










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.17 no.1 Bogotá Jan./Apr. 2012
DESCOMPOSICIÓN DE LA HOJARASCA Y SU RELACIÓN CON LOS MACROINVERTEBRADOS ACUÁTICOS DEL RÍO GAIRA (SANTA MARTA - COLOMBIA)
Leaf Litter Decomposition and its Relation with Aquatic Macroinvertebrates of the Gaira River (Santa Marta - Colombia)
MELISA EYES-ESCALANTE1, M.Sc.; JAVIER RODRÍGUEZ-BARRIOS2, M.Sc.; LUIS CARLOS GUTIÉRREZ-MORENO1, M.Sc.
1 Grupo Biodiversidad del Caribe Colombiano, Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico, km 7, vía Puerto Colombia, Barranquilla, Colombia. meyesescalante@hotmail.com
2 Grupo de Biodiversidad y Ecología Aplicada, Instituto de Investigaciones Tropicales, Universidad del Magdalena. jarodriguezba@gmail.com Correspondencia: Melisa Eyes-Escalante. Teléfono: 378 03 63 Celular: 313 565 30 47. Carrera 41D # 84A-49, Barranquilla, Atlántico. Colombia. meyesescalante@hotmail.com
Presentado 3 de marzo de 2011, aceptado 2 de noviembre de 2011, correcciones 9 de noviembre de 2011.
RESUMEN
Se evaluó la tasa de descomposición de la hojarasca, de cuatro especies nativas y tres especies exóticas, seleccionadas por su mayor frecuencia en la vegetación de la ribera del río Gaira, localizado en la Sierra Nevada de Santa Marta. Se seleccionaron cuatro tramos de muestreo a diferentes alturas. Adicionalmente, se determinaron riqueza, abundancia y función trófica alimenticia de los macroinvertebrados que colonizaron los sustratos artificiales que contenían la hojarasca. En el tramo medio se presentó el mayor coeficiente de descomposición de especies nativas y exóticas (K-W=15,9; n= 23; p= 0,001 y KW=15,5; n=36; p=0,0014 respectivamente). Las especies nativas presentaron la mayor tasa de descomposición (K-W=4; n=47; p=0,04), siendo la especie Pithecellobium longifolium en el tramo medio, la que presentó la mayor pérdida de peso en el tiempo. Se presentan relaciones entre tasa de descomposición, conductividad y temperatura. El ensamblaje de macroinvertebrados estuvo representado por 41 taxones, con una abundancia de 1895 individuos que aportaron una biomasa de 2,27 g. La estructura funcional de los macroinvertebrados asociados a la hojarasca, estuvo representada en los tramos alto, medio y bajo por Plecopteros (Depredadores) del género Anacroneuria con 10,14%, 68,43% y 7,60% respectivamente. Dípteros depredadores del taxón Tanypodinae fueron dominantes en la desembocadura con 0,51%. Los resultados de este estudio permiten determinar que la presencia de macroinvertebrados asociados a la hojarasca no está condicionada por la especie vegetal (nativa o exótica), porque no se observaron diferencias entre estas que generen un efecto sobre la tasa de descomposición.
Palabras clave: descomposición, nativa, exótica, macroinvertebrados, biomasa. (Santa Marta Colombia). Eyes-Escalante, et ál.
ABSTRACT
The rate of decomposition of leaf litter of four native species and three exotic species were evaluated. They were selected due to the most frequently collected by the bank vegetation Gaira River, located on the Sierra Nevada de Santa Marta. We took four sampling sections at different heights. Furthermore, it was evaluated the richness, abundance and trophic function of macroinvertebrates that colonized the artificial substrates containing leaf packs or meshes. The middle part had the greatest rate of decomposition of native and exotic species (KW = 15.9, n = 23, p = 0.001 and KW = 15.5, n = 36, p = 0.0014 respectively). The native species had the highest rates (KW = 4, n = 47, p = 0.04), with the species Pithecellobium longifolium in the middle stretch, which had the greatest mass loss over time. Descriptively a relationship between decomposition rate and current velocity is presented. The macroinvertebrate assemblage was represented by 41 taxa, with an abundance of 1895 individuals which provided a biomass of 2.27 g. The functional structure of macroinvertebrates associated with leaf litter was represented in the upper, middle and low reaches by Plecoptera (Predators) of the genus Anacroneuria with 10.14%, 68.43% and 7.60% respectively. The taxon Tanypodinae (Diptera) predators were dominant in the mouth with 0.51%. The result of this study can determinate whether the presence of macroinvertebrates associated with leaf litter, is not conditioned by the plant species (native or exotic), and not enter these differences that create an effect on the rate of descomposition.
Key words: Descomposition, native, exotic, macroinvertebrates, biomass.
INTRODUCCIÓN
La principal fuente de materia orgánica que entra a ríos pequeños y medianos, proviene de la hojarasca de la vegetación de las riberas (Vannote et al., 1980), (Pozo et al., 1997), (Hoffman, 2005), aportando hasta 99% de la energía que entra a estos ríos (Fisher y Likens, 1973; Winterbourn, 1976; Benfield, 1997). El aporte y descomposición de hojarasca es uno de los procesos más estudiados en ríos pequeños en donde la vegetación de ribera es abundante, porque una vez las hojas ingresan al río comienza un proceso de descomposición por factores mecánicos, biológicos y fisicoquímicos que permiten que el material depositado sea aprovechado en forma inmediata.
Las metodologías propuestas para calcular tasas de descomposición de las hojas de las plantas terrestres en los ríos se han sugerido desde hace algún tiempo como una medida integrada para estudiar la salud de los ecosistemas acuáticos (Webster y Benfield, 1986; Vymazal, 1995; Qualls y Richardson, 2000). Cabe resaltar que este tipo de trabajos juega un papel importante debido a que su enfoque ecológico genera información relevante para futuros estudios que estén encaminados a tomar acciones de manejo adecuado para este tipo de ecosistemas.
Los estudios realizados por González y García, 1998, demuestran que existe una relación importante entre los flujos energéticos de los ríos con la estructura de macroinvertebrados, porque estos son organismos que representan un grupo muy diverso y cumplen con una variedad de funciones amplia dentro del ecosistema acuático como descriptores de posibles alteraciones en términos de su abundancia y biomasas. En el Neotrópico los estudios de Graça et al., 2002, señalan la importancia de los macroinvertebrados en la descomposición de hojarasca principalmente en ríos de primer orden, porque esta fauna béntica puede acelerar los procesos de fragmentación, facilitando la disolución de los compuestos químicos de las hojas y la colonización de otros microorganismos. El objetivo del presente trabajo es evaluar la variación en la descomposición de hojarasca de especies nativas y exóticas y su relación con los macroinvertebrados acuáticos en un gradiente altitudinal del río Gaira, Santa Marta, Colombia, asociado a variables biológicas y ambientales.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
El estudio se llevó a cabo en la cuenca del río Gaira, ubicado en la vertiente noroccidental de la Sierra Nevada de Santa Marta (SNSM), en el departamento del Magdalena, Colombia entre 11°52’06" y 11°10’08" latitud Norte y 74°46’22" y 74°11’07" latitud Oeste. El río nace a una altura de 2750 m, en una zona de transición entre bosque muy húmedo subtropical y bosque muy húmedo montano bajo (Tamarís-Turizo y López, 2006), con una longitud aproximada de 32,53 km desde su nacimiento hasta su desembocadura en el mar Caribe (Frayter et al., 2000).
ESCALA ESPACIO-TEMPORAL
En el río se seleccionaron cuatro tramos a lo largo de un gradiente altitudinal, el primer sitio (tramo alto) se ubicó en el sector de San Lorenzo a 1600 msnm, localizado entre los 11°10’2" N y 74°10’41,5" W. El segundo sitio (tramo medio) se ubicó en el sector de la hacienda La Victoria a 900 msnm, localizado entre los 11°07’44,2" N y 74°05’35,8" W. El tercer sitio (tramo bajo) se ubicó en el sector de puerto Mosquito a 50 msnm, localizado entre los 11°10’26’’ N, 74°10’37’’ W y el cuarto sitio (desembocadura) se ubicó en el sector del Rodadero sur a 2 msnm, localizados entre los 11°11’33,57’’ N, 74°13’ 52,38’ W. Se realizaron cuatro muestreos entre febrero y mayo de 2009, los cuales consistieron en la postura y recolecta de mallas con hojas de especies nativas y exóticas para evaluar el coeficiente de descomposición de la hojarasca que entra al río siguiendo el modelo de investigación propuesto por Elosegi y Sabater, 2009. El primer muestreo (M1) se realizó a los siete días posteriores a la postura de las mallas, el segundo (M2) a los 14 días, el tercero (M3) a los 28 días y el cuarto (M4) a los 56 días.
TOMA DE DATOS
En cada tramo seleccionado se colocaron seis experimentos que contenían ocho mallas de 20 x 20 cm con poro de 0,5 cm; de tal manera que cuatro mallas eran para la hojarasca de la especie nativa y cuatro para la especie exótica. En el tramo alto se colocaron hojas de: Cecropia peltata (conocido como guarumo, es nativo para esta zona y su distribución va desde zonas bajas hasta 1600 msnm) y Eucaliptus camaldulensis (conocido como eucalipto, especie introducida para esta zona, nos basamos en un anterior estudio realizado en el río Gaira) (Guerrero y Zúñiga, 2005). En el tramo medio: P. longifolium (conocido como guamo de río y considerado una especie nativa) y Coffea arabica (café, especie introducida). En el tramo bajo: Anacardium excelsum (conocido como caracolí, el cual no se extiende por encima de 900 msnm) y Mangifera indica (mango, un frutal introducido a estas zonas de río y playa) y en la desembocadura: Avisenia germinans (conocido como mangle salao, propio de esta zona de desembocadura) y M. indica. En total se colocaron 24 mallas por cada especie vegetal con un peso inicial de 5 g. La selección de las especies nativas o exóticas fue basada en la distribución de estas de acuerdo a la altura y a su presencia original o introducida históricamente en la vegetación ribereña del río Gaira.
VARIABLES HIDROLÓGICAS
En cada tramo muestreado se midieron variables hidrológicas correspondientes a la velocidad de la corriente y caudal. Simultáneamente se midieron variables fisicoquímicas como: temperatura, pH, oxígeno disuelto, conductividad y sólidos disueltos totales. Los datos de precipitación se tomaron con base en los registros de las estaciones meteorológicas correspondientes a: tramo alto: estación 1501506 San Lorenzo, tramo medio: estación de Minca y tramo bajo y desembocadura: estación 1501505 aeropuerto Simón Bolívar.
FASE DE LABORATORIO
Las muestras de hojarasca se lavaron cuidadosamente sobre tamices de 180 y 63 µm dispuestos consecutivamente con el fin de recolectar macroinvertebrados. Posteriormente se secaron las muestras a 60 °C en estufa y se pesaron en una balanza analítica obteniendo el peso final de la muestra.
Para determinar variación, composición y abundancia de macroinvertebrados que colonizan y se establecen en la hojarasca en descomposición, se realizó la determinación taxonómica de los especímenes utilizando las claves taxonómicas de Merrit y Cummins, 1996. La asignación de los grupos funcionales alimenticios (GFA) se realizó de acuerdo a la clasificación propuesta por Tomanova et al., 2006 y Chará et al., 2010, agrupándolos de acuerdo al grupo funcional alimenticio, así: Depredadores (D), Colectores Filtradores (CF), Colectores Recolectores (CR), Raspadores (R), Trituradores (T). La biomasa se estimó mediante regresiones de longitud vs. peso de cada taxón identificado, utilizando las regresiones propuestas por Benke et al., 1999 y Cressa, 1999.
ANÁLISIS ESTADÍSTICO
Se calculó el coeficiente de descomposición acuática de la hojarasca utilizando el modelo exponencial negativo propuesto por Petersen y Cummins, 1974: Mt = Mi e -kt, en donde Mt es la masa remanente en el tiempo t, Mi es la masa inicial, -k es el coeficiente exponencial de descomposición y t el lapso de tiempo durante el cual se lleva el proceso. Se realizó una comparación entre los coeficientes de descomposición obtenidos a través de una prueba de Kruskal-Wallis (Programa Statgraphics Centurion XV). Se determinaron la composición y estructura de la comunidad de macroinvertebrados, se establecieron los GFA y los aportes de biomasa y se realizó un análisis de varianza multivariado no paramétrico (PNMANOVA) para observar variaciones entre tramos, muestreos y condición vegetal (hojarasca de especies nativas y exóticas). El nivel de asociación entre la tasa de descomposición de la hojarasca y el ensamblaje de macroinvertebrados presentes con las variables ambientales se estableció mediante un análisis BIOENV (Legengre y Legengre, 1998) utilizando correlaciones de Spearman y la distancia Bray Curtis. Para los análisis estadísticos se utilizó el paquete R versión 2.11.1 (R Development Core Team 2010 disponible en www.r-proyect.org).
RESULTADOS
CARACTERIZACIÓN DEL RÍO GAIRA
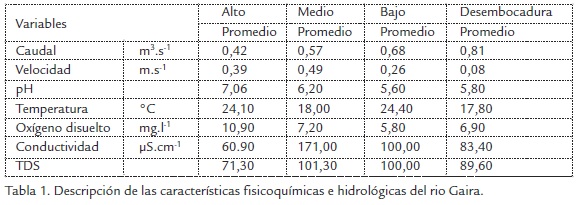
Los datos arrojados por el IDEAM mostraron que los valores de precipitación co- rrespondiente a M1 y M2 en cada unos de los tramos estudiados estuvieron marcados por un período seco. Para los muestreos M3 y M4 los valores registraron un período de lluvias (Figura 1). El caudal (m3.s-1) muestra un gradiente ascendente de los volúmenes de agua desde las zonas altas (tramo alto) hasta la desembocadura, el cual se duplica en el recorrido del río dentro del área objeto de estudio desde 0,42 a 0,81 m3s-1. La velocidad de la corriente mostró un flujo heterogéneo en el sistema, porque se observó el mayor valor promedio en el tramo medio (0,49 ms-1) y el menor valor en la desembocadura (0,08 ms-1).
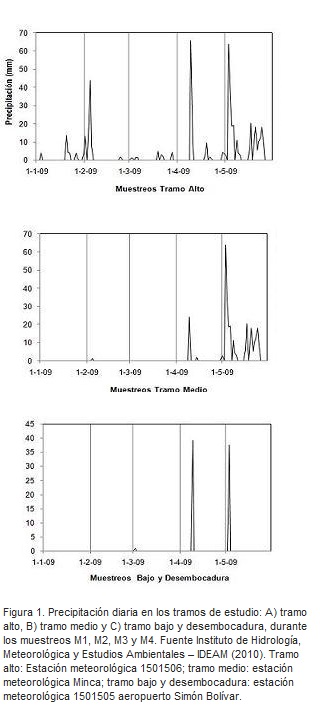
En relación con las características fisicoquímicas del agua, la temperatura varió de un tramo a otro, mostrando un comportamiento influenciado por la morfología propia de cada sitio, esto explica los comportamientos de pH y oxígeno, los cuales siguieron un patrón similar. En cuanto a los valores de conductividad y TDS estos fueron proporcionales a los niveles registrados de oxígeno disuelto en los distintos tramos estudiados (Tabla 1). Desde el punto de vista fluvial se observó, que los datos no siguieron un patrón marcado por el gradiente altitudinal, sino por características de cada tramo, tales como: vegetación, lecho rocoso que caracterizó a cada tramo y desembocadura en el mar Caribe.
COEFICIENTE DE DESCOMPOSICIÓN DE HOJARASCA DE ESPECIES NATIVAS Y EXÓTICAS
El coeficiente de descomposición de hojarasca de especies nativas presentó diferencias al considerar el gradiente altitudinal en la cuenca del río Gaira (K-W=15,5; n=36; p=0,0014), registrándose la mayor tasa de descomposición en el tramo medio y la menor en la desembocadura. Por otro lado, durante el tiempo de exposición de la hojarasca, no se presentaron diferencias en el tramo alto (K-W=0,88; n=20; p=0,82).
Lo contrario se observó en el tramo medio (K-W=9,44; n=20; P=0,02), el tramo bajo (K-W=8,23; n=21; p= 0,04) y en la desembocadura (K-W=8,59; n=22; p=0,03). Considerando los valores absolutos y los máximos entre los intervalos de tiempo se observó un comportamiento ascendente en los tramos alto, medio y bajo. En la desembocadura se evidencia un comportamiento diferente donde la tasa disminuye desde el intervalo de tiempo 1, 2 y 3, con un leve incremento en el intervalo de tiempo 4 (Tabla 2).
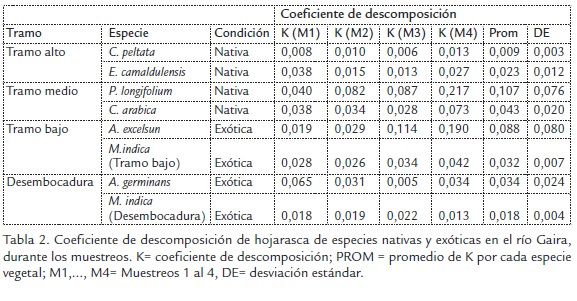
En cuanto al coeficiente de descomposición de la hojarasca de especies exóticas este presentó diferencias a lo largo del gradiente altitudinal (K-W=15,9; n=23; p=0,001). En cuanto a los registros de los intervalos de tiempo no se presentaron diferencias. En el tramo alto (K-W= 7,19; n= 18; p= 0,06), tramo bajo (K-W= 1,18; n= 20; p= 0,75) y desembocadura (K-W= 6,02; n= 22; p= 0,11) a exepción del tramo medio donde si se observaron diferencias (K-W= 12,8; n= 21; p= 0,0048). Se observó que entre la condición nativa y exótica se presentaron diferencias en la velocidad de descomposición (K-W=4; n=47; p=0,04); siendo las hojarascas de especies nativas las que se descompusieron más rápidamente en los tramos medio, bajo y desembocadura.
Para determinar el mejor subgrupo de variables ambientales, que definen la tasa de descomposición de la hojarasca (BIOENV), se encontró que la temperatura y la conductividad son las que presentan el mayor nivel de asociación (rs= 0,46, p< 0,005).
ENSAMBLAJE DE MACROINVERTEBRADOS ASOCIADOS A HOJARASCA DE ESPECIES NATIVAS Y EXÓTICAS EN EL RÍO GAIRA
Patrón de riqueza, abundancia y biomasa. Se recolectaron 1895 ejemplares distribuidos en 12 órdenes, 32 familias y 41 taxones, con un aporte en biomasa de 2,27 g. A nivel de tramos el taxón más abundante fue Tanypodinae con 1101 individuos que representan 58%, seguido de Chironominae con 147 individuos que representan 8% y Leptohyphes sp. con 142 individuos que representan 7% del total. En cuanto a biomasa el taxón dominante correspondió a Anacroneuria con 1,817 g que representan 80% de la biomasa aportada (Tabla 3).
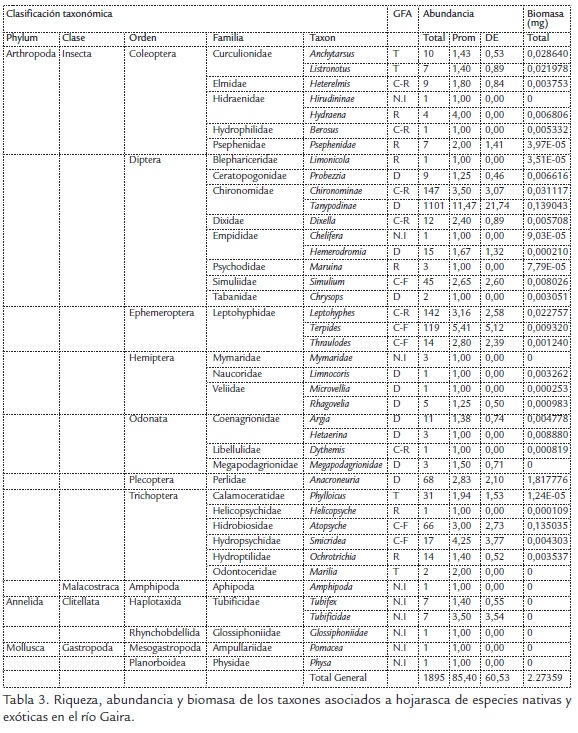
En relación con los tramos el mayor valor de abundancia y biomasa se presentó en el tramo medio con 682 individuos que representan 36% y 1,65 g que representa 72% de la biomasa asociada al material vegetal expuesto en el experimento. La menor abundancia se presentó en la desembocadura con 153 individuos que representan 8% y 0,024 g que corresponde a 1% (Tabla 4). El análisis de NPMANOVA, evidenció que existen diferencias significativas en la biomasa de macroinvertebrados entre los tramos (SeudoF: 2,6, g.l.= 3, p = 0,01).
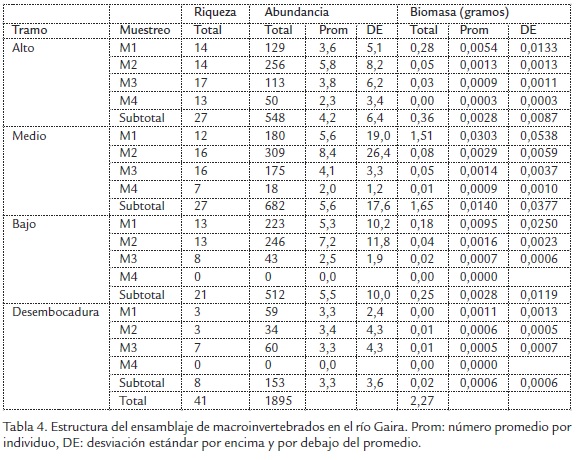
En cuanto al patrón temporal, los mayores valores de riqueza y abundancia se registraron en M2 con 27 taxones y 845 individuos que representan 45% del total recolectado. El menor valor se presentó en M4 con 18 taxones y 68 individuos que representan 3% del total recolectado, en cuanto a la biomasa registrada, durante M1 se presentó el mayor valor (1,97) que representa 87% y el menor valor se presentó en M4 (0,01 g) que representa menos de 1% (Tabla 3). El análisis NPMANOVA (SeudoF: 1,81, g.l.= 3, p = 0,01) evidenció que existen diferencias significativas entre las medias de la abundancia de macroinvertebrados en el tiempo.
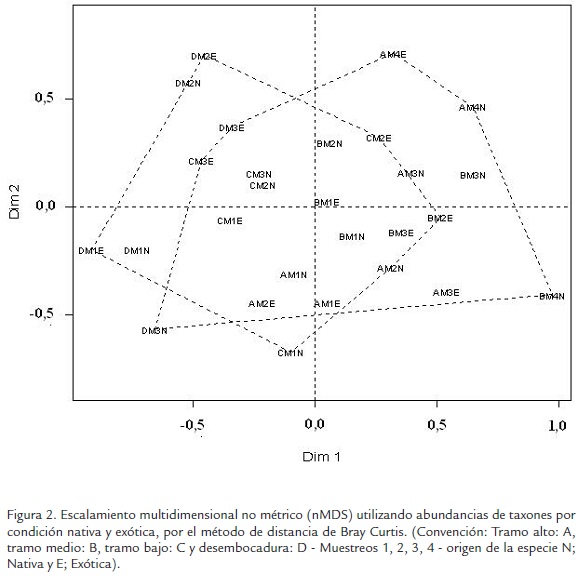
En cuanto a la preferencia de los macroinvertebrados asociados a la hojarasca, por condición vegetal (nativas y exóticas) no se observaron diferencias, NPMANOVA (SeudoF= 0,03, gl= 1, p= 0,86). Se recolectaron 989 individuos en la hojarasca de especies nativas que representan 52% y para la hojarasca de especies exóticas 906 individuos que representaron 48%. La ordenación de macroinvertebrados en las diferentes localidades y condición nativa - exótica, confirmó que la composición y estructura de estos organismos presentó solapamiento entre la hojarasca de especies vegetales nativas y exóticas (Figura 2), indicando homogeneidad en el ensamblaje de estos componentes vegetales.
VARIACIÓN DE LOS GFA
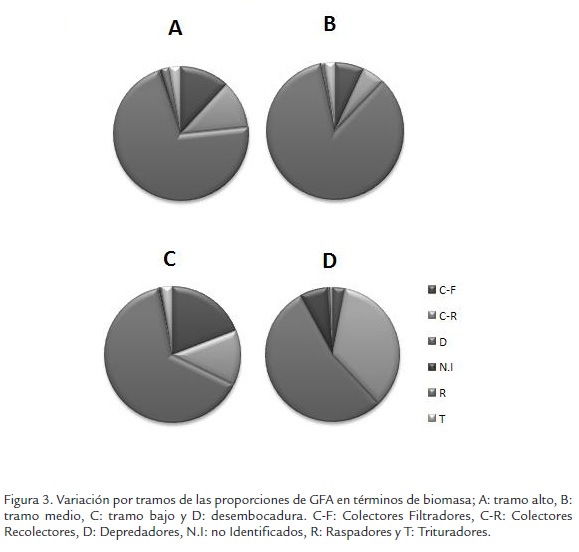
En términos de biomasa los resultados mostraron que el mayor valor de biomasa correspondió a depredadores (D), este grupo presentó 0,26 g en el tramo alto, que corresponden a 71%, tramo medio (1,5 g) que corresponden a 85%, tramo bajo (0,19 g) que corresponden a 64% y el desembocadura (0,010 g) que corresponden a 54% (Figura 3).
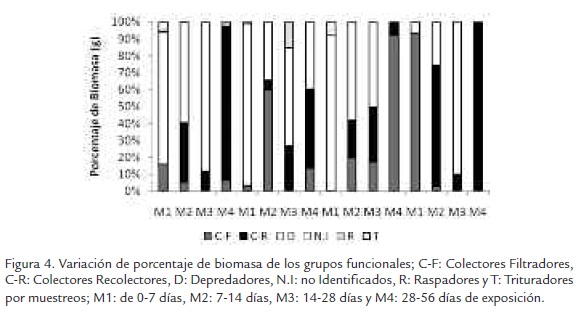
En relación con los muestreos en el tramo alto se presenta una tendencia sucesional marcada por los depredadores al inicio de los muestreos, en donde los C-R van ocupando espacio hasta predominar hacia el último muestreo. En el tramo medio la dominancia está marcada por depredadores en M1 seguida en M2 por colectores filtradores. En el tramo bajo se observó como los colectores filtradores van apareciendo a partir de M2 hasta predominar en M4. En la desembocadura ocurre lo contrario a lo expuesto en los anteriores tramos en donde la dominancia inicialmente está marcada por los colectores filtradores los cuales en M2 y M4 son remplazados por depredadores (Figura 4).
Para determinar el mejor subgrupo de variables ambientales, que definen el ensamblaje de macroinvertebrados (BIOENV que la temperatura, la conductividad y la velocidad de la corriente son las que presentan el mayor nivel de asociación (rs= 0,59, p< 0,005).
DISCUSIÓN
CARACTERIZACIÓN DEL RÍO GAIRA
El comportamiento del caudal según el gradiente altitudinal de la cuenca, es típico de las escorrentías en ambientes de montaña, donde el área de recarga del río se amplifica desde el nacimiento hasta la desembocadura. Otro aspecto que determina este comportamiento es la estructura del sistema hidráulico de la cuenca, donde el río en su recorrido recibe un mayor volumen de agua desde los afluentes que se incorporan al canal principal (escorrentías segundarias y terciarias).
Es importante resaltar que el caudal en el río se duplica desde las zonas altas hasta la desembocadura. Sin embargo, este comportamiento es inverso a la velocidad de la corriente, estos resultados pueden estar asociados a aspectos del relieve y de la topografía de la cuenca, donde los tramos o zonas con mayor pendiente aceleran la movilidad del agua o la velocidad de caída por efecto de la fuerza de gravedad. Este aspecto es relevante en la medida en que existen fenómenos mecánicos asociados a la descomposición de hojarasca en medios lóticos donde la fragmentación de material facilita procesos químicos y la acción biológica, tal como el establecimiento y desarrollo de fauna béntica (Allan, 1995). Gordon et al., 1992, introdujeron el concepto de hábitat hidráulico para definir esta clase de ambientes, en donde, muchos grupos de invertebrados han desarrollado estructuras morfológicas que les permiten soportar diferentes velocidades de la corriente.
DESCOMPOSICIÓN DE HOJARASCA Y EFECTO DE LA CONDICIÓN DE ORIGEN DE LAS ESPECIES VEGETALES (NATIVA Y EXÓTICA)
La tasa de descomposición de hojarasca de especies nativas y exóticas presentó diferencias según el gradiente altitudinal en el río Gaira; estas diferencias pueden estar asociadas a factores mecánicos como la velocidad de la corriente, proceso al que se le atribuye la fragmentación física que sufren las hojas. Algunos autores plantean que este es el principal agente de descomposición de hojarasca (Campbell et al., 1992), lo cual es consistente en este estudio, con los valores de mayor tasa de descomposición de hojarasca de especies nativas y exóticas en el tramo medio y la mayor velocidad de la corriente del agua registrada en los tramos alto y medio. Los resultados evidencian un efecto de la condición nativa y exótica sobre la tasa de descomposición, donde la hojarasca de especies nativas se descompone con más rapidez que las exóticas, resultados similares a los reportados por Guerrero y Zuñiga, 2005, en un estudio realizado en el tramo medio de río Gaira.
ENSAMBLAJE DE MACROINVERTEBRADOS ASOCIADOS A HOJARASCA DE ESPECIES NATIVAS Y EXÓTICAS EN EL RÍO GAIRA
Los tramos de estudio se caracterizan por la presencia del grupo funcional de colectores, los cuales presentaron en términos de biomasa un aporte bajo; sin embargo, el suministro de estos favoreció la presencia de depredadores, los cuales aprovechan el suministro orgánico que estos le proveen (Fisher y Gray, 1983), (Wallace y Webster, 1996).
Los datos reportados demuestran que el río Gaira es un ecosistema fluvial con una diversidad y abundancia de macroinvertebrados significativa, con 1895 ejemplares, agrupados en 41 táxones, distribuidos en 32 familias, cifras que superan otros estudios realizados en los tramos medio y bajo por Rueda-Delgado et al., 2006, donde registraron un mayor número de individuos, distribuidos en 27 familias.
Tal como lo registran los estudios de Ramírez y Pringle, 2001, Rueda-Delgado, 2005, Gutiérrez, 2006, Rodríguez, 2006, Rodríguez-Barrios et al., 2007, realizados en ríos de Colombia, los dípteros seguidos por los efemerópteros son los que mayor abundancia aportan en esta clase de ambientes loticos. Sin embargo, en cuanto a su aporte en biomasa estos no superan 10%.
En relación con los GFA depredadores y colectores recolectores CR dominaron en términos de riqueza y abundancia, la biomasa fue dominada por los depredadores. Chara et al., 2010, argumentan que aunque el grupo de los pequeños colectores es abundante su importancia ecológica no supera a otros grupos con menos abundancia pero con mayor aporte de biomasa, como es el caso de los depredadores.
En términos de biomasa los depredadores dominaron en 82%, jugando un papel importante en estos ríos, tal como se expone en los estudios realizados por Cheshire et al., 2010, donde reportan en ríos australianos la importancia de fragmentadores y depredadores. Para Colombia Chara et al., 2010, reportan que 64% y 19% corresponde a fragmentadores y depredadores. La disminución del grupo de colectores en este estudio coincide con los trabajos de Greathouse y Pringle, 2005, los cuales contradicen las predicciones realizadas por Vannote et al., 1980 y Grubaugh et al., 1996, en donde no registran un efecto de la altura sobre estos grupos.
La baja proporción del grupo de los trituradores (fragmentadores) en este estudio la cual no supera el 2% de la biomasa aportada coincide con los trabajos realizados por Ramírez et al., 1998, Greathouse y Pringle, 2005, Gutiérrez, 2006, Rueda-Delgado et al., 2006 y Wantzen et al. 2002, los cuales atribuyen este comportamiento a que su función es remplazada por raspadores y colectores que en este estudio aportaron 1% y 10% respectivamente.
Asociación entre las variables biológicas y las variables ambientales El BIOENV realizado permitió establecer que la temperatura, conductividad y velocidad de la corriente son los mejores descriptores, pues generaron el mayor efecto sobre la tasa de descomposición y ensamblaje de macroinvertebrados acuáticos. La velocidad de la corriente fue una variable que influyó de manera significativa sobre el ensamblaje de macroinvertebrados, en cuanto a las variaciones que estos organismos presentaron entre tramos, coincidiendo con registros en otros ecosistemas tropicales, en donde se ha podido relacionar el efecto de la velocidad en zonas de rápidos, con la mayor riqueza y diversidad de taxones del bentos (Ríos, 2008), presentándose un efecto positivo y significativo entre la magnitud de esta variable hidrológica sobre la densidad en estos organismos (Brttain y Eikeland, 1988; Boyero y Bosch, 2002).
CONCLUSIONES
La variación del caudal del río Gaira está dinamizada por las lluvias locales, por lo tanto no se detecta un patrón definido en relación a los muestreos según su categorización como periodo de lluvia o seco, en la medida en que esta categorización corresponde a los históricos climáticos, el régimen de caudales observado es típicos de los ríos de la vertientes norte de la sierra nevada de Santa Marta, los cuales son sistemas de escorrentías pulsantes, que permiten un escurrimiento rápido del canal; estos eventos pueden no ser detectados con eficiencia debido a que los muestreos son realizados de forma puntual, tal como se diseño en este estudio.
En relación a la tasa de descomposición de la hojarasca se observaron diferencias según el gradiente altitunial del río Gaira, estas diferencias pueden estar asociadas a dos factores: uno mecánico definido por la velocidad de la corriente, proceso que es atribuible por la fragmentación física que tubo la hojasrasca expuesta en las mallas y el segundo puede deberse al uso que la fauna de macroinvertebrados hace sobre este sustrato que coloniza, porque utilizan este recurso vegetal para satisfacer varias necesidades ecológicas, entre ellas alimentación, sitio de refugio, construcción de capullos o galerías, o búsqueda de presas.
En cuanto al efecto de la condición nativa o exótica sobre tasa de descomposición, la hojarasca de especies nativas se descompone con más rapidez que las exóticas, aunque no se realizaron análisis sobre la composición química y estructural de las hojas, que permita definir una medida o escala de resistencia entre especies a la descomposición de su hojarasca, el resultado obtenido puede ser atribuible a procesos de co-evolución, donde las hojas de especies nativas son reconocidas como material nutricional y con estructuras vegetativas que pueden ser cortadas, fragmentadas e ingeridas con facilidad, generando por lo tanto una mayor demanda de macroinvertebrados sobre este recurso. El análisis discriminado de biomasa de los grupos funcionales asociados a la hojarascas de especies nativas o exóticas, muestra que los macroinvertebrados más grandes del grupo funcional CR se asocia preferencialmente a las especies exóticas, mientras que los ejemplares de mayor tamaño del grupo funcional CF se asocia con preferencia a la hojarasca de las especies nativas. Estos resultados permiten proponer este comportamiento como una de las causas que explica la mayor velocidad en la descomposición la hojarasca de origen nativo, al comparar su contribución en términos de biomasa.
AGRADECIMIENTOS
Los autores agraden a la división financiera de la Universidad Nacional de Colombia y a los grupos de Biodiversidad y Ecología Aplicada y Limnología Neotropical de la Universidad del Magdalena, al Grupo de Investigación Biodiversidad del Caribe Colombiano de la Universidad del Atlántico por su apoyo en el desarrollo exitoso del presente trabajo, tanto en la fase de campo como en la de laboratorio, ofreciendo sus instalaciones a lo largo de este estudio. Finalmente, a dos evaluadores anónimos por los comentarios y sugerencias que aportaron considerablemente a este manuscrito.
BIBLIOGRAFÍA
ALLAN JD. Stream Ecology, Estructure and Function of Running Waters. Edit. Chatman y Hall; 1995. [ Links ]
BENFIELD EF. Comparisons of litterfall input to streams. En Webster JR, Meyer JL (Eds.) Stream Organic Matter Budgets. J N Am Benthol Soc. 1997;16:3-161. [ Links ]
BENKE AC, HURYN AD, SMOCK LA, WALLACE JB. Length-mass relationships for freshwater macroinvertebrates in North America with particular reference the southeasten Unites States. J N Am Benthol Soc. 1999;18(3):308-343. [ Links ]
BRITTAIN JE, EIKELAND TJ. Invertevrate drift - A review. Hydrobiologia. 1988;166:77-93. [ Links ]
BOYERO L, BOSCH J. Spatial and temporal variation in macroinvertebrate drift in tropical streams. Biotropica. 2002;34:567-574. [ Links ]
CAMPBELL IC, JAMES KR, HART BT, DEVEREAUX A. Alloctonus coarse particulate organic material in forest and pasture reaches of two south-eastern Australian stream. II. Litter processing. Freshwater Biol. 1992;27:253-365. [ Links ]
CHARA AM, CHARA JD, ZUÑIGA M, PEDRAZA GX, GIRALDO LP. Clasificación trófica de insectos acuáticos en ocho quebradas protegidas de la ecorregión cafetera colombiana. Universidad Scientiarum. 2010;15:27-36. [ Links ]
CHESHIRE K, BOYERO L, PEARSON RG. Food webs in tropical Australian streams; shredders are not scarse. Freshwater Biol. 2010;50:748-764. [ Links ]
CRESSA C. Dry mass estimates of some tropical aquatic insects. Rev Biol Trop. 1999;47(1-2):133-141. [ Links ]
ELÓSEGUI A, SABATER S. Conceptos y técnicas en Ecología Fluvial. Madrid, Fundación BBVA. 2009;153-179. [ Links ]
FISHER SG, GE LIKENS. Energy flow in bear brook, New Hampshire: an integrative approach to stream ecosystem metabolism. Ecol Monogr. 1973;43:421-439. [ Links ]
FISHER SO, GRAY LJ. Secondary production and organic matter processing by collector macroinvertebrates in a desert stream. Ecology. 1983;64:1217-1224. [ Links ]
FRAYTER V, JIMENEZ E, PABON R, RIBERA V. Plan de manejo integral de la cuenca hidrográfica del río Gaira [tesis de pregrado]. Santa Marta: Programa de ingeniería agronómica. Universidad del Magdalena; 2000. [ Links ]
GONZÁLEZ M, GARCÍA D. Restauración de ríos y riberas, 2.ª ed. Madrid: Fundación Conde del Valle de Salazar y Mundi-Prensa; 1998. [ Links ]
GORDON N, MCMAHON T. FINLAYSON B. Stream hydrology an introduction for ecologists. John Wiley & Sons; 1992. [ Links ]
GRAÇA MAS, POZO J, CANHOTO C, ELOSEGI A. Effects of Eucalyptus plantations on detritus, decomposers, and detritivores in streams. The Scientific World Journal. 2002;2:1173-1185. [ Links ]
GREATHOUSE EA, PRINGLE CM. Does the river continuum concept apply on tropical island? Longitudinal variation in a Puerto Rican stream. Can J Fish Aquat Sci. 2005;53:896-909. [ Links ]
GRUBAUGH JW, WALLACE JB, HOUSTON ES. Longitudinal changes of macroinvertebrate communities along an Appalachian stream continuum. Can J Fish Acuat. Sci. 1996;53:896-909. [ Links ]
GUERRERO W, ZÚÑIGA F. Descomposición acuática de hojarasca de especies nativas y exóticas en la cuenca media baja del río Gaira, Magdalena [tesis de pregrado]. Santa Marta: Programa de Biología, Facultad de Ciencias Básicas, Universidad del Magdalena; 2005. [ Links ]
GUTIÉRREZ JD. Caracterización del metabolismo y de la oferta de recurso de materia orgánica para la fauna de macroinvertebrados béntonicos en una quebrada de montaña de orden menor [tesis de Doctorado]. Bogotá: Departamento de Biología, Universidad Nacional de Colombia; 2006. [ Links ]
HOFFMAN A. Dynamic of fine particulate organic mater (FPMO) and macroinvertebrates in natural and artificial leaf packs. Hidrobiologya. 2005;549:167-178. [ Links ]
IDEAM. Estudio Nacional del Agua 2010. Instituto de Hidrología, Meteorología y Estudios Ambientales. 2010. Bogotá D.C. [ Links ]
LEGENDRE P, LEGENDRE L. Numerical ecology. 2nd English edition. Amsterdam: Elsevier Science BV; 1998. [ Links ]
MERRIT RW, CUMMINS KW. An introduction to the acuatic insects of North America. Lowa: Kendall Hunt Publishing; 1996. [ Links ]
PETERSEN RC, CUMMINS KW. Leaf processing in a woodland stream. Freshwater Biol. 1974,4:343:368. [ Links ]
POZO J, GONZÁLES E, DÍAZ J, MOLINERO J, ELÓSEGUI A. Inputs of particulate organic matter to streams whit different riparian vegetation. J N Am Benthol Soc. 1997;16:602-611. [ Links ]
QUALLS RG, RICHARDSON CJ. Enrichment affects litter decomposition, immobilization, and soil microbial P in wetland mesocosms. Soil Sci Soc Am J. 2000;64:799-808. [ Links ]
R DEVELOPMENT CORE TEAM. R: A language and environment for statistical computing [2.11.1]. R foundation for statistical computing, Vienna, Austria. URL: http://www.R-project.org. [ Links ]
RAMÍREZ A, PRINGLE CM. Structure and production of a benthic insect assemblage in a neotropical stream. Rev Biol Trop. 1998;17:443-463. [ Links ]
RAMÍREZ A, PRINGLE CM. Spatial and temporal patterns of invertebrate drift in stream draining a Neotropical landscape. Freshwater Biol. 2001;46:47-62. [ Links ]
RIOS B. Comunidades de macroinvertebrados en un río altoandino [tesis de Doctorado]. Barcelona, España: Universidad de Barcelona; 2008. [ Links ]
RODRÍGUEZ JA. Dinámica de la materia orgánica en la cabecera de una quebrada de montaña (tesis de maestría). Bogotá: Departamento de Biología, Universidad Nacional de Colombia; 2006. [ Links ]
RODRIGUEZ-BARRIOS J, OSPINA-TORES R, GUTIERREZ JD, OVALLE H. Densidad y biomasa de macroinvertebrados acuáticos derivantes en una quebrada tropical de montaña (Bogotá, Colombia). Caldasia. 2007;29:397-412. [ Links ]
RUEDA-DELGADO G. Lineamientos de un programa de aseguramiento de la oferta hídrica del río Gaira a partir de la evaluación de la integridad biológica de la cuenca. DADMA. Universidad del Magdalena, Santa Marta, Colombia; 2005. [ Links ]
RUEDA-DELGADO G, WANTZEN KM, BELTRAN M. Leaf-litter descomposition in an Amazonian floodplain stream: effects of seasonal hydrological changes. J N Am Benthol Soc. 2006;25:233-249. [ Links ]
TAMARIS-TURIZO C, LOPEZ HJ. Aproximación a la zonificación climática de la cuenca del río Gaira. Intropica. 2006;3:69-76. [ Links ]
TOMANOVA S, GOTIA E, HELESIC J. Trophic levels and functional feeding group of macroinvertebrates in neotropical stream. Hidrobiología. 2006;556:251-264. [ Links ]
VANNOTE RL, MINSHALL GW, CUMMINS KW, SEDELL JR, CUSHING CE. The River Continuum Concept. Can J Fish Aquat Sci. 1980;37:130-137. [ Links ]
VYMAZAL J. Algae and element cycling in wetlands. Lewis publishers, Boca Raton Florida; 1995. [ Links ]
WANTZEN KM, WAGNER R, SUETFELD R, JUNK WJ. How do plant-herbivore interactions of trees influence coarse detritus processing by shredders in aquatic ecosystems of different latitudes?. Verh Int Verein Limnol. 2002;28:1-7. [ Links ]
WALLACE IB, WEBSTER JR. The role of macroinvertebrate in stream ecosystem function. Ann Rev Ecol Syst. 1996;17:567-594. [ Links ]
WEBSTER JR, BENFIELD EF. Vascular plant breakdown in freshwater ecosystem. Ann Rev Ecol Syst. 1986;17:567-594. [ Links ]
WINTERBOURN MJ. Fluxes of litter falling into a small beech forest stream. N Z J Mar Freshwater Res. 1976;10:399-416. [ Links ]

