SOBRE LA GERMINACIÓN DE SEMILLAS Y BROTAMIENTO DE LOS CORMOS DE LA HIERBA DE LA EQUIS Dracontium grayumianum
Inductive Effect of Coconut Water on Germination of Seeds and Sprouting of Corms of Dracontium grayumianum
CARLOS PATIÑO TORRES1, M.Sc., Ph. D.; FERLEY MOSQUERA GAMBOA2, Agrónomo; ROBERT TULIO GONZÁLEZ3, Biólogo 1 Universidad Nacional Abierta y a Distancia - UNAD, Palmira, Colombia. 2 Universidad del Pacífico, Buenaventura, Colombia. Autor para correspondencia: CARLOS PATIÑO TORRES, Docente Auxiliar ECAPMA - Universidad Nacional Abierta y a Distancia - UNAD, Carrera 28 # 40-56 B. Versalles, Palmira, Colombia. cpatinot@yahoo.com
Presentado 28 de febrero de 2010, aceptado 10 de junio de 2010, correcciones el 2 de marzo de 2011.
RESUMEN
El objetivo de este estudio fue determinar el efecto promotor de agua de coco, ácido giberélico, de la estratificación fría y escarificación mecánica, sobre la germinación de semillas de Dracontium grayumianum, y el efecto de ácido giberélico y agua de coco sobre el brotamiento de cormos de la misma especie. Las semillas no sometidas a los tratamientos inductores fueron incapaces de germinar, pero la inmersión en agua de coco tuvo efectos notables, produciendo un porcentaje de germinación del 50%, superior al efecto logrado con los demás tratamientos. El endospermo líquido del coco también tuvo efecto favorable sobre el brotamiento de cormos bajo condiciones de vivero, al igual que el tratamiento con una solución de ácido giberélico. Este es el primer reporte del uso de agua de coco como agente promotor de la germinación de semillas con alto nivel de latencia, lo que coloca este recurso como una alternativa adicional, altamente eficaz y de bajo costo, para ser utilizado en estrategias de propagación vegetal de especies con semillas de latencia profunda.
Palabras clave: Dracontium sp., agua de coco, latencia, dormancia, citocininas.
ABSTRACT
The objective of this study was to determine the promotional effect of coconut water, gibberellic acid, cold stratification and mechanical scarification on seed germination of Dracontium grayumianum, and the effect of gibberellic acid and coconut water on the sprouting of corms of the same species. The seeds without the inductive treatment were unable to germinate, but the immersion in coconut water had significant effects, producing a germination rate of 50%, higher than the effect achieved with other treatments. The liquid endosperm of coconut also had favorable effect on the sprouting of corms under nursery conditions, like the treatment with gibberellic acid solution. This is the first report of the use of coconut water as a promoter of seed germination with high latency, which places this resource as an additional alternative, highly efficient, and cost-effective, for use in plant propagation strategies of species with seeds of deep dormancy.
Key words: Dracontium sp., Coconut water, Dormancy, Latency, Citokinins
INTRODUCCIÓN
El pacífico colombiano es reconocido a nivel mundial por ser parte del -hotspot- de biodiversidad -Tumbes-Chocó-Darién-, albergue de un número de especies vegetales, animales y microbianas incontable, aún no estudiadas por la comunidad científica. La hierba de la equis (Dracontium grayumianum G. Zhu y Croat) , es una de las varias aráceas presentes en la región, aprovechada por diferentes grupos étnicos de la zona con fines farmacéuticos y alimenticios, que actualmente enfrenta graves problemas de erosión genética, debido a los alarmantes problemas de deforestación, y a un aprovechamiento intensivo no sustentable.
Dada la importancia de esta especie para las comunidades negras e indígenas del pacífico colombiano, el enorme potencial que se prevé para su uso a nivel internacional, la falta de estudios científicos sistemáticos y la urgente necesidad de establecer estrategias que eliminen o mitiguen las causas de la pérdida de germoplasma, se hace fundamental adelantar estudios básicos sobre la biología de propagación de la especie, si se quieren establecer métodos eficientes de propagación, los cuales a su vez, sustentarán cualquier programa de conservación genética y también el establecimiento de sistemas productivos sostenibles.
El aislamiento y alta homogeneidad fenotípica de las poblaciones naturales, la no observación in situ de semillas en estado germinativo, y la falta de germinación de semillas de D. grayumianum en ensayos preliminares, permiten suponer un nivel de latencia elevado en aquellas, por lo que el presente estudio se planeó con el objetivo de determinar en vivero, la eficacia de sistemas de propagación sexual y por cormos de la especie, bajo condiciones naturales, así como para evaluar el efecto de varios tratamientos estándares para la promoción de germinación. Igualmente se estudió el efecto del endospermo líquido del coco (agua de coco) como sustancia inductora de la germinación, dado que la consideración de sus cualidades químicas permitió presumir un efecto positivo sobre el proceso.
MATERIALES Y MÉTODOS
LOCALIZACIÓN, MUESTREO Y RECOLECCIÓN DE MATERIAL VEGETAL
Las semillas y cormos se obtuvieron a partir de diferentes poblaciones, pequeñas y aisladas, encontradas en varios sitios ubicados a los márgenes del río Calima, en el corregimiento La Colonia en el Bajo Calima, municipio de Buenaventura. Todas las zonas de recolección presentaron una temperatura promedio de 26,7 ºC, con precipitación media de 7.445 mm/año, humedad relativa de 87% y brillo solar de 2,5 horas luz/día. Los suelos de la zona se han clasificado como entisoles, originados por procesos aluviales y alta meteorización, con pH ácido (4,5 -5,6), alta concentración de aluminio y bajo contenido de bases intercambiables (Melo et al., 1997). La vegetación natural predominante en la zona correspondió a yarumos (Cecropia spp.), chiperos (Zygia longifolia) y caña brava (Gynerium sagittatum). En todos lo sitios muestreados se tomaron plantas individuales fenológicamente similares, a partir de las cuales se obtuvieron semillas sexuales y cormos. Estos se recogieron en bolsas de polietileno negro, utilizando una bolsa por cada planta muestreada. Las bolsas se sellaron y se mantuvieron bajo condiciones ambientales hasta su uso en vivero.
FASE DE VIVERO
Las semillas provenientes de cada una de las plantas se sembraron en bolsas de polietileno de un kilogramo de capacidad. Como sustrato para la siembra se utilizó suelo de formación aluvial recogido de las zonas de crecimiento de las plantas. Se utilizaron 600 g de suelo por bolsa y se sembraron 10 semillas en cada una. Se consideraron germinadas aquellas semillas en las cuales emergió la radícula, para lo cual se hicieron observaciones periódicas a partir de la primera semana de efectuada la siembra, hasta la octava. El porcentaje de germinación de semillas se calculó utilizando la fórmula:
% de germinación=número de semillas germinadas x 100/número de semillas sembradas
Para la determinación del nivel de brotamiento de cormos se sembraron cuatro cormos por bolsa de polietileno con 600 g de suelo, utilizando una siembra superficial. El nivel de brotamiento correspondió al número total de cormos brotados.
Los tratamientos inductores de germinación a los cuales se sometieron las semillas fueron: escarificación mecánica, estratificación con frío, inmersión en solución de ácido giberélico (AG3; 100 mg L-1 por tres días) e inmersión en endospermo líquido de coco (agua de coco) proveniente de frutos jóvenes verdes, por tres días. A los cormos se aplicaron los dos últimos tratamientos. Tanto los tratamientos a semillas como a cormos incluyeron los testigos correspondientes sin tratamiento inductor.
Para la escarificación mecánica, la abrasión de semillas se efectuó manualmente utilizando lija número 120, teniendo cuidado de no dañar el embrión. Una vez retirada la cubierta, las semillas se lavaron abundantemente con agua lluvia, luego de lo cual se sembraron para la determinación posterior del porcentaje de germinación.
En el tratamiento de estratificación con frío, las semillas se dejaron inmersas en agua lluvia por tres días, al cabo de los cuales se almacenaron en nevera en bolsas de papel, cubiertas a su vez con una bolsa plástica, a una temperatura de 3 ºC por 20 días. Después de este tiempo las semillas se sembraron como se describió.
DISEÑO Y ANÁLISIS ESTADÍSTICO
Los ensayos se hicieron utilizando un diseño en bloques al azar, correspondiendo el bloque, al genotipo de la planta donadora de semillas y cormos. Se usaron tres repeticiones por tratamiento. El análisis de varianza se efectuó utilizando SPSS v. 14 y la comparación de medias se realizó con la prueba de Tukey, a un α = 0,05.
RESULTADOS
PORCENTAJE DE GERMINACIÓN DE LAS SEMILLAS DE D. GRAYUMIANUM
Los resultados obtenidos en los experimentos de germinación de semillas de D. grayumianum, tanto bajo condiciones naturales como de germinación inducida, se presentan en la Figura 1. Cuando las semillas no recibieron tratamiento alguno, estas fueron incapaces de germinar, incluso después de tres meses de efectuada la siembra, lo que implica la existencia en las semillas de esta especie, de mecanismos endógenos de latencia bastante difíciles de superar bajo condiciones naturales.
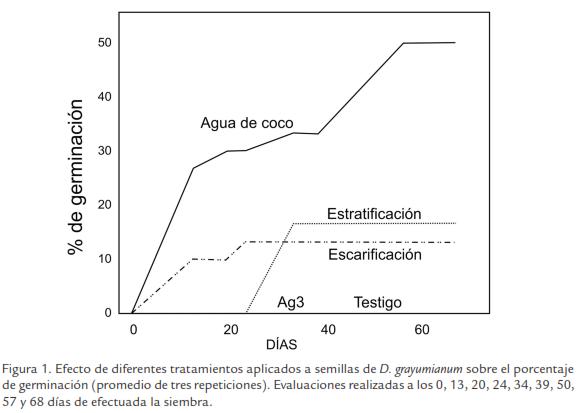
De los tratamientos aplicados para promover germinación, la inmersión en agua de coco fue la alternativa con mejores resultados, alcanzándose niveles máximos de germinación de 50,0%, a partir del día 57 después de la siembra. El efecto de agua de coco fue evidente a partir del día 13, momento en el que se presentó un porcentaje de germinación del 26,7%. La escarificación mecánica presentó también efectos positivos, menores sin embargo, a los obtenidos con agua de coco. En este caso, 10% de las semillas germinaron al día 13 después de la siembra, alcanzando un nivel máximo de germinación a partir del día 24 con un 13,3% de germinación. La estratificación fría produjo niveles de germinación de 16,7% a partir del día 34, manteniéndose en éste valor hasta el día 68 de la evaluación. Contrario a lo que se esperaba, con base en lo reportado para semillas de muchas especies, el ácido giberélico no tuvo ningún efecto sobre el porcentaje de germinación, durante todo el período evaluado.
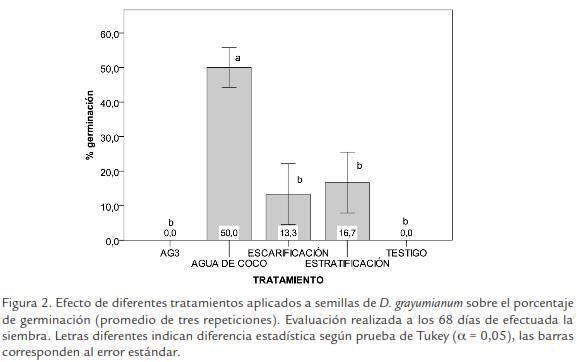
El análisis de varianza para los resultados obtenidos a los 68 días, demostró diferencias altamente significativas entre las respuestas de germinación de las semillas a los tratamientos aplicados. De acuerdo con la prueba Tukey, el tratamiento consistente en inmersión en agua de coco produjo efectos que presentan diferencia estadística con todos los demás tratamientos. Los tratamientos de estratificación fría, escarificación mecánica, inmersión en ácido giberélico no presentaron diferencias estadísticas significativas entre sí, ni con respecto al testigo (Fig. 2).
NIVEL DE BROTAMIENTO DE CORMOS DE D. GRAYUMIANUM
La Figura 3 resume los resultados obtenidos para el brotamiento de cormos de D. grayumianum, sometidos a diferentes tratamientos promotores de brotamiento o bajo condiciones naturales, después de 34 días de la siembra.
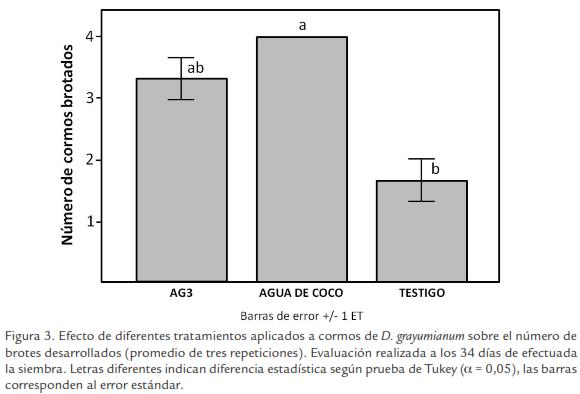
Tanto los tratamientos inductores (inmersión en agua de coco e inmersión en ácido giberélico), como el testigo, produjeron brotamiento, en diferentes niveles. De nuevo, el control presenta los más bajos índices de brotamiento. Para el día 13 después de efectuada la siembra, el tratamiento control había alcanzado 25% de brotamiento, mientras los tratamientos de inmersión en agua de coco y de inmersión con ácido giberélico habían producido niveles de brotamiento de 83,25% y de 75%, respectivamente. Estadísticamente, a los 34 días de la siembra, el andeva no detectó diferencias significativas entre los efectos de los tres tratamientos, aunque si lo hizo la prueba de Tukey (Fig. 3).
Después de 34 días de efectuada la siembra los niveles de brotamiento alcanzados habían llegado a 100% para el caso del tratamiento con el agua de coco y a 83,25% en el caso del tratamiento con ácido giberélico, el tratamiento control sólo logró 41,75% de brotamiento. Estos niveles de brotamiento se mantuvieron constantes hasta el día 89 después de la siembra de cormos.
DISCUSIÓN
Existen varios estudios que permiten suponer que los efectos obtenidos en la tasa de germinación de las semillas de D. grayumianum sometidas al tratamiento de inmersión en agua de coco, y la mayor proporción de brotamiento de cormos, son debidas, sino total-mente, al menos en parte, a la composición nutritiva y hormonal rica del endospermo líquido de coco (Ge et al., 2004; Ge et al., 2005; Richter et al., 2005; Yong et al., 2009; Stirk et al., 2005). El agua de coco contiene diversidad de hormonas con acción citoquinina, por ejemplo, del tipo isoprenoide, las cuales están implicadas en el proceso de división celular, y aromáticas, implicadas en procesos posgerminativos (Del Pozo et al., 2005).
Dado que las semillas de D. grayumianum pueden contener niveles muy bajos de citocininas (en el rango de pM), la adición de agua de coco podría bastar para suplir a la semilla con niveles y tipos adecuados de estas hormonas, que permitan romper la latencia observada en semillas bajo condiciones naturales. Generalmente, las aráceas presentan dormancia de tipo fisiológico, es decir, que el embrión no ha alcanzado el estado de madurez una vez las semillas han caído al suelo, lo que implica que el eje embrionario no ha alcanzado un estado de desarrollo apto para producir suficiente cantidad de citocininas. De hecho, Pickett, 1913, y Yang et al., 1999, demostraron experimental-mente que la germinación en Arisaema dracontium y Arisaema triphyllum (dos aráceas relacionadas con D. grayumianum), toma entre dos a cinco meses, e indicaron que en el primer año de crecimiento el embrión daba origen a un cormo, el cual producía raíces pequeñas, los nutrientes eran transferidos de la semilla al cormo, todo antes de la formación de cualquier hoja primaria u otros órganos fotosintéticos. Según Yang et al., 1999, Rennert publicó un artículo en 1902, en el que comparó la morfología y anatomía de plántulas de A. dracontium y A. Triphyllum, sugiriendo que la lenta germinación del primero podría estar relacionada a inmadurez anatómica y fisiológica del embrión, aunque el embrión parecía ser morfológicamente maduro.
Yang et al., 1999, encontraron que la germinabilidad de semillas en A. dracontium varió entre 25 y 55% dependiendo de la fuente de las semillas, provenientes de diferentes lugares. En el presente estudio, incluso después de seis meses de haber sembrado las semillas de D. grayumianum, estas no germinaron bajo las condiciones descritas. Aparte de su efecto en la movilización de nutrientes del tejido endospermático al embrión, el efecto de citocininas contenidas en agua de coco, especialmente las de tipo isoprenoide, podría darse con mayor intensidad a través de su implicación en la regulación del ciclo celular, por su efecto sobre síntesis de DNA y a través de su efecto sobre elongación de la célula, procesos clave durante la germinación. Igualmente, estas hormonas han sido implicadas concluyentemente en procesos de diferenciación, condición inherente al desarrollo de una plántula a partir del embrión, lo que apoya aún más, su efecto promotor en la germinación.
En relación con el incremento en el porcentaje de brotamiento de cormos de D. grayumianum, el efecto principal del agua de coco podría deberse a su contenido de citocininas aromáticas, las cuales promueven formación de clorofila y generación de raíces, estimulando crecimiento y desarrollo de plántulas. De hecho, las plántulas que se desarrollaron de cormos tratados con agua de coco, presentaron un follaje más denso y hojas de mayor tamaño, comparadas con los demás tratamientos.
En este estudio, también se observaron efectos positivos en la germinación para los tratamientos de estratificación fría y mecánica. La estratificación fría ha sido señalada como factor clave en el rompimiento de dormancia de varias especies de aráceas y consiste, en someter semillas a bajas temperaturas previa imbibición de las mismas. El efecto de la estratificación fría parece ser la promoción del desarrollo del embrión cuando éste presenta algún tipo de inmadurez morfológica y/o fisiológica.
Hay numerosos estudios que demuestran que la estratificación con frío es una medida efectiva para romper dormancia en semillas (Flórez y Pedroza, 2006). En el caso de A. dracontium, las semillas de varias poblaciones evaluadas, excepto una, mostraron incremento en la germinabilidad relativa, de 25 a 40% después de ser expuestas a baja temperatura (3 ºC) por tres semanas antes de ser incubadas (Yang et al., 1999). En el presente estudio, las semillas se dejaron en agua destilada por tres días, después de lo cual se mantuvieron a 3 ºC por 20 días, considerando lo reportado para otras aráceas. Con éste tratamiento se obtuvo una porcentaje de germinación de 13,3%, lo que corrobora algún mecanismo de dormancia secundaria en semillas de D. grayumianum.
De acuerdo con Yang et al., 1999, los efectos de la estratificación pueden estar relacionados con reducción en los niveles de ABA e incremento en el contenido de ácido giberélico, según se ha demostrado en semillas estratificadas de muchas especies. La promoción de crecimiento del embrión también ha sido reportada en semillas de cereza. La estratificación implica cambios de temperatura, y ésta es vista como el factor principal que regula dormancia y germinación de semillas en especies de climas templados (Baskin y Baskin, 1988).
Aunque las poblaciones de D. grayumianum solo están presentes en hábitats no sujetos a estaciones, también es cierto, que crecen en los suelos de las márgenes de ríos, sitios propensos a ciclos casi diarios de inundación, lo que podría causar fluctuaciones de temperatura que podrían a su vez, regular algún tipo de dormancia relacionada con factores fisiológicos o morfológicos del embrión.
De otra parte, en cuanto se relaciona con el efecto de escarificación mecánica, tratamiento que también favoreció la germinación de semillas de D. grayumianum, es la dureza de la cubierta seminal la que condiciona si se ha de producir la emergencia o nó de la radícula. En este sentido, disminuir el espesor de la cubierta, de gran resistencia en el caso de las semillas de la hierba de la equis, favorece sin duda la disminución de la resistencia que ofrece la cubierta seminal al embrión en crecimiento. El comportamiento de las semillas obtenido en este ensayo podría indicar que la dormancia de las semillas de D. grayumianum también se debe en parte, a la dureza de su cubierta, es decir, también presentan un tipo de dormancia debida a factores mecánicos.
Es sorpresivo el hecho de que el tratamiento con ácido giberélico no tuviese ningún efecto sobre la germinación de las semillas de D. grayumianum. Existe considerable evidencia de que giberelinas (AG) y ABA endógenos están involucrados en mecanismos de germinación de semillas (Nonogaki et al., 2010; Schoonheim et al., 2009). Varios procesos bioquímicos en las semillas en germinación parecen estar mediados directamente por giberelinas, tales como movilización de reservas alimenticias y debilitamiento de la pared celular.
Es probable que la falta de efecto del ácido giberélico sobre la germinación de las semillas de D. grayumianum en los presentes ensayos se deba a que ésta sustancia se aplicó sola a las semillas, por lo que no pudo interactuar con el tipo y/o cantidades adecuadas de otros fitoreguladores, especialmente citocininas. Muchos estudios han demostrado que la acción de giberelinas en germinación puede potencializarse cuando se usan junto con citocininas. De otra parte, el hecho de que el ácido giberélico no fuese suficiente para romper la latencia de las semillas, parece indicar que en el caso de D. grayumianum, la luz sólo juega un papel secundario en su proceso de germinación, pues es bien sabido que GA reemplazan en su efecto a la luz (Thomas et al., 1997). Igualmente, la dureza de la cubierta de la semilla de D. grayumianum puede constituirse en una barrera que impide la traslocación de giberelinas a los tejidos internos del endospermo, en donde estas hormonas ejercen su mayor actividad.
En el caso del brotamiento de cormos, AG3 tuvo efectos positivos, al permitir mayor porcentaje de brotamiento, pero también al producir un crecimiento más vigoroso, en términos de densidad de follaje y tamaño de hojas, aunque menor que el producido por agua de coco, en relación con el tratamiento control. En este caso, el efecto principal de giberelinas podría suponerse debido a la promoción de una mayor tasa de elongación celular y al fortalecimiento del crecimiento de las plántulas.
Los resultados obtenidos en este estudio con semillas de D. grayumianum muestran claramente que las mismas tienen mecanismos endógenos responsables del alto nivel de latencia observado en ellas. Los efectos del tratamiento de la escarificación mecánica muestran que aunque la latencia mecánica tiene lugar, debido a la dureza de la cubierta de las semillas, esta no es exclusiva y tan solo juega un papel de menor importancia relativa, pues el tratamiento de inmersión en agua de coco, aplicado sólo, fue suficiente para incrementar la tasa de germinación. En esta vía, también puede deducirse que las semillas de D. grayumianum pueden presentar algún tipo de dormancia fisiológica, lo que generaría pobre producción de citocininas y por tanto un requerimiento de citocininas exógenas para poder germinar. Igualmente, los resultados con la estratificación fría apoyan una dormancia de tipo fisiológica o morfológica. Los resultados vistos integralmente, también hacen suponer algún tipo de termodormancia.
Desde una perspectiva ecológica, la dormancia secundaria encontrada en las semillas de D. grayumianum, puede tener consecuencias importantes en las estrategias reproductivas y ciclos de vida de sus poblaciones. De forma natural, las poblaciones de la hierba de la equis se encuentran aisladas y con muy pocos miembros, pero compartiendo nichos ecológicos muy similares. Esta estructura poblacional puede ser consecuencia de una baja eficiencia reproductiva de la especie debida a viabilidad baja de las semillas, a un alto nivel de latencia de las mismas o a una combinación de las dos. De ser ciertas, estas condiciones favorecerían la reproducción por cormos, los cuales en los ensayos demuestran ser mucho más eficientes reproductivamente. Genéticamente, esta condición podría demostrarse por un bajo nivel de heterocigosis en las poblaciones y por tanto con muy poca variabilidad genotípica dentro de poblaciones aisladas; sin embargo, podría esperarse que poblaciones aisladas presenten algún grado de divergencia genotípica, constituyéndose en ecotipos adaptados a condiciones particulares del hábitat en que se hallan.
Los resultados apuntan a concluir que las poblaciones de D. grayumianum son altamente susceptibles a la erosión genética, debido a la falta de plasticidad y de respuesta a condiciones ambientales cambiantes. La comprobación de si este es o no el caso, depende de estudios adicionales tendientes a caracterizar la especie tanto a nivel morfológico como molecular que demuestren su constitución genética con fines comparativos.
AGRADECIMIENTOS
Los autores manifiestan su agradecimiento a la comunidad del Bajo Calima, por facilitar los trabajos de recolección de semillas y cormos en su región.
BIBLIOGRAFÍA
BASKIN CC, BASKIN JM. Germination ecophysiology for herbaceous plant species in a temperate región. Am J Bot. 1988;75:286-305.
[ Links ]DEL POZO JC, LOPEZ-MATAS MA, RAMIREZ-PARRA E, GUTIERREZ C. Hormonal control of the plant cell cycle. Plant Physiol. 2005;123:173-183.
[ Links ]FLÓREZ V, PEDROZA J. Germinación y dormancia. Universidad Nacional de Colombia, Bogotá; 2006.
[ Links ]GE L, HONG YONG J, GOH NK, CHIA LS, TAN SN, ONG ES. Analysis of some cytokinins in coconut (Cocos nucifera L.) water by micellar electrokinetic capillary chromatography after solid-phase extraction. J Chromatogr A. 2004;1048(1):119-126.
[ Links ]GE L, HONG YONG J, GOH NK, CHIA LS, TAN SN, ONG ES. Identification of kinetin and kinetin riboside in coconut (Cocos nucifera L.) water using a combined approach of liquid chromatography-tandem mass spectrometry, high performance liquid chromatography and capillary electrophoresis. J Chromatogr B. 2005;829(1-2):26-34.
[ Links ]MELO CO, MARTÍNEZ H, HUERTAS FA. Curso taller sobre Evaluación de la Diversidad Florística y Análisis Estructural de Ecosistemas Boscosos Naturales. Ministerio del Medio Ambiente. Universidad del Tolima. Buenaventura, 1997. p. 130.
[ Links ]NONOGAKI H, BASSEL GW, DEREK BEWLEY J. Germination-Still a mystery. Plant Sci. 2010;179(6):574-581.
[ Links ]PICKETT FL. The germination of seeds of Arisaema. Proc Indian Natl Sci Acad B Biol Sci. 1913;23:125-128.
[ Links ]RICHTER EM, de JESUS DP, MUÑOZ RA, do LAGO CL, ANGNES L. Determination of anions, cations, and sugars in coconut water by capillary electrophoresis. J Braz Chem Soc. 2005;16(6a):1134-1139.
[ Links ]SCHOONHEIM PJ, DA COSTA PEREIRA D, DE BOER AH. Dual role for 14-3-3 proteins and ABF transcription factors in gibberellic acid and abscisic acid signalling in barley (Hordeum vulgare) aleurone cells. Plant Cell Environ. 2009;32:439-447.
[ Links ]STIRK WA, GOLD JD, NOVÁK O, STRNAD M, VAN STADEN J. Changes in endogenous cytokinins during germination and seedling establishment of Tagetes minuta L. Plant Growth Regul. 2005;47(1):1-7.
[ Links ]THOMAS TH, HARE PD, STADEN JV. Phytochrome and cytokinin responses. Plant Growth Regul. 1997;23:105-122.
[ Links ]YANG J, LOVETT-DOUST J, LOVETT-DOUST L. Seed germination patterns in green dragon (Arisaema dracontium, araceae). Am J Bot. 1999;86(8):1160-1167.
[ Links ]YONG JW, GE L, NG YF, TAN SN. The chemical composition and biological properties of coconut (Cocos nucifera L.) water. Molecules. 2009;14:5144-5164.
[ Links ]
