










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. v.17 n.1 Bogotá jan./abr. 2012
EFECTOS DE LA SEPARACIÓN MATERNA TEMPRANA SOBRE EL DESEMPEÑO EN EL LABERINTO EN CRUZ ELEVADO EN RATAS ADULTAS
Effects of Early Maternal Separation on The Performance in the Elevated Plus Maze in Adult Rats
DIEGO ARMANDO LEÓN RODRÍGUEZ1, Estudiante Maestría en Neurociencias; ZULMA JANETH DUEÑAS GÓMEZ1, Ph. D.
1 Facultad de Medicina, Departamento de Ciencias Fisiológicas, Universidad Nacional de Colombia, Sede Bogotá. Correspondencia: zjduenasg@unal.edu.co
Presentado 19 de agosto de 2009, aceptado 28 de enero de 2010, correcciones 5 de marzo de 2012.
RESUMEN
Se ha demostrado que alteraciones en la interacción madre-cría produce efectos duraderos sobre el desarrollo cerebral y comportamental; de esta forma sujetos expuestos a estrés por separación materna temprana (SMT) presentan variaciones en los comportamientos indicadores de ansiedad. El objetivo del presente estudio fue investigar los efectos específicos del estrés por SMT sobre los comportamientos indicadores de ansiedad en ratas Wistar adultas machos y hembras. Las ratas fueron anidadas con ciclo luz/oscuridad invertido (7 p.m./7 a.m.), agua y comida ad libitum. El procedimiento de separación se realizó dos veces diarias durante los días postnatales 1 al 21 (7:00 a 10:00 y 13:00 a 16:00 p.m.). Los comportamientos indicadores de ansiedad fueron evaluados por medio del laberinto en cruz elevado (LCE) cuando las crías alcanzaron un peso de 230 g. Se encontró que el estrés por SMT tiene efectos específicos para cada sexo sobre los comportamientos relacionados con la ansiedad, las hembras separadas maternalmente presentaron un perfil ansioso menor que las no separadas y los machos separados mostraron mayor conflicto exploración/evitación. Estos resultados corroboran hallazgos preliminares de nuestro laboratorio, en los que se evidencia interacción entre la vulnerabilidad ante desafíos ambientales tempranos y los mecanismos compensatorios del cuidado materno.
Palabras clave: Separación materna, estrés, ansiedad, eje HPA, laberinto en cruz elevado.
ABSTRACT
It has been demonstrated that disruption of mother-pup interaction during early life exerts long-lasting effects on the brain and behavioral development. Therefore subjects exposed to early maternal separation stress (MS) show variations in anxiety-like behaviors. The aim of this study was to investigate the specific effects of SMT stress on anxiety-like behaviors in adult male and female Wistar rats. Rats were housed with reversed light dark cycle (light on at 7 p.m., off at 7 a.m.), water and food ad libitum. Separation was carried out in postnatal days 1 to 21, twice daily in dark cycle (7:00 a 10:00 y 13:00 a 16:00 p.m.). The anxiety-like behaviors were tested through the elevated plus maze (EPM) when the pups reached 230 g of weigh. We found that the MS stress has sex specific effects on anxiety-like behaviors: the maternal separated females displayed a lesser anxious outline than the not separated ones and the separated males showed a large exploration/avoidance conflict. These results confirm previous effects of our labs, which may be related to an interaction between vulnerability to environmental challenge and maternal care compensatory behaviors
Key words: Maternal separation, stress, anxiety, HPA axis, elevated plus maze.
INTRODUCCIÓN
En los mamíferos, incluyendo los seres humanos, las experiencias tempranas son de gran importancia para el desarrollo de circuitos cerebrales y patrones de comportamiento. Dentro de las experiencias más importantes se encuentra la interacción madre-cría; esta relación se presenta en la forma de vínculo afectivo (Bowlby, 1982; Bowlby, 1995; Harlow et al., 1966), el cual es propiciado por la actividad de sistemas neurodinámicos presentes en la cría y en la madre y es regulado a través de comportamientos maternos como las posturas de amamantamiento, acicalamiento y lamidas y de señales provenientes de la madre como olores, vocalizaciones y calor (Hoffer, 2006). Esta compleja interacción entre madre y cría podría tener por objeto brindar seguridad a los recién nacidos y modular la actividad de sus sistemas neuroendocrinos y fisiológicos que son básicos para el sostenimiento de la vida y que a largo plazo les permitirán desarrollar mecanismos de afrontamiento para sortear las futuras demandas ambientales (Macrì et al., 2008; Sullivan et al., 1986).
La relación entre madre e hijos en mamíferos suele ser flexible, hecho que le permitirá a la nueva descendencia ajustarse a la variabilidad ambiental, de forma tal que las características de esta relación, como la latencia e intensidad de los comportamientos maternos, funcionarían como predictor del ambiente futuro y contribuirán al moldeamiento de los sistemas endocrinos, neuronales y comportamentales (Macrì y Würbel, 2006). Esta flexibilidad en las experiencias tempranas es mayor en seres humanos, en los que los patrones de crianza dependen no solo de factores biológicos sino también culturales, sociales y cognoscitivos. A partir de la segunda mitad del siglo XX se generó gran interés por documentar los efectos de ambientes tempranos adversos sobre el funcionamiento adulto, tanto en humanos como en animales. En diferentes estudios se encontró que relaciones maternales pobres o negligentes se asocian con mayor vulnerabilidad para la aparición de disfunciones neurocomportamentales y un incremento en la susceptibilidad para adquirir enfermedades (Heim y Nemeroff, 2001). Además, existen evidencias epidemiológicas en las que se asocia estrés temprano con desarrollo de enfermedades mentales como trastornos de ansiedad, depresión, abuso de sustancias y trastornos de comportamiento (Gilmer y McKinney, 2003).
LA SEPARACIÓN MATERNA TEMPRANA
La relación materna y sus efectos sobre el desarrollo ha sido estudiada extensamente mediante el uso de protocolos animales de manipulación postnatal. En dichos procedimientos se altera el tiempo de permanencia de las crías con sus madres y se evalúa cómo esta separación afecta los desempeños en pruebas de comportamiento y funcionamiento neuroendocrino. Se han explorado diferentes protocolos de separación, en los que se varía tiempo de separación de las crías de la madre (desde unos pocos minutos hasta 24 horas), número de días del procedimiento y momento del desarrollo en que se encuentran los recién nacidos. Dentro de estos protocolos, los más usados son los de separación materna temprana (SMT) y los de manipulación temprana (MT), los cuales difieren en el tiempo de retiro de las crías desde su nido; estas diferencias han llevado a resultados diferentes y muchas veces opuestos. En general se ha encontrado consistentemente que la MT causa disminución en la intensidad de la respuesta emocional ante estresores, mientras que los resultados sobre los efectos de la SMT obte nidos en diversos estudios, no son sólidos; en algunos no se observan diferencias con la no separación, en otros se observa decremento en la intensidad de la respuesta emocional y en otros los indicadores de ansiedad aumentan. Autores como Levine, 2005, y Macrí y Würbel, 2006, han propuesto que las diferencias se relacionan con la interacción compleja entre los comportamientos compensatorios de la madre y el tiempo de exposición al estrés fisiológico (para una revisión más detallada remitirse a Macrí y Würbel, 2006) el cual depende directamente del tiempo y número de separaciones diarias. Este trabajo está enfocado a evaluar las consecuencias que trae para ratas adultas, el haber sido sometido a estrés por SMT, sobre los comportamientos relacio-nados con las respuestas de ansiedad.
Con respecto a las respuestas de ansiedad, estudios de Caldji et al., 2000a, Caldji et al., 2000b, Francis y Meaney, 1999, Huot et al., 2001, Huot et al., 2002, Huot et al., 2004, Liu et al., 2000a y Liu et al., 2000b, han reportado que el uso de un protocolo de 4 horas diarias de separación por 14 días es suficiente para generar un incremento en respuestas de temor condicionado e incondicionado e hiperactividad del eje HPA en las respuesta a estresores (Marmendal, 2005). Sin embargo, otros estudios, siguiendo procedimientos similares, no han encontrado tales alteraciones (Shalev y Kafkafi, 2002). En hallazgos previos en nuestro laboratorio usando un protocolo de seis horas diarias, divididas en 2 periodos de separación, durante 21 días, hemos encontrado que los efectos de SMT son diferenciales para cada uno de los sexos, las hembras separadas maternalmente exhiben reducción en comportamientos relacionados con ansiedad (Moreno et al., 2009). Estos hallazgos iniciales pueden orientarnos hacia la consolidación de una teoría más consistente de la separación materna. Para la realización de este estudio y con el fin de validar los resultados previos obtenidos por el grupo, se amplió el número de sujetos control y experimentales, se hizo un registro detallado de comportamientos indicadores de ansiedad y se incluyeron otras variables etológicas que brindan mayor información sobre la forma como el estrés temprano puede estar afectando de forma específica las respuestas incondicionadas de ansiedad de machos y hembras.
MEDICIÓN DE LAS RESPUESTAS INCONDICIONADAS DE ANSIEDAD
Dentro de los instrumentos para medir variaciones en los comportamientos relacionados con ansiedad, el laberinto en cruz elevado (LCE) es uno de los que cuenta con una mejor validez etológica. Este laberinto tiene como fundamento los comportamientos de evitación incondicionada que presentan las ratas hacia espacios abiertos y elevados. Pellow et al., 1985, realizaron estudios en los que encontraron que el paso de un animal por los brazos abiertos conlleva a incremento en los niveles plasmáticos de corticoesterona, en las respuestas de congelamiento y mayor producción de bolos fecales; estos cambios fueron tomados como indicadores de un incremento en la ansiedad de los animales ante la entrada a brazos abiertos. En la exploración natural del laberinto, hay tendencia clara en los animales por permanecer mayor tiempo dentro de los brazos cerrados. El LCE ha demostrado ser sensible a los efectos de fármacos ansiolíticos y ansiogénicos: animales a los que se les administró benzodiacepinas pasaron mayor tiempo en los brazos abiertos y disminuyeron las respuestas de congelamiento y niveles de corticoesterona. De esta forma, el LCE se ha convertido en un instrumento eficaz para evaluar de forma rápida, económica y confiable las respuestas incondicionadas de ansiedad en ratas. Adicionalmente a las medidas clásicas como número de entradas y tiempo de permanencia en los brazos, el LCE permite analizar comportamientos naturales de las ratas como estiramientos, acicalamientos, sacar la cabeza del laberinto y pararse en las patas traseras, que permiten hacer análisis más profundos con respecto a las respuestas ansiosas.
A pesar de la utilidad del LCE, aún existen pocos estudios en SMT que lo usen para evaluar sus efectos sobre ansiedad (Estanislau y Morato, 2005), dentro de estos trabajos (Kalinichev et al., 2002; McIntosh et al., 1999), se encuentran reportes poco consistentes y contradictorios, en los que solamente se usan las medidas tradicionales. Otro elemento que suele ser subestimado en los estudios sobre SMT es la influencia del sexo, es frecuente que solamente se evalúen machos para evitar problemas de la variabilidad comportamental asociada con cambios hormonales en las etapas del ciclo estral. Adicionalmente, existen diferencias evidentes en los perfiles de respuesta ante estímulos que generan ansiedad entre machos y hembras. Estas diferencias no son aprendidas y se encuentran asociadas con mecanismos neurobiológicos y endocrinos innatos propios de cada sexo, involucrados en la modulación de estrés y la respuesta ante amenazas (Blanchard et al., 1995), de forma tal, que ratas hembra suelen mostrar mayores niveles de ACTH y corticosterona ante estresores, así como mayor actividad en pruebas como LCE y campo abierto (Ray y Hansen, 2004). Con respecto a los estudios de SMT y sus consecuencias y su consecuencia por género, se ha reportado la existencia de efectos específicos tanto para machos como para hembras, pero estos efectos no son consistentes dentro de los pocos estudios que los han explorado (Díaz Luján et al., 2008). En nuestro laboratorio hemos encontrado que la valoración de ciertas medidas etológicas complementarias dentro del LCE pueden llegar a ser de gran utilidad para enriquecer la discusión acerca de los efectos diferenciales de la SMT sobre las respuestas de ansiedad en ratas machos y hembras (Moreno et al., 2009); sin embargo, se debe tener en cuenta que el análisis de los efectos de la SMT debe ser realizado dentro y no entre cada uno de los géneros con el fin de evitar que las diferencias neuroendocrinas que median las respuestas entre cada uno de los grupos, interfiera con los efectos del estrés por SMT. En ese sentido la rigurosidad de la evaluación de las etapas del ciclo estral en la cual se encuentran las hembras, es fundamental. El objetivo de este trabajo fue evaluar los comportamientos relacionados con la ansiedad, medidos a través del laberinto en cruz elevado, en ratas Wistar adultas que han sido sometidas a SMT frente a aquellas que no lo fueron y determinar posibles deferencias en estos efectos dentro de cada uno de los grupos.
MATERIALES Y MÉTODOS
SUJETOS
Se utilizaron ratas albinas de la cepa Wistar de 100 días obtenidas del bioterio de producción de la Facultad de Medicina Veterinaria de la Universidad Nacional de Colombia, sede Bogotá. Las ratas fueron anidadas de forma individual por un periodo de dos semanas para permitir su adaptación a las condiciones de temperatura (± 22 °C), humedad y ciclo luz/oscuridad (12/12), este ciclo fue invertido comenzando con oscuridad a las 7:00 a.m. y luz a las 7:00 p.m. Se controlaron estímulos ambientales como olores y ruidos. Todos los animales tuvieron acceso a agua y alimento ad-libitum.
Una vez cumplido el tiempo de adaptación las ratas parentales se anidaron por parejas de hembras y machos durante 21 días, cumplido este tiempo se retiraron los machos de los nidos y no se manipularon las hembras hasta el alumbramiento. El día de alumbramiento se sexaron las crías y se asignaron de forma semialeatoria a los grupos separación y control, tratando de obtener un número similar de crías en cada uno de los grupos. Los sujetos controles, fueron las crías de la misma camada que no fueron sometidas al protocolo de SMT, pero que sí recibieron manipulaciones periódicas relacionadas con la limpieza de las cajas. El número total de sujetos usados en las pruebas de comportamiento fue de 48 ratas, distribuidas de la siguiente forma: hembras separadas 11, hembras control 10, machos separados 11 y machos control 16. Todos los procedimientos durante esta investigación se realizaron de conformidad con las normas éticas y legales exigidas para la investigación con animales de laboratorio en Colombia (Ley 84 de 1989 y Resolución No. 8430 de 1993 del Ministerio de Salud); al igual se tuvieron en cuenta las recomendaciones para el manejo y cuidado de animales de laboratorio propuestos por la normatividad de la Unión Europea (8616091EU) y del Instituto Nacional de Salud de los Estados Unidos de América (National Research Council, 1996).
DISEÑO EXPERIMENTAL
Estudio experimental no invasivo de casos y controles. El grupo experimental consiste en sujetos sometidos a estrés postnatal perinatal mediante un procedimiento de separación materna temprana. El grupo control corresponde a sujetos que no han sido sometidos a estrés postnatal por separación materna temprana.
PROCEDIMIENTO DE SEPARACIÓN MATERNA TEMPRANA
El día del nacimiento de las crías se contó como día cero. A partir del día postnatal 1 hasta el 21 se aplicó un protocolo de separación materna temprana de 360 minutos, el cual consistía en dos periodos de separación de 180 minutos cada uno (7:00 a 10:00 y 13:00 a 17:00); se dispuso de un tiempo entre separaciones de 180 minutos, en el que las crías eran retornadas a sus respectivos nidos para ser alimentadas y así evitar efectos causados por desnutrición. Para la separación materna, primero se removió la madre del nido a una caja separada, luego las crías se ubicaron en otra caja plástica que contenía una cama de aserrín previamente calentada, después las nodrizas se ubicaron de nuevo en el nido junto con las crías no separadas. Las cajas con ratas separadas fueron llevadas a otra habitación en la que se mantuvieron las condiciones de temperatura, humedad y control de ruidos. Pasados los 180 minutos de separación, se retornaron las ratas separadas a la habitación original, se retiró la madre del nido, se devolvieron las crías separadas a la caja nido y se colocó de nuevo la madre junto con todas sus crías. A partir del día 21 (destete), se suspendió el procedimiento de separación y se anidaron las ratas en cajas por grupos de sexo y tratamiento, donde crecieron en condiciones similares, con libre acceso a agua y comida, hasta que tuvieron una edad adulta para realizar las pruebas de comportamiento.
PRUEBA LABERINTO EN CRUZ ELEVADO
Las respuestas incondicionadas de ansiedad fueron evaluadas a través del LEC cuando los animales alcanzaron un peso superior a 230 g, momento en el que se consideraron adultos jóvenes. En promedio los machos obtuvieron este peso hacia los 55 días y las hembras hacia los 70 días. El LCE es en una plataforma acrílica de color negro en forma de cruz que se encuentra elevada a 50 cm del suelo y está compuesta por dos brazos abiertos de 50 x 10 cm y dos cerrados de 50 x 10 con paredes de 40 cm de altura y descubierta en la parte superior. En cada sesión experimental, los animales fueron puestos de manera individual en el centro del laberinto, enfrentando una rama abierta, con la luz encendida. El animal era retirado del laberinto después de 5 minutos de exploración libre. La sesión se realizó en la tarde (14:00 a 17:00) porque en este periodo los niveles de corticosterona son más bajos. Las hembras fueron evaluadas solamente cuando se encontraban en la fase de diestro de su ciclo estral, porque existe evidencia que en este periodo los comportamientos de las hembras son más homogéneos con el de los machos (Gouveia et al., 2004). La determinación de la etapa del ciclo en la cual se encontraban las hembras, se realizó por medio de citología funcional 4 horas antes de la sesión en el LCE, de tal forma que para el momento de la prueba, los niveles de estrés producido por el ciclaje, han retornado a su estado basal. Para la citología, se toma la rata por la cola, apoyando la palma de la mano izquierda sobre el dorso del animal, se sujeta y se levanta la cola y con un gotero que tiene solución salina, se introduce en la vagina, se succiona suavemente, se saca el gotero y se deposita su contenido en un portaobjetos. De inmediato se observa al microscopio para identificar el tipo celular y así determinar en que etapa del ciclo esta la rata. En la habitación solamente se dispuso el LCE, el recinto fue iluminado con luz artificial blanca y se aisló acústicamente. Los comportamientos de los sujetos fueron grabados por medio de una cámara de video ubicada en el techo de la habitación.
La codificación de los comportamientos se realizó por medio del programa X-plo Rat 2005, con el que se registraron los siguientes patrones comportamentales:
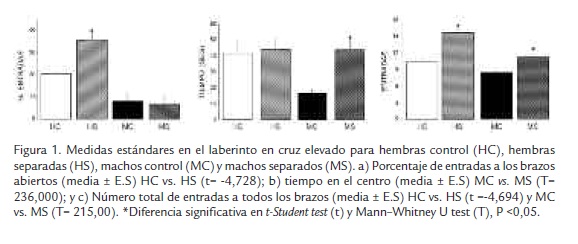
Medidas estándares: a) Frecuencia de entradas a brazos abiertos y cerrados: se considera que un animal entra a un brazo cuando pisa con las cuatro paras dicho brazo, a mayor número de entradas a los brazos abiertos, menor ansiedad; b) entradas totales a los brazos: una mayor cantidad de entradas a estos brazos se relaciona con mayor actividad general; c) tiempo gastado en cada uno de los brazos: mayor cantidad de tiempo en brazos abiertos indican menor ansiedad. Con estos datos se calculó el porcentaje de entradas a brazos; porcentaje de tiempo en los brazos abiertos, cerrados y el centro.
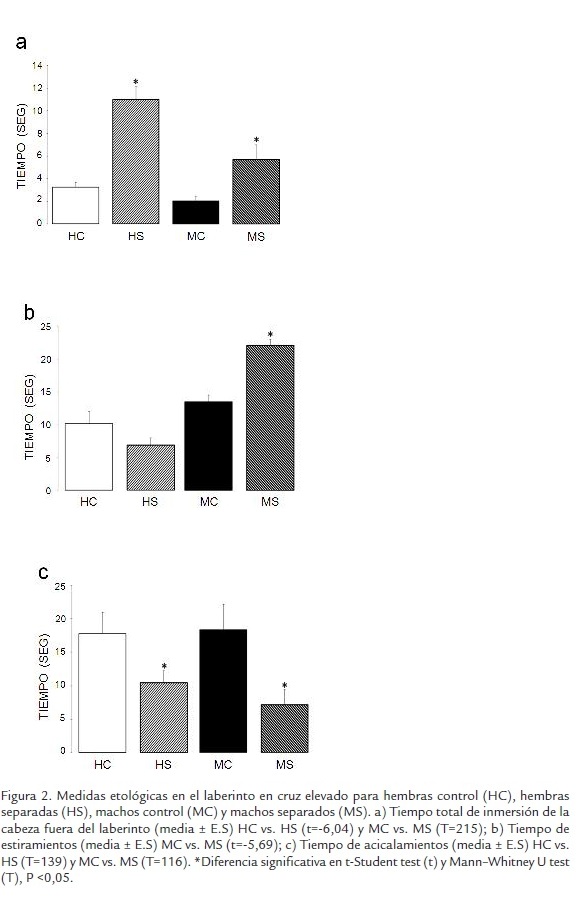
Medidas etológicas: a) Frecuencia y duración de estiramientos: se considera que ha ocurrido un estiramiento cuando el animal extiende completamente su cuerpo para explorar un área del laberinto sin moverse hacia este espacio; b) Frecuencia y duración de levantamientos: se considera que ha ocurrido un levantamiento cuando la rata se alza sobre sus patas traseras en posición vertical para explorar el espacio, no existe una clara relación de esta conducta con la ansiedad, se ha propuesto una asociación con hiperactividad; c) Hundimiento o Inmersión de la cabeza: se codifica como hundimiento de la cabeza cada vez que el sujeto explora por fuera del laberinto sumergiendo su cabeza en el vacío, este es un comportamiento exploratorio del espacio externo del laberinto que se ha asociado estrechamente con baja ansiedad y d) Acicalamientos: se cataloga como acicalamiento cada vez que el animal sacude sus patas sobre su cuerpo como si se estuviera limpiándose, no se ha reportado una relación directa de este comportamiento con los comportamientos de ansiedad. Una vez finalizadas las observaciones, se sacrificaron los animales, se extrajeron sus cerebros y se recolectó su sangre para estudios posteriores.
ANÁLISIS DE LOS DATOS
Para el análisis estadístico de los datos se realizaron comparaciones entre las medias de los grupos controles y separados con la prueba t-Student, si las muestras no cumplían con los criterios de normalidad u homogeneidad se aplicó la prueba no paramétrica U de Mann-Whitney. Estos análisis fueron realizados con el programa estadístico Sigma Stat 3.5. No se realizaron análisis de varianza ANDEVA porque nuestro interés no es observar interacción entre los factores género y la SMT, un análisis en conjunto de estos dos factores distorsionaría los efectos de la SMT, como se mencionó anteriormente, existen diferencias entre los géneros en las repuestas de ansiedad que no son debidas a las experiencias tempranas.
RESULTADOS MEDIDAS ESTÁNDARES
Como se muestra en la Figura 1A, las hembras separadas presentaron mayor porcentaje de entradas a los brazos abiertos, mientras que en los machos no se observaron diferencias significativas, lo cual indicaría menor ansiedad en las hembras separadas maternalmente, con respecto a las hembras no separadas. Para el caso de los machos, se encontró aumento significativo en el tiempo de permanencia en el centro en el grupo separado maternalmente en comparación con los machos controles (Figura 1B), esta tendencia indica leve reducción de ansiedad pero no lo suficiente como para permitirle explorar los brazos abiertos. Tanto en machos como en hembras separadas se observó que incrementaron las entradas a los dos brazos (Figura 1C), lo cual indica una mayor hiperactividad en el grupo separación con respecto a los controles.
MEDIDAS ETOLÓGICAS
Inmersión de la cabeza: tal como se observa en la Figura 2A, tanto machos como hembras separadas maternalmente mostraron incremento en el tiempo de inmersiones de la cabeza por fuera del laberinto, comparado con los sujetos control de cada uno de sus grupos, lo que indica que los grupos separados, en especial las hembras presentan menor ansiedad. Estiramientos del cuerpo: tal como se muestra en la Figura 2B, solamente se presentaron diferencias significativas entre los machos, el grupo separación ocupó mayor tiempo en estiramientos del cuerpo que el grupo control, esta tendencia es un indicador de un conflicto entre exploración y evitación. Acicalamientos: hubo disminución significativa del tiempo de acicalamientos en los grupos separados tanto de machos como de hembras con respecto a sus respectivos controles (Figura 2C), este puede estar relacionado con mayor tiempo invertido en la exploración del laberinto. Levantamientos: no se encontraron diferencias significativas entre los grupos separados y no separados.
DISCUSIÓN Y CONCLUSIONES
Los resultados encontrados en este estudio apoyan hallazgos preliminares de nuestro laboratorio, en los que se ha visto que el estrés crónico por separación materna, durante la lactancia, conlleva a alteraciones en los comportamientos dentro del laberinto en cruz elevado (Moreno et al., 2009), lo cual corrobora la confiabilidad del uso del protocolo de SMT por 6 horas diarias divididas en dos periodos de tres horas, durante los 21 días de amamantamiento y sus efectos consistentes sobre los comportamientos relacionados con la ansiedad. Las alteraciones comportamentales se presentan tanto en machos como en hembras, pero la forma como se ven modificados los perfiles de comportamientos relacionados con ansiedad es específica para cada uno de los sexos. En un primer acercamiento a los efectos de la separación materna sobre el desempeño de las ratas en el LCE se puede concluir que existe una disminución marcada en las conductas indicadoras de ansiedad en ratas hembras y que este efecto de ansiolisis es menor en los machos, resultados concordantes con lo encontrado en los estudios de Eklund y Arborelius, 2006, y Slotten et al., 2006, quienes usaron un protocolo de SMT similar al nuestro. No obstante, los resultados en el estudio de Eklund y Arborelius, 2006, no arrojaron diferencias marcadas en entre machos, probablemente porque el instrumento de medida no les permitió hacer una observación fina de los comportamientos de los animales; el uso de las mediciones etológicas realizadas en esta investigación, nos permitió hacer un análisis más profundo de los efectos de SMT sobre el comportamiento de ratas adultas y detectar efectos específicos de este estrés sobre el comportamiento de machos y hembras.
Los comportamientos que realizan los roedores dentro del LCE se pueden agrupar en tres factores: ansiedad, actividad locomotora y evaluación del riesgo (Rogers y Johnson, 1995). El factor ansiedad básicamente se compone del porcentaje y el tiempo de entradas a los brazos abiertos y del tiempo y frecuencia de inmersiones de la cabeza por fuera del laberinto, al observar estos indicadores, podemos decir que las ratas hembra que fueron separadas maternalmente presentan una clara disminución en su perfil ansioso con respecto a ratas hembras no separadas, en el caso de los machos esta reducción de la ansiedad no es tan evidente, pero se refleja en el leve incremento en el tiempo y frecuencia de inmersiones de la cabeza por fuera del laberinto. La actividad locomotora se vería reflejada en el número de entradas totales y de entradas a brazos cerrados, estos dos indicadores se incrementaron tanto en machos como en hembras separadas, resultados que concuerdan con hallazgos previos que indican que la separación materna incrementa niveles de actividad locomotora (Marin y Planeta, 2004), cabe resaltar que estos incrementos en la actividad locomotora en roedores tienden a correlacionar positivamente con perfiles ansiolíticos (Rogers y Johnson, 1995). Adicionalmente, diferentes estudios han arrojado evidencias que demostrarían que los incrementos en actividad locomotora y ansiolisis aumentan la sensibilidad a los efectos de psicoestimulantes como la cocaína (Kikusui et al., 2005; Planeta y Marin, 2002), estos datos en su conjunto pueden ser tomados en cuenta para la construcción de un modelo para estudiar la influencia de las experiencias tempranas sobre la aparición de trastornos del desarrollo como el déficit de atención e hiperactividad (Kwak et al., 2009), problemas de control de impulsos y vulnerabilidad al consumo de estimulantes (Marin y Planeta, 2004).
El tercer factor que surge en los análisis del LCE está compuesto por el tiempo de permanencia en el centro del laberinto y la frecuencia y duración de estiramientos, incrementos en estas tendencias conductuales podrían reflejar procesos de valoración del riesgo y un conflicto entre la evitación de potenciales amenazas y la exploración de espacios nuevos (Rogers y Johnson, 1995). Adicionalmente, se ha reportado que los acicalamientos y el tiempo de permanencia en los brazos cerrados se relacionan negativamente con estos dos comportamientos; se infiere que los animales que presentan altos niveles de acicalamiento o permanecen más tiempo en los brazos cerrados ya han tomado una decisión comportamental y por lo tanto no presentan conflicto evitación /exploración. De esta forma, los machos separados mostraron claramente un conflicto evitación/exploración, determinado por pasar mayor tiempo en el centro (Fig. 1a) y presentar mayor número y tiempo de estiramientos del cuerpo (Fig. 2b), por el contrario, los machos y hembras control pasaron mayor tiempo en los brazos cerrados, mostraron pocos estiramientos y gran número de acicalamientos (Fig. 2c), lo que indicaría que las hembras y machos control tienden a tomar una decisión evitativa, las hembras separadas, una exploratoria, y los machos control presentarían mayor conflicto para tomar cualquier decisión.
Las diferentes medidas etológicas, permiten hacer un análisis con mayores matices y aproximarnos de forma más exhaustiva a la complejidad de los comportamientos incondicionados de roedores. Los roedores así como la mayor parte de mamíferos, poseen sistemas afectivos que influyen de forma dinámica y compleja en su comportamiento (Panksepp, 1998), en esta medida, una aproximación a los perfiles de ansiedad debe contemplar esta multideterminación, en la que los animales no solamente actúan por evitación sino que también hay en marcha sistemas afectivos competitivos como la exploración para búsqueda de recursos.
Como se mostró en los resultados, los efectos de la separación materna temprana son específicos para cada uno de los géneros, por ello sería pertinente tener en cuenta estos efectos diferenciales de las experiencias tempranas al momento de realizar estudios sobre desarrollo, en especial en modelos de ansiedad. El manejo no adecuado de diferencias entre sexos, ya sea por inclusión de solo uno de los géneros o por no realizar análisis para cada uno de ellos puede suscitar distorsiones en los resultados, este puede ser un factor muy importante para que en algunos estudios se encuentren efectos opuestos, por ejemplo el estudio de Daniels et al., 2003, en el que la SMT incrementó conductas de evitación, pero no se realizó un análisis de las diferencias dentro de cada uno de los géneros. A pesar que las diferencias intergénero en la modulación de las amenazas puedan ser un obstáculo, también es una gran oportunidad para indagar sobre cuáles son los mecanismos responsables de estas diferencias y así comprender más cercanamente la neurodinámica implicada en el desarrollo de conductas patológicas como los trastornos de ansiedad, déficit atencional, problemas en el control de impulsos, depresión, abuso de sustancias, entre otras. Probablemente el estrés temprano altere la fisiología de estructuras como el área preóptica, el núcleo terminal estriado y el núcleo del tracto solitario, estructuras que presentan una neurodinámica específica para cada sexo y que son importantes para la regulación del eje HPA en el cual se modulan respuesta neuroendocrinas ante estímulos amenazantes (Ulrich Lai y Herman, 2009), A su vez, es probable que también existan modificaciones en estructuras como el hipocampo, la amígdala y la corteza prefrontal, que aunque no presentan un dimorfismo sexual tan marcado si participan en la modulación del eje HPA (Caldji et al., 2000b).
Los mecanismos por medio de los cuales el estrés por SMT induce estas diferencias comportamentales aún no son claros. El estrés que se produce por la separación no es la única fuente de cambios durante el desarrollo; se ha reportado que las ratas madres cambian su repertorio comportamental de cuidado materno ante la separación crónica de sus crías, lo cual puede estar relacionado con modificaciones endocrinas en la rata madre (Macrì y Würbel, 2006). La interpretación de la influencia de estrés temprano resulta más compleja, porque durante los primeros días de desarrollo, la regulación del eje HPA no se realiza a través de liberación de corticosterona, dado que su liberación es mínima durante este tiempo (Levine, 1967), por el contrario es realizada a través de arginin vasopresina (AVP; Makara et al., 2008). Es posible que los efectos de SMT sobre el sistema nervioso central y sistema endocrino estén mediados por la liberación de AVP, lo cual probablemente induciría modificaciones funcionales en el adulto sobre sistemas de neurotransmisores como el GABA, en especial en la expresión de receptores para este fármaco (Caldji et al., 2000b).
El uso de SMT en roedores como un modelo de las consecuencias que traen las experiencias adversas tempranas sobre el comportamiento adulto, tiene gran relevancia para la indagación de los factores involucrados en el desarrollo de pautas comportamentales específicas; además ha permitido hacer una amplia exploración de la forma como ciertos sistemas cerebrales y endocrinos dependen de experiencias tempranas, dentro de las que el papel de la madre como agente regulador de desarrollo de los sistemas neuroendocrinos es vital (Hoffer, 2006). Este modelo de SMT ha permitido esbozar nuevas teorías sobre el desarrollo de una mayor o menor vulnerabilidad para la adquisición de patologías mentales y comportamentales como abuso de sustancias, depresión, trastornos de ansiedad, problemas de aprendizaje, deficiencias atencionales e hiperactividad, no obstante, debemos ser cautelosos al hacer inferencias desde modelos en animales a humanos porque las dinámicas de desarrollo neurocomportamentales son diferentes entre especies. Adicionalmente, los determinantes del comportamiento humano son mucho más complejos y una aproximación a la patología mental humana solo puede ser hecha teniendo en cuenta la diversidad de interacciones entre la genética de nuestra especie y ambiente en el que esta se expresa.
AGRADECIMIENTOS
Queremos agradecer especialmente a Diego Andrés Sierra, estudiante de Medicina de la Universidad Nacional de Colombia, quien durante su internado estuvo colaborando con el cuidado y mantenimiento de los sujetos experimentales. Este trabajo fue financiado por la Facultad de Medicina de la Universidad Nacional de Colombia, sede Bogotá, Quipú: 203010011339 (8518), 2008.
BIBLIOGRAFÍA
BLANCHARD DC, GRIEBEL G, BLANCHARD RJ. Gender bias in the preclinical psychopharmacology of anxiety: Male models for (predominantly) female disorders. J Psychopharmacol. 1995;9:79-82. [ Links ]
BOWLBY J. Attachment. 2 ed. New York: Basic Books; 1982. [ Links ]
BOWLBY J. Maternal Care and Mental Health, 2ª ed. edición, Northvale, NJ; London: Jason Aronson. Geneva, World Health Organization, Monograph series no. 3; 1995. [ Links ]
CALDJI C, DIORIO J, MEANEY MJ. Variations in maternal care in infancy regulate the development of stress reactivity. Biol Psychiat. 2000a;48:1164-1174. [ Links ]
CALDJI C, FRANCIS D, SHARMA S, PLOTSKY PM, MEANEY MJ. The effects of early rearing environment on the development of GABAA and central benzodiazepine receptor levels and noveltyinduced fearfulness in the rat. Neuropsychopharmacol. 2000b;22:219-229. [ Links ]
DANIELS WM, PIETERSEN CY, CARSTENS ME, STEIN DJ. Maternal separation in rats leads to anxiety-like behavior and a blunted ACTH response and altered neurotransmitter levels in response to a subsequent stressor. Met Brain Dis. 2004;19:3-14. [ Links ]
DÍAZ LUJÁN VG, CASTELLANOS NN, LEVIN G, SUAREZ MM. Amitriptyline: sexdependent effect on sympathetic response and anxiety in rats submitted to early maternal separation and variable chronic stress in adulthood. Int J Dev Neurosci. 2006;26:415-422. [ Links ]
EKLUND M, ARBORELIUS L. Twice daily long maternal separations in Wistar rats decreases anxiety-like behaviour in females but does not affect males Behav Brain Res. 2006;172:278-285. [ Links ]
ESTANISLAU C, MORATO S. Prenatal stress produces more behavioral alterations than maternal separation in the elevated plus-maze and in the elevated Tmaze. Behav Brain Res. 2005;163(1):70-77. [ Links ]
FRANCIS DD, MEANEY MJ. Maternal care and the development of stress responses. Curr Opin Neurobiol. 1999;9(1):128-134. [ Links ]
GILMER W, MCKINNEY W. Early Experience and depressive disorders: Human and non-human primate studies. J Affective Disorders. 2003;75(2):97-113. [ Links ]
GOUVEIA A, DOS SANTOS U, FELISBINO F, DE AFONSECA T, ANTUNES G, MORATO S. Influence of the estrous cycle on the behavior of rats in the elevated Tmaze. Behav Process. 2004;67:167-171. [ Links ]
HARLOW HF, HARLOW MK, DODSWORTH RO, ARLING GL. Maternal Behavior of Rhesus Monkeys Deprived of Mothering and Peer Associations in Infancy. P Am Philos Soc. 1966;110(1):58-66. [ Links ]
HEIM C, NEMEROFF C. The role of childhood trauma in the neurobiology of Mood and Anxiety disorders: Preclinical and clinical studies. Biol Psychiat. 2001;49 (12):1023-1039. [ Links ]
HOFFER M. Psychobiological Roots of Early Attachment. Curr Dir Psychol Sci. 2006;15(2):84-88. [ Links ]
HUOT R L, THRIVIKRAMAN KV, MEANEY MJ, PLOTSKY PM. Development of adult ethanol preference and anxiety as a consequence of neonatal maternal separation in Long Evans rats and reversal with antidepressant treatment. Psychopharmacol. 2001;158(4):366-373. [ Links ]
HUOT RL, PLOTSKY PM, LENOX RH, MCNAMARA RK. Neonatal maternal separation reduces hippocampal mossy fiber density in adult Long Evans rats. Brain Res. 2002;50:52-63. [ Links ]
HUOT RL, GONZALEZ ME, LADD CO, THRIVIKRAMAN KV, PLOTSKY PM. Foster litters prevent hypothalamic-pituitary-adrenal axis sensitization mediated by neonatal maternal separation. Psychoneuroendocrino. 2004;29(2):279-289. [ Links ]
KALINICHEVA M, EASTERLINGA K, PLOTSKYB M, HOLTZM S. Long-lasting changes in stress-induced corticosterone response and anxiety-like behaviors as a consequence of neonatal maternal separation in Long-Evans rats. Pharmacol Biochem Be. 2002;73:131-140. [ Links ]
KIKUSUI T, FACCIDOMO S, MICZEK K. Repeated maternal separation: differences in cocaine-induced behavioral sensitization in adult male and female mice. Psychopharmacol. 2005;178:202-210. [ Links ]
KWAK H, LEE J, KWON K, KANG C, CHEONG Y, CHUN W, KIM S, LEE H. Maternal Social Separation of Adolescent Rats Induces Hyperactivity and Anxiolytic Behavior. The Korean Society of Pharmacology. 2009;13:79-83. [ Links ]
LEVINE S. Maternal and environmental influences on the adrenocortical response to stress in weanling rats. Science. 1967;156:258-260. [ Links ]
LEVINE S. Developmental determinants of sensitivity and resistance to stress. Psychoneuroendocrino. 2005;30:939-946. [ Links ]
LIU D, DIORIO J, TANNENBAUM B, CALDJI C, FRANCIS D, et al. Maternal care, hippocampal glucocorticoidreceptors, and hypothalamic-pituitary-adrenal responses to stress. Science. 1997;277(5332):1659-1662. [ Links ]
LIU D, CALDJI C, SHARMA S, PLOTSKY P M, MEANEY MJ. Influence of neonatal rearing conditions on stress-induced adrenocorticotropin responses and norepinepherine release in the hypothalamic paraventricular nucleus. J Neuroendocrinol. 2000a;12(1):5-12. [ Links ]
LIU D, DIORIO J, DAY JC, FRANCIS DD, MEANEY MJ. Maternal care, hippocampal synaptogenesis and cognitive development in rats. Nature Neuroscience. 2000b;3(8):799-806. [ Links ]
MACRÍ S, WÜRBEL H. Developmental plasticity of HPA and fear responses in rats: a critical review of the maternal mediation hypothesis. Horm Behav. 2006;50:667-80. [ Links ]
MACRÍ S, CHIAROTTI F, WÜRBEL H. Maternal separation and maternal care act independently on the development of HPA responses in male rats. Behav Brain Res. 2008;191:227-234. [ Links ]
MAKARA GB, DOMOKOS A, MERGL Z, CSABAI K, BARNA I, ZELENA D. Gender-specific regulation of the hypothalamo-pituitary-adrenal axis and the role of vasopressin during the neonatal period. Stress, Neurotransmitters, and Hormones: Ann NY Acad Sci. 2008;1148:439-445. [ Links ]
MARIN M, PLANETA C. Maternal separation affects cocaine-induced locomotion and response to novelty in adolescent, but not in adult rats. Brain Res. 2004. p. 83-90. [ Links ]
MARMENDAL M. Maternal Separation in the Rat: Long-term Effects of Early Life Events on Emotionality, Drug Response and Neurobiology [Doctoral dissertation]. Department of Psychology, Göteborg University, Sweden; 2005. [ Links ]
MCINTOSH J, ANISMAN H, MERALI Z. Shortand long-periods of neonatal maternal separation differentially affect anxiety and feeding in adult rats, gender dependent effects. Brain Res Dev Brain Res. 1999;113:97-106. [ Links ]
MORENO L, LAMPREA M, DUEÑAS Z. Diferencias en los comportamientos asociados con La ansiedad de ratas macho y hembra expuestas a un protocolo de estrés crónico por separación Maternal temprana. Suma Psicológica. 2009;16(1):31-43. [ Links ]
PANKSEPP J. Affective Neuroscience. New York: Oxford University Press; 1998. [ Links ]
PELLOW S, CHOPIN P, FILE SE, BRILEY M. Validation of open: closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci Meth. 1985;14:149-167. [ Links ]
PLANETA S, MARIN M. Effect of cocaine on periadolescent rats with or without early maternal separation. Braz J Med Biol Res. 2002;35:1367-1371. [ Links ]
RAY J, HANSEN S. Temperament in the rat: Sex differences and hormonal influences on harm avoidance and novelty seeking. Behavi Neurosci. 2004;118:488-497. [ Links ]
RODGERS RJ, JOHNSON NJ. Factor analysis of spatiotemporal and ethological measures in the murine elevated plus-maze test of anxiety. Pharmacol Biochem Behav. 1995 Oct;52(2):297-303 [ Links ]
SHALEV U, KAFKAFI N. Repeated maternal separation does not alter sucrosereinforced and open-field behaviors. Behavioral Neuroscience Branch. 2002;73(1):115-122. [ Links ]
SLOTTEN HA, KALINICHEV M, HAGAN JJ, MARSDEN CA, FONE KC. Longlasting changes in behavioural and neuroendocrine indices in the rat following neonatal maternal separation: Gender-dependent effects. Brain Res. 2006;1097:123-32. [ Links ]
SULLIVAN RM, HOFER MA, BRAKE SC. Olfactory-guided orientation in neonatal rats is enhanced by a conditional change in behavior state. Dev Psychobiol. 1986;19:615-623. [ Links ]
ULRICH-LAI Y, HERMAN J. Neural regulation of endocrine and autonomic stress responses. Nat Revi Neurosci. 2009;10:397-409. [ Links ]

