










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.17 n.1 Bogotá jan./abr. 2012
EFICIENCIA EN EL RETORNO POTENCIAL DE NUTRIENTES VÍA HOJARASCA DE UN BOSQUE TROPICAL DE RIBERA. SIERRA NEVADA DE SANTA MARTA COLOMBIA
Efficient Way Back Litter Nutrient Potential of a Tropical Forest Ofbank. Sierra Nevada of Santa Marta Colombia
NATALIA FUENTES MOLINA1, M.Sc.; JAVIER ALFREDO RODRÍGUEZ BARRIOS2
1 Grupo Madre Tierra, Universidad de la Guajira, Km 5, vía a Maicao Riohacha Guajira, PBX: 728 27 29; nataliafuentesmolina@yahoo.com
2 Doctor en Ciencias-Biología, Grupo de Biodiversidad y Ecología Aplicada, Instituto de Investigaciones Tropicales, Universidad del Magdalena. Carrera 32 # 22 08, Santa Marta, Colombia. PBX: 421 79 40; jarodriguezba@gmail.com
Presentado 12 de mayo de 2011, aceptado 11 de octubre de 2011, correcciones 13 de octubre de 2011.
RESUMEN
En tres formaciones vegetales representativos de la ribera del río Gaira, (bosque muy húmedo subtropical (bmh-ST), bosque húmedo subtropical (bh-ST) y monte espinoso tropical (me-T)), fueron medidos durante un período de seis meses (lluvioso y seco) los flujos de los nutrientes nitrógeno (N) y fósforo (P) a través de la hojarasca. Las concentraciones de estos nutrientes fueron semejantes en las tres formaciones vegetales (1,71% N y 0,12% P para el bh-ST; seguidos por el me-T con 1,50% N y 0,10% P y el bmh-ST con 1,39% N y 0,08% P), presentándose las diferencias más notorias para N, siendo el bh-ST establecidos en el tramo medio de la cuenca el que presentó las concentraciones más elevadas. Los mayores retornos de biomasa y nutrientes se presentaron en el bh-ST y el me-T establecidos en el tramo medio y bajo de la cuenca. La hojarasca mostró alta concentración de N y consecuentemente, dados los altos valores de producción de las diferentes fracciones, un retorno potencial alto de N (78,6 kg ha-1 año-1). P presentó una concentración foliar con un retorno potencial de 4,9 kgha1año-1y altos valores en los índices de eficiencia de uso (IEV: 2888,5) y la reabsorción foliar (ERN: 98,2), fue el nutriente más limitante.
Palabras clave: nutrientes, caída de hojarasca, Cuenca.
ABSTRACT
In three representative forests along the river Gaira, (subtropical wet forest, subtropical moist forest and tropical thorn mount), were measured over six months (wet and dry seasons) fluxes of nitrogen and phosphorus through the litter. Concentrations of nutrients (nitrogen and phosphorus) in the litter were relatively similar in the three Nevada de Santa Marta Colombia. Fuentes, Rodríguez. vegetation types (1.71% N and 0.12% P for the subtropical moist forest, followed by the tropical thorn mount with 1.50% N and 0.10% P and the subtropical wet forest with 1.39% N and 0.08% P), with the most significant differences found for nitrogen, which is the major nutrient with the absolute maximum in the subtropical rain forest set in the middle stretch of the basin. The greatest returns on biomass and nutrients occurred in the subtropical moist forest and tropical thorn mount set in the middle and lower reaches of the basin. The leaves showed high concentration of N and consequently, given the high production values of the different fractions, a high potential return of N (78.6 kg ha-1 yr-1). The foliar P concentration showed a potential return of 4.9 kgha1año-1 and high values of the indices of efficiency in their use (IEV:2888.5) and foliar resorption (ERN: 98.2), was the nutrient most limiting.
Key words: Nutrients, litterfall, riparian forest, watershed.
INTRODUCCIÓN
El efecto de la biomasa aérea y la hojarasca sobre la productividad de los ecosistemas lóticos ha sido estudiada por varios autores (Simony Benfiel, 2001; Paul y Hall, 2002); encontrándose que la hojarasca y en especial las hojas caídas son las que generan el mayor ingreso de nutrientes a este tipo de ecosistemas (Vitouseket al., 1995; Landsberg y Gower, 1997), contribuyendo en su conservación. Sin embargo, las actividades antrópicas generadas sobre ecosistemas forestales, repercuten sobre los ecosistemas lóticos una vez que se altera la vegetación de las márgenes, denominada riparia o ribereña, que se encuentra en íntima relación con el ambiente acuático.
Los nutrientes confinados en la hojarasca foliar constituyen fuente importante de recursos, por lo que la cuantificación de su producción y su naturaleza, son cruciales para comprender el reciclaje de nutrientes en ecosistemas lóticos y forestales (Ramírez et al., 2007); además de la caída de hojarasca, existen procesos de fuertes implicaciones en la dinámica de los nutrientes, tal es el caso de la reabsorción de nutrientes foliares, que conduce a mayor eficiencia de su uso, la cual puede ser el resultado de mayor fracción de nutrientes reabsorbida desde las partes senescentes de la planta previa a la abscisión, o de mayor cantidad de materia orgánica o carbono fijado por unidad de nutriente utilizado (Hirose, 1975; Vitousek, 1984).
Aunque muchos aspectos de la estructura y función de ecosistemas boscosos fluviales han sido investigados en décadas anteriores (Cushing et al.,1995; Malanson, 1995), la importancia de estos en la dinámica de los aportes de nutrientes a las comunidades acuáticas aún es poco conocido (Albariño, 1999; Ríos y Bailey, 2006). Sin embargo, el reemplazo de bosques nativos o la repoblación forestal de cuencas hidrográficas con especies exóticas, es una práctica cada vez más común (Albariño, 1999). Lo anterior es realmente cierto cuando las políticas ambientales se realizan a partir de elementos estructurales, dejando de lado la funcionalidad de las especies vegetales como un proceso que puede reflejar de una forma más evidente este tipo de alteraciones (Benfield, 1996; Flory y Milner, 1999).
El objeto del presente estudio consistió en evaluar la dinámica de los nutrientes a través de la caída de hojarasca y la eficiencia en su uso, realizada en tramos de bosque de ribera, representados por tres formaciones vegetales, ubicados a lo largo de un gradiente altitudinal del río Gaira. Estos procesos permiten valorar la capacidad que tienen dichas formaciones vegetales para aportar materia orgánica y nutriente al cuerpo hídrico y observar su desempeño desde la óptica del uso funcional de los nutrientes limitantes, para el crecimiento y desarrollo de las comunidades acuáticas asociadas. La caracterización de estos aspectos genera información de importancia para establecer la dinámica espacio-temporal de los nutrientes. Para esto, se valoraron i. Las tasas de los nutrientes (N y P) aportados por las diferentes fracciones que componen la hojarasca, ii. El retorno potencial de nutrientes vía hojarasca foliar, iii. Las tasas de reabsorción de nutrientes foliares móviles y iv. La eficiencia en su uso.
MATERIALES Y MÉTODOS
DESCRIPCIÓN DEL ÁREA DE ESTUDIO
El estudio se realizó entre los meses de septiembre y febrero de 2009, en la cuenca hidrográfica del río Gaira, entre los 11°52'06"-11°10'08"N y 74°46'22"-74°11'07"W, la cual se encuentra ubicada en la vertiente noroccidental de la sierra nevada de Santa Marta, en el departamento de Magdalena, Colombia (Tabla 1); en un área de 10.464,3 ha, la dirección del flujo cursa de este a oeste, con una longitud de 32,53 km aproximadamente desde su nacimiento hasta su desembocadura en el mar Caribe (Frayter et al., 2000).

En el presente estudio se seleccionaron tres tramos de 200 m (aproximadamente), ubicados: (1) el sector de San Lorenzo (tramo alto); (2) el sector la victoria (tramo medio); y (3) el sector de Puerto Mosquito (tramo bajo) corresponde a la zona donde se presentan mayores intervenciones antrópicas producto de actividades agrícolas y turísticas, reemplazando la vegetación nativa de estos ecosistemas por otras especies exóticas (Tabla 2).

MÉTODOS DE CAMPO Y LABORATORIO
La producción de hojarasca fue evaluada como la biomasa acumulada mensualmente (Búrquezet al., 1999; Enrigth, 1999) abarcando períodos de lluvia y sequía (Rai y Proctor, 1986) durante seis meses; para lo cual se establecieron tramos de estudio permanentes; para la recolección de hojarasca se ubicaron seis trampas de forma aleatoria (unidades muestréales de 1 m2 por cada trampa) por tramo (Proctor, 1983), número considerado aceptable (Stocker et al., 1995) que permite un error estándar de la media menor a cinco por ciento (Anderson e Ingram, 1993). Las trampas fueron elaboradas en tela de malla fina (ojo de maya de 0,25 mm), que permitieron recuperar los órganos vegetales de menor tamaño y la salida del agua de lluvia y un marco fijo de tubo de ½ pulgada que logró impedir el movimiento por acción del viento (Modificado de Rodríguez, 2006).
Para el seguimiento del proceso de reabsorción de nutrientes foliares móviles, se tomaron mensualmente muestras de hojas maduras, en buen estado fotosintético y libres de plagas o enfermedades (León et al., 2009). Las hojas verdes y la hojarasca se secaron en una estufa a 60 °C (Aussenac et al., 1972), hasta peso constante en una balanza analítica. Posteriormente, para conocer el aporte en biomasa de los distintos componentes de la hojarasca, el material fue discriminado, en las siguientes fracciones: hojas, madera y ramas con diámetro <5cm (Hernández, 2000; Caldentey et al., 2001), estructuras reproductivas y material misceláneo. Como misceláneo se consideró a cualquier material que no se puedo identificar como una de las fracciones antes mencionadas (Cuevas y Lugo, 1998).
Posteriormente, cada fracción fue homogenizada, molida y tamizada con molino Wiley y malla 20 para la mineralización en húmedo (Luh Huang y Schulte, 1985), dejando los elementos en condiciones adecuadas para las determinaciones del contenido total de N y P presente en cada fracción. El P fue cuantificado colorimétricamente siguiendo el método del ácido ascórbico (Klute, 1986); y el N fue reducido a compuestos de amonio por medio de digestión Kjeldahl (Tabatabai y Bremner, 1991).
Como medida para evaluar la eficiencia en el uso de los nutrientes se empleó el índice de eficiencia de Vitousek (Vitousek, 1984),
IEV=PSh/PNh; (1)
donde PSh es el peso seco de la hojarasca foliar (g·m-2·mes-1), y PNh es el peso del nutriente en la hojarasca foliar (g·m-2·mes-1). La reabsorción se evaluó empleando el método propuesto por Finzi et al., 2001, que está fundamentada en la relación existente entre los nutrientes presentes en las hojas verdes y en la hojarasca foliar (Del Arco et al., 1991).
ERN= (Chv-Ch/Chv) x100 (2)
donde Chv es la concentración de nutrientes en las hojas verdes (%), y Ch es la concentración de nutrientes en la hojarasca (%).
ANÁLISIS DE DATOS
Para el análisis estadístico de las concentraciones de nutrientes (N y P) presentes en la hojarasca, se realizaron las pruebas estadísticas, mostrando un comportamiento aleatorio de las variables, pero los supuestos de normalidad y homogeneidad resultaron no ser cumplidos en la gran mayoría de los casos, por lo que todos los análisis estadísticos que se presentan, fueron realizados con base en pruebas no paramétricas, en el paquete estadístico R Versión 2.11.1. Los análisis de las concentraciones de nutrientes en las diferentes fracciones de la hojarasca analizada comprendieron los siguientes procedimientos: i. análisis de escalamiento multidimensional no métrico (nMDS) en los tres tramos y durante el período seco-lluvioso; ii. análisis de varianza multivariado no paramétrico a dos vías (PERMANOVA), se utilizó para evaluar el nivel de significancia de los patrones encontrados en el análisis exploratorio de los nMDS, para las concentraciones de N y P (PERMANOVA 1) y las diferentes fracciones de hojarasca (PERMANOVA 2), iii. análisis de las variaciones encontradas en las concentraciones de N y P aportados por las diferentes fracciones en las formaciones vegetales se realizó aplicando análisis de varianza no paramétrico a una vía Kruskal-Wallis (K-W), iv. Se realizó la prueba W de Mann Whitney para evaluar las diferencias entre las concentraciones de nutrientes encontradas en los períodos de lluvia y sequía.
RESULTADOS PATRONES TEMPORALES
Los datos arrojados por las estaciones pluviométricas ubicados en cercanías al área de estudio, indican que los valores de precipitación para los primeros muestreos (M1, M2 y M3) corresponden a un período de alta precipitación (P2), presentándose los valores más altos (80,2 mm.d-1) y en los muestreos finales (M4, M5 y M6) los valores de precipitación no superaron los 20 mm.d-1 lo cual corresponde a un período de precipitación baja (FP1; Fig. 1A, 1B y 1C).
Presentándose así diferencias significativamente entre los períodos seco y lluvioso (U=35 p=0,0004, gl=16), aunque en términos generales se observa un régimen de precipitación variable con valores relativamente bajos. Los umbrales de precipitación evaluados, generaron efectos en los aportes de la biomasa provenientes de la hojarasca estacional; en los registros se presentan valores de precipitación altos en el T2 en relación con el T3 y estos a su vez superiores a los reportados en el T1.
La producción anual de hojarasca en el área de estudio fue de 2745,71 Kgha-1a-1 (Tabla 3), con los menores valores en el P2; los mayores valores de producción, estuvieron representados por las formaciones vegetales establecidas en el T2 y T3; se evidencia que la producción anual de hojarasca no presentó diferencias significativas entre los tramos evaluados, mostrando solo diferencias entre los períodos evaluados (K-W= 17,06, p=0,0043, n=98), con los picos de mayor producción en el P1 (Figura 1D).

La caída de hojarasca estuvo dominada por la fracción foliar (5561,85 Kgha-1a-1) y el material leñoso (3061,98 Kgha-1a-1); los menores valores fueron registrados para la fracción reproductiva y miscelánea (1100,23 Kgha-1a-1 y 1258,75 Kgha-1a-1 respectivamente).
CONCENTRACIÓN DE NUTRIENTES EN LA HOJARASCA
Los mayores contenidos de N y P en la hojarasca del bosque de la ribera del río Gaira, fueron registrados por las formaciones vegetales establecidas en T2 (1,63%N y 0,10%P) y T3 (1,52% N y 0,13% P; Figura 2A y 2C) y los menores valores en T1 (N: 1,39% y P: 0,08 %); presentando diferencias significativas entre las formaciones vegetales (K-W=11,34, p=0,003, n=98) sólo en el contenido de N. Los aportes de NH presentan sus mayores asociaciones con T1 y T2, NF presenta sus asociaciones con T2 al igual que NM y finalmente T3 presenta sus mayores relaciones con el nitrógeno del material leñoso (NL; Figura 2B y 2D).
La tendencia evidenciada en este estudio, es que las concentraciones de N tienden a ser mayores en las hojas (1,56%) que en las ramas (1,38%) y, a su vez, mayores en estos que en los frutos (1,16%); lo anterior difiere de las concentraciones de P donde los valores más altos corresponden a flores (0,14%) seguidos por hojas (0,09%) y finalmente material leñoso (0,05%).
Las fracciones de N evaluadas en las diferentes formaciones vegetales (Figura 2A, fueron estadísticamente diferentes (factor tramo: seudo F=5,41; g.l=2; p=0,005), con un claro incremento exponencial positivo de la fracción NM en relación al aumento de la biomasa del material aportado (Figura 2E). Se presenta una clara tendencia exponencial positiva en T1, donde los incrementos de biomasa corresponden con los incrementos del PH y PL (Figura 2F).
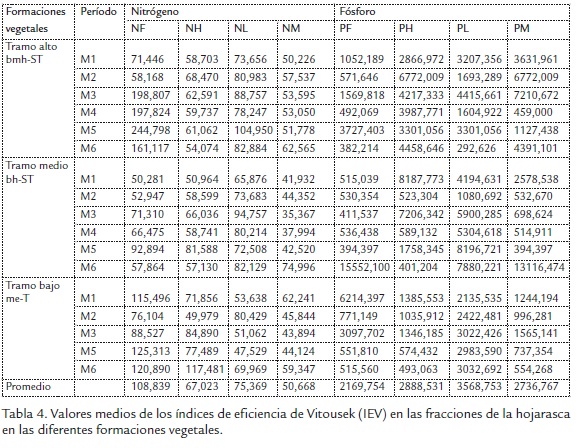
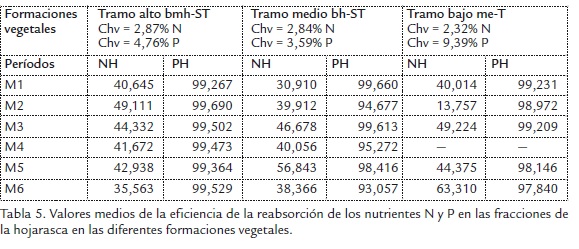
Las concentraciones de NH y NM presentaron alta asociación con el período seco (P1), el NF con el períodolluvioso (P2) y el l NL presentó asociaciones con los dos períodos de precipitación (Figura 3A). Las tendencias temporales de P se caracterizaron por presentar los máximos aportes en el PH y PM con asociaciones en los períodos P2 y P1 (Figura 3B).
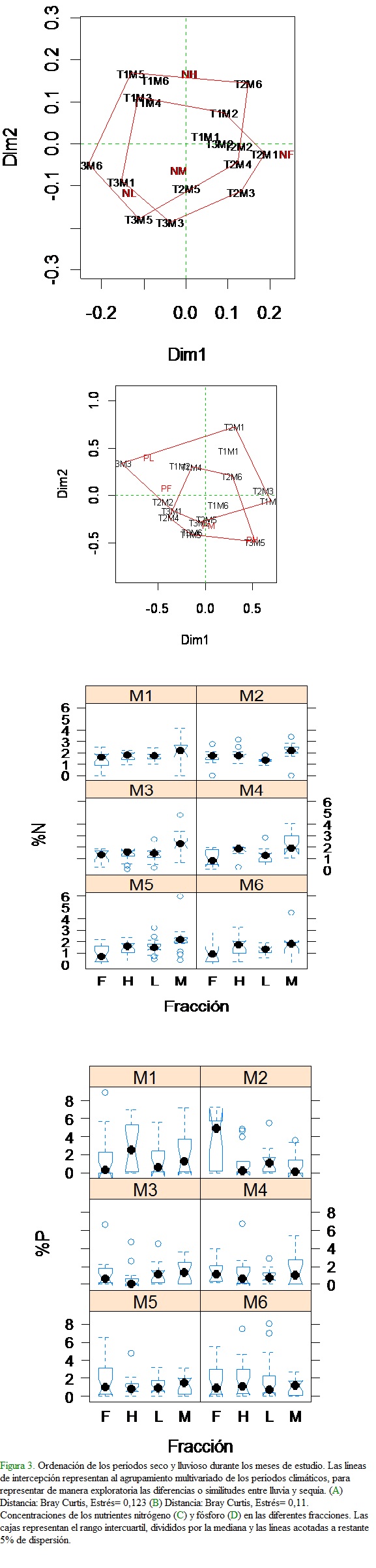
El patrón temporal en el aporte de N presenta una evidencia a su incremento en las fracciones NL y NM, a lo largo de los muestreos que abarcaron los dos períodos climáticos (Figura 3C); por su parte P presentó un patrón más homogéneo en sus contenidos, sin ser afectado por los eventos de precipitación en la cuenca (Figura 3D)
Los retornos potenciales de nutrientes vía hojarasca presentados, mostraron una tendencia hacia valores bajos que coincidieron con el incremento de altitud, donde los picos promedio de los flujos de N y P, se presentan en T3 (N: 43,42 kgha-1año-1 y P: 2,83 kgha1año-1), seguidos por el T2 (N: 40,46 kgha1año-1y P: 2,45 kgha-1año-1) y finalmente en T1 (N: 31,99 kgha-1año-1 y P: 1,13 kgha-1año-1).
Reabsorción de nutrientes y eficiencia en su uso: La menor reabsorción media anual de P se encontró en la formación vegetal establecida en T2 (ERN: 96,78); los valores medios de reabsorción para N fueron similares en las tres formaciones vegetales establecidas en T1, T2 y T3 (ERN: 42,377; ERN: 42,128 y ERN: 42,136 respectivamente). Para T2 la reabsorción de P también fue la menor como ocurrió para el N. La mayor eficiencia en el uso de los nutrientes foliares fue la encontrada para P (IEV: 2888,531) y la menor para N (IEV: 67,023); no obstante, los valores de eficiencia en la reabsorción de nutrientes ERN para N fueron significativos (ERN: 42,218), indicando mayor reabsorción. Asimismo, el nutriente más móvil fue P (ERN: 98,289).
Las formaciones vegetales establecidas en T1 mostraron la mayor eficiencia en el uso de N, seguida de T3 y T2 (Tabla 4); para P se registró en T2, siendo similares entre sí, las otras dos formaciones vegetales, en donde las diferencias no fueron grandes a pesar de presentarse valores altos, lo cual presupone una economía estrecha en el uso del nutriente a pesar de su disponibilidad en las formaciones vegetales.
La formación vegetal establecida en T3 es el más productivo de los aquí estudiados (3132,97 kg·ha-1·año-1) y presenta las mayores concentraciones de N. Sin embargo, las formaciones vegetales establecidas en T2 son las que presenta las mayores concentraciones de P que se retornan al sistema fluvial; al parecer esta situación es producto de menores exigencias de este nutriente por las especies allí establecidas, que no suponen mecanismos de conservación, como es el caso de la reabsorción foliar la cual presenta los valores más bajos en este tramo (96,78%).
DISCUSIÓN
En esta investigación se establece el papel del bosque de ribera del río Gaira, como fuente de aporte de N y P en sus fracciones de hojarasca, las tasas de reabsorción y eficiencia de su uso a través de la hojarasca. Para la región Caribe y específicamente en la sierra nevada de Santa Marta, el presente estudio es el primero en generar información sobre la importancia de estos aportes a escala de tramos y de cuenca, atendiendo a conceptos teóricos y experimentales realizados en latitudes de zonas templadas (McClain et al., 1997), al igual que en la mayoría de las investigaciones para regiones tropicales, la información sobre las concentraciones y reabsorción de nutrientes han sido derivadas de comparaciones entre las hojas y la hojarasca a nivel del bosque (Proctor et al., 1989; Veneklass, 1991).
Concentraciones de N y P a través de la hojarasca: El N que ingresó al bosque de ribera en el tramo T1 (1,39%), T2 (1,63%) y T3 (1,52%), están en el intervalo de los valores reportados para otros bosques tropicales, sólo los valores reportados en el bosque seco de la sierra de Huatla - México (Cárdenas y Campo, 2007) se encuentran por encima de los aportes promedio generados en el presente estudio. A su vez las concentraciones de P evaluadas en los tres tramos de estudio, T1 (0,08%), T2 (0,10%) y T3 (0,13%) están en el intervalo de los estudios realizados en otros bosques tropicales (Tabla 6).
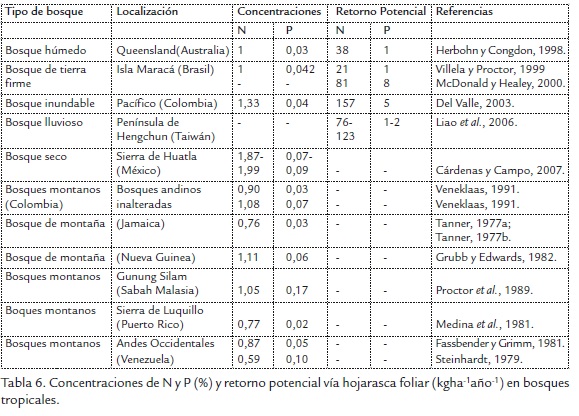
Las concentraciones de N y P presentaron amplia variación, lo que implica mayor inestabilidad y sugiere que su presencia fluctuó en rango amplio durante el estudio, esto puede deberse al efecto del clima y a la baja disponibilidad de nutrientes almacenados en las laderas del río. En este sentido, las concentraciones de los nutrientes fueron en general mayores en los meses menos lluviosos, situación asociada además a los picos de producción de hojarasca.
El patrón de concentración de nutrientes entre las fracciones vegetativas y reproductivas de la hojarasca no es una característica general de todas las especies vegetales sino que existe una amplia variación, convirtiéndose más bien en un conjunto de particularidades en respuesta al tipo de formación vegetal y al patrón fenológico de estas especies que responden a gradientes espaciales, temporales y biogeográficos; tal es el caso de las selvas húmedas de Parama (Golley et al., 1978) donde se encontró una alta variabilidad en las concentraciones de los nutrientes en las fracciones de hojarasca (hojas, frutos y flores) entre el período seco (febrero) y lluvioso (septiembre); sin embargo, en los bosques tropicales de Tanzania, Lundgreen, 1978, se encontró en los aportes, que las concentraciones de los nutrientes (N y P) en la hojarasca siguen el orden de magnitud a las ramas, hojas y órganos reproductivos, similar a los encontrados en nuestro estudio para N (foliar, leñoso y reproductivo), pero contrario al patrón de P (reproductivo, foliar y leñoso).
Las concentraciones promedio de NH del bosque ribereño del río Gaira en los tramos T2 (1,23 %), T1 (1,15%) y T3 (0,98 %), se encuentran en el promedio de los correspondientes a otras formaciones vegetales tropicales, donde las concentraciones reportadas fluctúan entre 0,39 y 1,76 % (Vitousek et al., 1995; Herbohn y Congdon, 1998) estas concentraciones coinciden con todos los aportes encontrados. Contrario a las concentraciones de PH donde las formaciones vegetales estudiadas, superan los valores reportados en los bosques tropicales cercanos al 0,02 % (Vitousek et al., 1995; Cuevas y Lugo, 1998, entre otros).
El valor crítico propuesto por Aerts, 1997, para las relaciones N/P en la hojarasca foliar (11,9) no fue alcanzado en ninguno de los tramos de estudio, en donde esta relación fue de 7,15, 7,86 y 5,34 para T1, T2 y T3 respectivamente, demostrando la ausencia de un déficit de nitrógeno.
RETORNO POTENCIAL DE NUTRIENTES A TRAVÉS DE LA HOJARASCA
El T3 es el más productivo (3132,97 kgha-1año-1) donde fue evidente la correspondencia de mayores aportes de N (43,42 kgha-1año-1) y P (2,83 kgha-1año-1); al parecer esta situación es producto de menores exigencias de estos nutrientes por las especies forestales ahí estudiadas, que no suponen la verificación de mecanismos de conservación, como es el caso de la reabsorción foliar (Londoño et al., 2005; Ramírez et al., 2007). Los retornos de nutrientes presentaron una amplia variación en todos los tramos, donde los retornos de N se correspondieron en su mayoría con los reportados en otros bosques tropicales (Tabla 6) a excepción de las registradas por Del Valle, 2003, en un bosque inundable del pacífico colombiano. Por el contrario, los retornos de P presentaron bajos valores dentro del intervalo de los reportes de bosques tropicales reflejando la baja disponibilidad de este para la nutrición vegetal.
REABSORCIÓN DE NUTRIENTES FOLIARES Y EFICIENCIA DE SU USO
Los aportes bajos de P a través de la hojarasca condujeron a su uso eficiente, reflejado a partir de los altos valores generados en los índices de eficiencia (IEV) y reabsorción (ERN) presentados en las Tabla 4 y Tabla 5, donde los valores IEV registrados para el P coincidieron con los reportes de otros bosques tropicales (Tabla 7) reiterándose el carácter importante que tiene este nutriente para las especies de la vegetación ribereña de la cuenca del río Gaira, al acudir a desarrollo de mecanismos de conservación como lo es la reabsorción foliar, para evitar las pérdidas por inmovilización biológica, entradas alóctonas al cauce, lixiviación, drenajes profundos entre otros (Londoño et al., 2005; Ramírez et al., 2007); al igual que para los valores de ERN los cuales fueron superiores a los reportados por los bosques tropicales (Tabla 7) verificándose una alta eficiencia en su empleo por las especies vegetales por este nutriente.
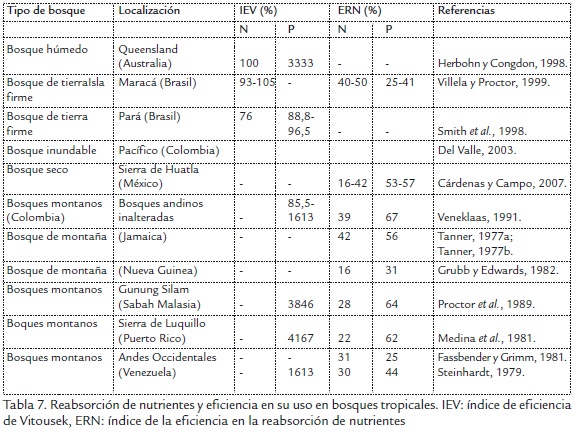
La proporcionalidad encontrada en las formaciones vegetales estudiadas entre las concentraciones de N sugirió un comportamiento similar en cuanto a la reabsorción, donde los valores de las hojas previo a la senescencia y de la hojarasca, se corresponden con los reportados por otros estudios en bosques montanos tropicales con excepción del bosque seco de Brunei - Borneo donde se registraron valores superiores (104-146%) que no se alcanzaron en ninguno de los bosques aquí estudiados; caso similar se presenta para los valores de eficiencia en el uso de N, los cuales se corresponden con los de otros bosques tropicales. Al respecto se ha expuesto que las especies vegetales con baja concentración foliar de un nutriente determinado podían ser usadas como indicador estándar de baja disponibilidad de aquel en un sitio concreto (Vitousek, 1984), como no es este el caso, lo presentado podría estar indicando alta demanda por parte de las especies vegetales y/o alta concentración en el suelo forestal (Tabla 7). Finalmente, los resultados encontrados permiten concluir que el bosque de ribera del río Gaira, es caracterizado por una producción elevada de biomasa (T3), altos niveles de N, pero con mayor eficiencia en el uso de P. La homogeneidad del régimen climático no influye significativamente sobre los descriptores evaluados.
AGRADECIMIENTOS
Los autores agradecen a la División de Investigación de la Universidad Nacional de Colombia (DIB) por la financiación del macro-proyecto "descriptores funcionales de un sistema fluvial de montaña" con el cual se pudo llevar a cabo este trabajo. También se agradece a los grupos de Biodiversidad y Ecología Aplicada de la Universidad del Magdalena, por su apoyo en el desarrollo exitoso del presente trabajo.
BIBLIOGRAFÍA
AERTS R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos. 1997;79:439-449. [ Links ]
ALBARIÑO R. Obtención de indicadores de calidad de sistemas acuáticos de montaña en relación a la actividad forestal, en el marco del proyecto indicadores de manejo forestal sustentable INTA EEA Bariloche; 1999. [ Links ]
ANDERSON J, INGRAM J. Tropical Soil Biology and Fertility: A Handbook Methods. C.A.B. International. Wallingford, Oxford, RU. 1993;221. [ Links ]
AUSSENAC G, BONNEAU M, TACON LE. Restitution des élémentsminérauxau sol par l'intermédiaire de la litiére et des précipitations dans quatre peuplements forestiers de l'Est de la France. OecoZ Plant. 1972;7:1-21. [ Links ]
BENFIELD E. Leaf breakdown in stream ecosystems. In: Hauer FR y GA Lamberti. Methods in stream ecology: Academic Press Inc., London, United Kingdom; 1996;579-589. [ Links ]
BÚRQUEZ, A, MARTÍNEZ A, NÚÑEZ S. Sonoran desert productivity and the effect of trap size on litterfall estimates in dryland vegetation. J Arid Environ. 1999;43:459-465. [ Links ]
CALDENTEY J, IBARRA M, HERNÁNDEZ J. Litter fluxes and decomposition in Nothofaguspumiliostands in the region of Magallanes, Chile. Forest Ecol Manag. 2001;148:145-157. [ Links ]
CÁRDENAS I, CAMPO J. Foliar nitrogen and phosphorus resorption and decomposition in the nitrogen-fixing tree Lysilomamicrophyllumin primary and secondary seasonally tropical dry forests in Mexico. Trop Ecol. 2007;23:107. [ Links ]
CUEVAS E, LUGO A. Dynamics of organic matter and nutrient return from litterfall in stands of ten tropical tree plantation species. Forest Ecol Manag. 1998;112: 263-279. [ Links ]
CUSHING C, CUMMINS K, MINSHALL E. River and stream ecosystems. Amsterdam, The Netherlands, Elsevier; 1995. [ Links ]
DEL VALLE J. Cantidad, calidad y nutrientes reciclados por la hojarasca fina en bosques pantanosos del Pacífico Sur colombiano. Interciencia. 2003;28:449. [ Links ]
ENRIGTH N. Litterfall dynamics in a mixed conifer-angiosperm forest in northern New Zealand. J Biogeogr. 1999;29:149-157. [ Links ]
FASSBENDER H, GRIMM V. Biogeoquímicos en un ecosistema forestal de los Andes Occidentales de Venezuela. II. Producción y descomposición de los estudios vegetales. Turrialba. 1981;31:39-47. [ Links ]
FINZI, AC, ALLEN AS, DE LUCIA EH, ELLSWOTTH DS, SCHLESINGER WH. Forest litter production, chemistry and decomposition following two years of free-air CO2 enrichment. Ecology. 2001;82:470-484. [ Links ]
FLORY E, MILNER A. Influence of riparian vegetation on in vertebrate assemblages in a recent for medstream in Glacier BayNational Park, Alaska. J N Am Benthol Soc. 1999;18:261-273. [ Links ]
FRAYTER V, JIMÉNEZ E, PABÓN R, RIVERA V. Plan de manejo integral de la cuenca hidrográfica del río Gaira. Tesis de Grado. Programa de Ingeniería Agronómica. Universidad del Magdalena. Santa Marta, Colombia; 2000. [ Links ]
GOLLEY F, MCGINMIS J, CLEMENTS A, CHAIL G, DUEYER M. Ciclagem de Mineraisemum Ecossistema de Floresta Tropical Unida. Universidades de Sao Paulo. Brasil; 1978. [ Links ]
GUTIÉRREZ YA. Uso del suelo, vegetación ribereña y calidad del agua de la microcuenca del río Gaira, Santa Marta, Colombia [Tesis de Maestría]. Turrialba, Costa Rica; 2009. [ Links ]
GRUBB P, EDWARDS P. Studies on mineral cycling in a montane rain forest in New Guinea. III. The distribution of mineral elements in the above ground material. J Ecol. 1982;70:623-648. [ Links ]
HERBOHN J, CONGDON R. Ecosystem dynamics at disturbed and undisturbed sites in north Queensland wet tropical rain forest. III. Nutrient returns to the forest floor through litterfall. J Trop Ecol. 1998;14:217-229. [ Links ]
HERNÁNDEZ J. Modificaciones en el aporte y la descomposición de hojarasca por la intervención silvícola en bosques naturales de lenga (Nothofaguspumilio). Memoria de Título, Facultad de Ciencias Forestales, Universidad de Chile, Santiago; 2000. [ Links ]
HIROSE T. Relations between turnover rate, resource utility and structure of some plant populations. A study of the matter budgest. J Fac Sci Tio Univ sect III.1975;11:355-407. [ Links ]
KLUTE A. Methods of soil analysis. Part 1. Physical and mineralogical methods. 2nd ed. Am. Soc. Agron. / Soil Sci Soc Am. Madison, WI, EEUU; 1986;1188. [ Links ]
LANDSBERG J, GOWER S. Applications of Physiological Ecology to Forest Management. NY, EEUU, Academic Press; 1997;345. [ Links ]
LEÓN P, GONZÁLEZ H, GALLARDO L. Retranslocación y eficiencia en el uso de nutrientes en bosques del centro de Antioquia; 2009. [ Links ]
LIAO JH, WANG HH, TSAI CHCH, HSEU ZY. Litter production, descomposition and nutrient return of uplifted Coral Reef tropical forest. Forest Ecol Manag. 2006;235: 4-185. [ Links ]
LONDOÑO A, MONTOYA D, LEÓN J, GONZÁLEZ M. Ciclaje y pérdida de nutrientes del suelo en bosques altoandinos de Antioquia, Colombia. Rev Fac de Agronomía. 2005;60:3717-3734. [ Links ]
LUH HUANG C, SCHULTE E. Digestion of plant tissue for analysis by ICP emission spectroscopy. Comm Soil Sci Plant. 1985;16:943-958. [ Links ]
LUNDGREN B. Soil conditions and nutrient cycling under natural and plantation forests in Tanzanian highlands. Reports in Forest Ecology and Forest Soils 31. Swedish University of Agricultural Sciences, Upssala, Suecia; 1978;428. [ Links ]
MALANSON G. Riparian landscapes. Cambridge University Press, Cambridge, United Kingdom;1995. [ Links ]
MCCLAIN M, RICHEY J, BRANDES J, PIMENTEL T. Dissolved organic matter and terrestrial-lotic linkages in the central Amazon Basin, Brazil. Global Biogeochem C Y. 1997;11:295-311. [ Links ]
MCDONALD MA, HAELEY JR. Nutrient cycling in secondary forests in the Blue Mountains of Jamaica. Forest Ecol Manag. 2000;139:257-278. [ Links ]
MEDINA E, CUEVAS E, WEAVER P. Composición foliar y transpiración de especies leñosas del Pico del Este, Sierra de Luquillo, Puerto Rico. Acta Cient Venezolana. 1981;32:159-165. [ Links ]
PAUL M, HALL R. Particle transport and transient storage along a stream-size gradient in the Hubbard BrooK Experimental. J N Am Benthol Soc. 2002;21:195-205. [ Links ]
PROCTOR J. Tropical forest litterfall. I. Problems of data comparison. En: Sutton SL, Whitmore TC, Chadwick AC (Eds.) Tropical Rain Forest: Ecology and Management. Blackwell Scientific Publications; Oxford RU; 1983;267-273. [ Links ]
PROCTOR J, PHILLIPS C, DUFF G, HEANEY A, ROBERTSON F. Ecological studies on Gunung Silam, a small ultrabasic mountain in Sabah, Malaysia. II. Some forest processes. J Ecol. 1989;77:317-331. [ Links ]
RAI, S, PROCTOR J. Ecological studies on four rainforests in Karnataka, India. II. Litterfall. J Ecol. 1986;74:455-463. [ Links ]
RAMÍREZ J, ZAPATA C, LEÓN J, GONZÁLEZ M. Caída de hojarasca y retorno de nutrientes en bosques montanos Andinos de Piedras Blancas, Antioquia, Colombia. Revista Interciencia. 2007;32:303-311. [ Links ]
RÍOS S, BAILEY R. Relationship between riparian vegetation and stream benthic communities at three spatial scales. Hydrobiologia. 2006;553:153-160. [ Links ]
RODRÍGUEZ J. Dinámica de la materia orgánica en la cabecera de una quebrada de montaña. Bogotá, Colombia [Tesis de Maestría]. Universidad Nacional de Colombia; 2006;96. [ Links ]
SIMON K, BENFIELD E. Leaf and woodbreakdown in cave streams. J.N. Am. Benthol. Soc. 2001;20: 550-563. [ Links ]
SMITH CK, GHOLZ HL, OLIVEIRA F. Litterfall and nitrogen-use efficiency of plantations and primary forest in the eastern Brazilian Amazon. Forest Ecol Manag. 1998;109:209-220. [ Links ]
STEINHARDT U. Untersuchungenuber den Wasser and Nahrstoffhaushalteinesandien Wolkenwaldes in Venezuela. Gottinger Bodenkundliche Berichte 1979;56:185. [ Links ]
STOCKER G, THOMPSON W, FITZSIMON A, THOMAS P. Annual patterns of litterfall in lowland and tableland rainforest in topical Australia. Biotropica. 1995;27:412-420. [ Links ]
TABATABAI M, BREMNER J. Automated instruments for determination of carbon, nitrogenand sulfur in soils by combustion techniques. En Soil analysis, modern instrumental techniques. 2nd ed. Nueva York, EEUU, Dekker; 1991. [ Links ]
TAMARÍS TCE, LÓPEZ HJS. Aproximación a la zonificación climática de la cuenca del río Gaira. Rev Intrópica. 2006:69-76. [ Links ]
VENEKLAAS E. Litterfall and nutrient fluxes in two montane tropical rainforest.Colombia. J Trop Ecol. 1991;7:319-336. [ Links ]
TANNER EVJ. Four montane rain forest of Jamaica: a quantitative characterization of the floristics, the soils, and the foliar mineral levels, and a discussion of the interrelations. J. Ecol. 1977a;65:883-918. [ Links ]
TANNER EVJ. Mineral cycling in montane rain forest in Jamaica [Ph. D. Thesis]. Cambridge, UK; University of Cambridge; 1977b. [ Links ]
VILLELA D, PROCTOR J. Litterfall mass, chemistry, and nutrient retranslocation in a monodominant forest on Maracá Island, Roraima, Brazil. Biotropica 1999;31:198-211 [ Links ]
VITOUSEK P. Litterfall, nutrient cycling and nutrient limitation in tropical forests. Ecology. 1984;65:285-298. [ Links ]
VITOUSEK P, GERRISH G, TURNER D, WALKER L, MÜELLER-DOMBOIS D. Litterfall and nutrient cycling in four Hawaiian montane rainforests. J Trop Ecol. 1995;11:189-203. [ Links ]

