










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.17 no.3 Bogotá Sept./Dec. 2012
DESARROLLO DE UN POXVIRUS RECOMBINANTE QUE EXPRESA LA GLICOPROTEÍNA D DEL HERPESVIRUS BOVINO-1
Development of a Recombinant Poxvirus Expressing Bovine Herpesvirus-1 Glycoprotein D
JULIÁN RUIZ SÁENZ1,2, Ph. D.; JORGE E. OSORIO3, Ph. D.; VÍCTOR J. VERA1, Ph. D.
1Grupo de Microbiología y Epidemiología. Universidad Nacional de Colombia, Bogotá, Colombia.
2Grupo de Investigación Centauro, Facultad de Ciencias Agrarias, Universidad de Antioquia. A.A. 1226, Medellín, Colombia.
3Department of Pathobiological Sciences, School ofVeterinary Medicine, University of Wisconsin, Madison, USA. Correspondencia: Julián Ruiz Sáenz. Carrera 75 # 65-87, bloque 47- 122. Medellín, Colombia. Tel.: 574-219 91 30. Fax: 574-219 91 60. julianruizsaenz@gmail.com
Presentado el 18 de enero de 2012, aceptado el 7 de septiembre de 2012, correcciones el 11 de septiembre de 2012.
RESUMEN
El herpesvirus bovino-1 (BHV-1) es un virus de genoma DNA perteneciente a la familia Herpesviridae, el cual afecta al bovino en el que provoca un amplio espectro de manifestaciones clínicas y pérdidas económicas. El principal componente inmunogénico de su envoltura es la glicoproteína D (gD), la cual ha sido caracterizada y utilizada como inmunógeno en distintos sistemas de expresión. El objetivo de este trabajo fue generar un poxvirus recombinante (Raccoonpox [RCN]) que expresara una versión truncada de la gD del BHV-1 para ser usado como inmunógeno. Para ello, se amplificó el gen que codifica para la versión truncada de la gD, la cual se clonó en el plásmido de transferencia pTK/ IRES/tpa que posee sitios de homología a la timidinakinasa del poxvirus, un sitio interno de entrada al ribosoma (IRES) y una señal secretoria (tPA), generando el constructo pTK/gD/IRES/tpa. Para generar el RCN recombinante, se tomaron células BSC-1, se infectaron con una cepa Silvestre del RCN (CDC/V71-I-85A) a un índice de multiplicidad de infección de 0,05 y se transfectaron con el constructo pTK/gD/IRES/tpa; generándose diferentes poblaciones virales con y sin el gen de interés. Para seleccionar los virus recombinantes que expresaban el gen de interés, se realizó una selección de recombinantes negativos para timidina kinasa y positivos para la gD por tres rondas de purificación de placas en monocapas de células RAT-2 las cuales son mutantes para timidina kinasa y en presencia de bromodeoxiuridina. Los virus recombinantes se confirmaron por PCR y secuenciación de nucleótidos y se denominaron RCN-gD.
Palabras clave: bovino, clonación molecular, rinotraqueitis, vacunas, virus, vectores genéticos.
ABSTRACT
Bovine Herpesvirus-1 is a DNA virus belonging to the family Herpesviridae, which affects cattle, causing a wide spectrum of clinical manifestations and economic losses. The main immunogenic component is its envelope glycoprotein D (gD), which has been characterized and used as immunogen in different expression systems. The aim of this work was to generate a recombinant poxvirus (Raccoonpox [RCN]) expressing a truncated version of BHV-1 gD to be used as a vaccine. To do this, it was amplified the gene for a truncated version of gD which subsequently was cloned in transfer plasmid PTK/IRES/tpa which has homology to sites of poxvirus thymidine kinase, an internal site of ribosome entry (IRES) and a secretory signal (tPA), generating the construct PTK/gD/IRES/tpa. To generate the recombinant RCN, we took BSC-1 cells and we infected with a wild type RCN (CDC/V71-I-85A) at a multiplicity of infection of 0.05, then cells were transfected with the construct PTK/gD/IRES/tpa, generating different viral populations with and without the gene of interest. To select recombinant viruses expressing the gene of interest, we performed a selection of recombinant thymidine kinase negative and positive for gD by three rounds of plaque purification on RAT-2 cells monolayers which are thymidine kinase null and using bromodeoxyuridine. Recombinant viruses were recovered and confirmed by PCR and nucleotide sequencing and so called RCN-gD.
Keywords: Cattle, molecular cloning, rhinotracheitis, genetic vectors, vaccines, virus.INTRODUCCIÓN
El herpesvirus bovino es un virus de genoma DNA perteneciente a la familia Herpesviridae, subfamilia Alfaherpesvinae (Wyler et al., 1989); el cual posee una amplia diversidad de cepas, las cuales son serológicamente indiferenciables. Sin embargo, con base en el análisis del genoma con endonucleasas de restricción y sus respectivos patrones de migración, se han clasificado en tres subtipos: BHV-1.1, BHV-1.2a, BHV-1.2b (Wyler et al., 1995). Los subtipos anteriormente conocidos como BHV-1.3a y BHV-1.3b han sido reclasificados como BHV-5, el cual presenta un alto potencial neuropatogénico (Büchen, 1995).
El BHV-1 afecta naturalmente al bovino, especie en la que provoca un amplio espectro de manifestaciones conocidas como rinotraqueitis; vulvovaginitis pustular infecciosa, balanopostitis, conjuntivitis, aborto, enteritis y encefalitis. La distribución de la infección es mundial y la prevalencia de reactores serológicos indica que prácticamente todos los bovinos mayores de tres años han estado en contacto con el virus (Pidone et al., 1999). En Colombia, se han realizado diferentes estudios seroepidemiológicos, los cuales además de demostrar la extensión de la infección, han permitido el aislamiento de algunas cepas nativas, las cuales fueron aisladas a comienzos de los años 70 (CIAT, 1972; CIAT, 1975), en la mitad de los años noventas (Molano y Rodríguez, 1995), y un grupo mayor (18 aislamientos) a finales de la última década, las cuales han sido sujeto de caracterización molecular, clasificando la mayoría de los aislamientos del país como BHV-1.1 y a un aislamiento de la Sabana de Bogotá como BHV-1.2a (Ruiz-Saenz et al., 2010; Ruiz- Saenz et al., 2012).Desde hace más de 20 años, existen en el comercio vacunas convencionales contra el BHV-1, las cuales se utilizan en muchos países en sus distintas variantes: generadas a partir de virus vivo modificado o inactivado, mono o polivalente, etc. Según la Organización Mundial de Sanidad Animal-OIE, 2004, dependiendo de la capacidad del producto de inducir inmunogenicidad, la vacuna puede ser efectiva en reducir las manifestaciones clínicas de la enfermedad y en consecuencia, reducir las pérdidas económicas asociadas a dicho proceso clínico; sin embargo, no logra proteger completamente de la infección por virus de campo.
Recientes avances en el desarrollo de vacunas han resultado en el desarrollo e implementación de vacunas más seguras y eficientes contra enfermedades humanas y veterinarias usando subunidades proteicas recombinantes, vectores virales y bacterianos, y vectores basados en DNA. Los problemas concernientes tanto a las vacunas atenuadas como a las vacunas inactivadas (reversión de virulencia, fallas en la inducción de inmunidad, etc.), impulsaron la producción de nuevas alternativas vacunales recombinantes (tanto replicativas como no replicativas) que contienen solo determinados componentes virales, los cuales han mostrado ser altamente inmunogénicos y en algunos casos permitir la diferenciación de animales vacunados de animales infectados en campo (Ruiz Saenz et al., 2009).
Para el BHV-1 se han descrito el desarrollo de diferentes vectores virales los cuales expresan glicoproteínas inmunogénicas, principalmente la gD. Entre estos vectores virales se resalta el uso del Herpesvirus Bovino-4 (BHV-4), un miembro de la subfamilia Gammaherpesvirinae, el cual expresando la gD del BHV-1 es altamente inmunogénico e induce la producción de anticuerpos neutralizantes anti-gD (Donofrio et al., 2006; Donofrio et al., 2008). Otra alternativa ampliamente explorada ha sido el uso de vectores adenovirales tanto de origen bovino como humano, y en versiones replicativas y no replicativas, encontrándose resultados contradictorios en los diferentes estudios (van Drunen Littel et al., 1993; Mittal et al., 1996; Reddy et al., 1999; Reddy et al., 2000; Gogev et al., 2002). Adicionalmente, se han desarrollado Baculovirus recombinantes como inmunógenos que llevan en su superficie moléculas quiméricas entre la glicoproteína de superficie gp64 del baculovirus y la gD derivada del BHV-1, lográndose inducir la producción de anticuerpos capaces de neutralizar el virus in vitro (Peralta et al., 2006; Peralta, 2007).
El poxvirus de mapaches (RaccoonPoxvirus - RCN) es un virus envuelto, de genoma DNA de doble cadena perteneciente a la Familia Poxviridae, Subfamilia Chordopoxvirinae, género Orthopoxvirus, nativo de América del Norte el cual fue aislado del tracto respiratorio superior de mapaches clínicamente sanos durante una evaluación de vigilancia epidemiológica en Maryland, Estados Unidos (Herman, 1964). Durante más de dos décadas, los RCN han sido usados como sistemas de expresión de genes heterólogos, demostrándose que la inserción de dichos genes heterólogos sobre el gen de la timidina kinasa viral (tk) reduce la virulencia del RCN a la vez que facilita la expresión del gen de interés (Esposito et al., 1988). El uso eficiente de este virus recombinante ha sido demostrado en diversas especies de mamíferos, entre ellos, zorros, conejos, gatos domésticos, lechones, ovejas y primates no humanos (Esposito et al., 1992). Distintas rutas de inoculación han sido evaluadas, demostrando no solo una amplia versatilidad de este vector para infectar diferentes células blanco en los organismos, sino también, una eficiente producción de inmunidad tanto celular como humoral contra enfermedades virales y bacterianas (Osorio et al., 2003; Memcher et al., 2004).
Teniendo en cuenta que los últimos estudios del BHV-1 realizados en el país demuestran que la prevalencia serológica llega a ser del 100 % por hato y más del 75 % por animales, tanto en explotaciones lecheras como en ganaderías de producción de carne (Ruiz- Saenz et al.,2010), y el hecho de la falta de una vacuna monovalente que induzca una eficiente respuesta inmune contra el BHV-1 y que nos permita diferenciar los animales vacunados de los animales expuestos naturalmente a la infección, el presente trabajo tuvo por objetivo generar un poxvirus recombinante (Raccoonpox [RCN]) que expresara una versión truncada de la gD del BHV-1 para ser usado a futuro como inmunógeno en bovinos.
MATERIALES Y MÉTODOS
CÉLULAS Y VIRUS
En el presente trabajo se utilizaron células de riñón de mono verde africano (BSC-1(ATCC #CCL-26)), células VERO (ATCC#CCL-18), fibroblastos de embrión de rata (RAT-2 (ATCC 1764)) y células MDBK (ATCC CCL-22) obtenidas del repositorio de células del Departamento de Ciencias Patobiológicas de la Facultad de Medicina Veterinaria de la Universidad de Wisconsin. Las células se mantuvieron a 37 °C en una atmósfera del 5 % de CO2 en DMEM (Gibco-Invitrogen, Inc. Grand Island, NY) suplementado con SFB al 10 % (Gibco-Invitrogen, Inc. Grand Island, NY), penicilina- estreptomicina (10.000 U/mL y 10 µ g/mL respectivamente) y anfotericina (0.25 µg/mL) (Gibco-Invitrogen, Inc. Grand Island, NY). Para los experimentos de transfección se usó OptiMEM® (Gibco-Invitrogen, Inc. Grand Island, NY) sin suplementación adicional de suero. El poxvirus de mapaches de referencia, RCN cepa CDC/V71-I-85A fue replicado en células VERO, mientras que el BHV-1 cepa IOWA fue replicado en células MDBK y almacenado a -80 °C. El RCN se mezcló 1:1 con solución de tripsina-versene (0,05 % de tripsina, EDTA 0,02 % en solución salina equilibrada) y se incubó durante 15 min a 37 °C para liberar partículas infecciosas de los agregados que puedan haberse formado durante el almacenamiento antes de la inoculación en células.
PLAQUEO Y TITULACIÓN POR UNIDADES FORMADORAS DE PLACA (UFP)
Se sembraron células VERO en platos de 24 pozos y fueron tratadas con diluciones decimales de los virus; una hora post-infección, se retiró el inóculo y se adicionó medio semisólido (MEM al 5 % de SFB con agarosa al 0,4 %) y se incubó por 72 horas a 37 °C en una atmósfera al 5 % de CO2, tiempo para el cual las monocapasfueronfijadas con formaldehido al 4 % y teñidas usando el colorante cristal violeta.
EXTRACCIÓN DEL DNA
Las muestras de virus tomadas a partir de cultivos celulares infectados fueron resuspendidas en 0,5 mL de buffer de lisis (10 mM Tris-HCl, pH 8,0; 100 mM EDTA; 0,5 % SDS) e incubadas a 50 °C en presencia de 20 g/mL de proteinasa K (Fermentas®), durante 3 horas. El ADN se extrajo del lisado celular usando fenol: cloroformo: alcohol isoamílico (25:24:1), seguido por precipitación con 50 µl de acetato de sodio (3M) y 1 mL isopropanol 100% (previamente conservado a -20 °C). Inmediatamente, el DNA fue recolectado por centrifugación y lavado dos veces con etanol al 70 % y disuelto en 200 µl de buffer TE (Tris 10 mM, EDTA 1mM).
PLÁSMIDO VECTOR DE TRANSFERENCIA
Se utilizó el vector pTK-IRES-tPA, el cual posee el sitio de inserción del DNA exógeno flanqueado por secuencias del gen tk del RCN y se pueden recombinar en el gen tk de los ortopoxvirus de tipo silvestre. Adicionalmente, el vector posee un potenciador de la traducción que es el sitio interno de entrada ribosomal (IRES) del virus de la encefalomiocarditis y la señal secretoria de la proteína activadora del plasminógeno tisular (tPA), los cuales es bien sabido que mejoran de forma significativa los niveles de expresión de genes exógenos en un RCN recombinante (Osorio et al., 2003).
AMPLIFICACIÓN Y CLONACIÓN DE LA VERSIÓN TRUNCADA DE LA GD DEL BHV-1
Para obtener la versión truncada de la gD del BHV-1, se realizó PCR con los cebadores: gDF 5'-TATTGGATCCATGCAAGGGCCGACATTGGCCGTG-3' y gDR 5'-TTACGAATTCTTAGGGCGTAGCGGGGGCGGGCG-3' y posterior restricción con las enzimas con BamHI y EcoRI, generando un fragmento de 1,089 pb aproximadamente. Este fragmento de PCR fue clonado en el plásmido de transferencia pTK-IRES-tPA previamente digerido con EcoRI y BamHI generando el plásmido pTK-IRES-tPA-gD, el cual tiene un tamaño de cerca de 5082 pb, el cual codifica para versión truncada de la gD de 355 aminoácidos. La selección del plásmido se realizó por resistencia a Ampicilina (GIBCO BRL) y la confirmación de la correcta inserción del transgen por secuenciación de nucleótidos y digestión con la enzima.
GENERACIÓN DEL POXVIRUS RECOMBINANTE
Los virus recombinantes se obtuvieron utilizando el sistema de recombinación de secuencias homólogas in vitro. Brevemente, para la generación del RCNgD, se sembraron platos de seis pozos con 500.000 células de BSC-1, las cuales posteriormente se infectaron con el RCN cepa de referencia (CDC/V71-I-85A) a un Índice de Multiplicidad de Infección (MOI) de 0,05; 90 minutos posinoculación, las células fueron transfectadas con el vector de transferencia pTK-IRES-tpa-gD usando el sistema FuGENE ® 6 (Roche®) y se incubó a 37 °C durante 72 horas. Posterior a la infección/transfección, las células se congelan y descongelado tres veces y el lisado fue inoculado en células RAT-2 en medio M199 con 5 % de SFB y 50 ug/mL 5-bromo-2-desoxiuridina (BrdUsigma®). Tras una incubación de dos horas, se retiró el medio y la monocapa infectada se cubrió con MEM con 5 % de SFB y 1,2 % de agarosa de bajo punto de fusión. En presencia de BrdU, solo los virus RCN tk- dan lugar a la formación de UFP. Las placas de crecimiento viral resistentes a la BrdU fueron recolectadas aproximadamente 14 días después de la inoculación (PI) y se replaquearon dos veces en células RAT-2 (mutantes de timidina kinasa) cultivadas en un medio con BrdU. Los clones virales positivos se amplificaron y caracterizaron para su posterior crecimiento a gran escala en las células Vero. En la figura 1 se presenta una representación gráfica resumida del sistema tradicional de clonación del gen de interés y posterior transfección/infección en células de mamífero (Byrd y Hruby, 2004).
CONFIRMACIÓN DE LOS RECOMBINANTES
Se realizó extracción de ADN, PCR y secuenciación de nucleótidos utilizando los cebadores: RCNgDF 5'- ATGCAAGGGCCGACATTGGCCGTG-3' y RCNgDR 5'-TTAGGGCGTAGCGGGGGCGGGCG-3', para confirmar la secuencia de la gD expresada por el RCN-gD.
Las condiciones de PCR fueron un minuto de desnaturalización y activación de polimerasa en 95 °C, seguido por 30 ciclos de desnaturalización a 95 °C durante 30 segundos, 62° C durante 30 segundos, y 72 °C durante 30 segundos con una extensión final de 10 minutos a 72 °C. Los productos de PCR se verificaron en gel de agarosa al 0,8 % (p/v) preparado en buffer TAE y teñido con bromuro de etidio.
RESULTADOS Y DISCUSIÓN
Se replicó el BVH-1 en células MDBK durante 48 horas para posteriormente realizar extracción de DNA viral y PCR usando los cebadores descritos anteriormente a partir del sobrenadante del cultivo con efecto citopático característico. Se amplificó un fragmento de 1,089 pb que codifica para una versión truncada de la glicoproteína D del BHV-1 la cual posee un codón de terminación en el aminoácido 355, corriente arriba del dominio de anclaje de membrana (van Drunen littel van Den Hurk et al., 1990). Se ha demostrado que dicha proteína es altamente inmunogénica en bovinos, permitiendo la producción de anticuerpos neutralizantes y de inmunidad celular, humoral y de mucosas (van Drunen Littel van den Hurk et al., 1993, van Drunen Littel van den Hurk et al.,1994; van Drunen Littel van den Hurk, 2006).
Para el proceso de clonación, se utilizó el plásmido de referencia pTK-IRES-tPA, en el cual se realizó la ligación del gen de la gD amplificada para su posterior selección por resistencia ampicilina generando el plásmido pTK-IRES-tPA-gD (Fig. 2).
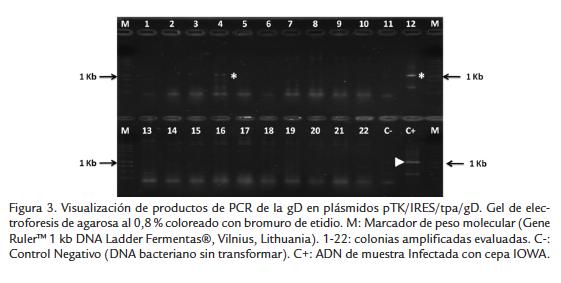
A partir del DNA extraído de las colonias amplificadas en mini preparación se realizó PCR para la gD del BHV-1 para confirmar la presencia de dicho gen en las colonias. Se realizó evaluación por PCR de aproximadamente 60 colonias para encontrar cinco colonias que mostraron la presencia del gen de interés (Fig. 3). Dichas colonia se amplificaron en una maxi-preparación de la cual se obtuvo un lote de bacterias transformadas que se almacenó posteriormente a -80°C. Los plásmidos fueron secuenciados para confirmar la presencia del gen de la gD y se realizó digestión con la enzima XmaI para confirmar la correcta inserción.
Una vez confirmada la presencia del gen de interés en el plásmido de transferencia, este se usó para realizar el ensayo de infección/transfección usando 1 µg de DNA plasmídico por pozo de células BSC-1, las cuales posteriormente fueron infectadas con el RCN a MOI de 0,05 y se incubó durante 72 horas a 37 °C, tiempo para el cual hubo presencia de efecto citopático (Figs. 4a, 4c). Posteriormente, las células fueron recuperadas, lisadas y el sobrenadante puesto sobre una monocapa confluente de células Rat-2 en presencia de medio semisólido y BrdU, para posteriormente ser llevadas a incubación durante 14 días tiempo para el cual solo los virus recombinantes TK- logran replicarse y dar lugar a la formación de placas de lisis (Figs. 4d, 4e).
Cada placa de lisis fue replicada en una monocapa nueva en presencia de medio con BrdU realizándose dos rondas de plaqueo y confirmando por PCR la presencia del gen de la gD en los virus RCNgD (Fig. 5)
Los poxvirus han sido ampliamente utilizados como inmunógenos, incluso algunos ya han sido licenciados para uso comercial en diversas partes de Estados Unidos y Europa para su uso en diversas especies de interés veterinario (Larson y Schultz, 2007; Guthrie et al., 2009; Soboll et al., 2010); incluso en Colombia, recientemente se autorizó la comercialización de un Canarypox que expresa las glicoproteínas H y F del virus del Distemper canino (ICA, Reg 4734 y 4735 de 2011). En general, los poxvirus han demostrado ser muy buenos inmunógenos y dado que no tienen potencial patogénico en una especie diferente a su especie huésped natural son uno de las más importantes alternativas para vacunación con virus con capacidad replicativa en el mundo (Vanderplasschen y Pastoret, 2003). Puntualmente, el RCN ha sido empleado como un vector viral que expresa proteínas de Yersinia pestis (Rocke et al., 2008; Rocke et al., 2009), como un vector que expresa eficientemente la glicoproteína G del virus de la Rabia (Osorio et al., 2003) y más recientemente como un vector viral que expresa la Hemaglutinina (H5) del virus de la influenza aviar de alta patogenicidad (H5N1) (Hwa et al., 2010), demostrando en todos los casos ser un eficiente sistema para la inducción de inmunidad humoral, inmunidad en mucosas e inmunidad celular (Walsh y Dolin, 2011).
Un constructo similar basado en un virus de Vaccinia Ankara -MVA expresando la misma versión de esta glicoproteína del BHV-1, ha sido generado y evaluado en conejos y ratones modelos de infección por el BHV-1, encontrando este recombinante muy útil para inducir inmunidad humoral específica y reducir los niveles de excreción viral posteriores al reto con BHV-1 de referencia (Ferrer et al., 2011). Es importante resaltar que el RCNgD generado en el presente trabajo podría tener mejores propiedades inmunogénicas que el MVA-gDs desarrollado por Ferrer et al., en 2011, dado que el RCNgD contiene IRES del virus de la encefalomiocarditis y señal de secreción del activador de plasminógeno tisular (tPA) lo cual se ha visto en que mejora de forma significativa los niveles de expresión del transgen de interés (Osorio et al., 2003; Rocke, 2009). Teniendo en cuenta que ninguno de estos vectores poxvirales ha sido explorado en bovinos, resaltamos la importancia de explorar la capacidad inmunogénica del RCNgD recombinante generado en el presente trabajo y su potencial uso como vector en bovinos tanto para la gD del BHV-1 como para otras proteínas inmunogénicas de agentes patógenos de importancia económica en el campo médico veterinario.
Por otra parte, la seguridad para el personal que aplica la vacuna o que se expone al RCN recombinante ha sido probada, mostrando que no existe riesgo significativo ante la exposición directa (Rocke et al., 2004) e incluso dada la nula patogenicidad tanto en diferentes especies de animales como en humanos, se han realizado estudios que proponen su uso como un vector seguro y eficaz tanto para generar inmunidad como para tratar algunos tipos de cáncer (Evgin et al., 2010).
CONCLUSIONES
En el presenta trabajo se presenta el desarrollo técnico de un poxvirus de mapaches que expresa la gD del BHV-1 (RCNgD), un virus que tiene amplio potencial para ser desarrollado a escala como una potencial vacuna monovalente contra la infección por el BHV-1. Es necesario por tanto realizar estudios de in vitro de expresión y estabilidad de la gD recombinante en el vector y llevar dicho recombinante a pruebas in vivo de inducción de inmunidad y de protección ante el reto con cepas de referencia.
AGRADECIMIENTOS
Los autores quieren agradecer al personal del laboratorio del Dr. Jorge Osorio en la Universidad de Wisconsin, Estados Unidos, por el apoyo técnico en el desarrollo del proyecto. JRS es becario del programa de apoyo a doctorandos nacionales de Colciencias. Este trabajo fue financiado por la División de Investigación Universidad Nacional de Colombia, proyectos n.°: 20201007738 y 202010013254.
BIBLIOGRAFÍA
BÜCHEN OSMOND C. Alphabetical list of ICTV approved virus families and genera. Creado Abril, 1995. Última actualización: 8 de abril de 1998. [citado noviembre de 2011]. Disponible en: URL:http://ncbi.nlm.nih.gov/ICTVdb/ICTVdB [ Links ]
BYRD CM, HRUBY DE. Construction of recombinant vaccinia virus: cloning into the thymidine kinase locus. Methods Mol Biol. 2004;269:31-40. [ Links ]
CIAT. Centro Internacional de Agricultura Tropical. Informe anual de salud animal. Bogotá; 1972. p. 447. [ Links ]
CIAT. Centro Internacional de Agricultura Tropical. Informe anual de salud animal. Cali; 1975. p. 63. [ Links ]
DONOFRIO G, CAVIRANI S, VANDERPLASSCHEN A, GILLET L, FLAMMINI CF. Recombinant bovine herpesvirus 4 (BoHV-4) expressing glycoprotein D of BoHV-1 is immunogenic and elicits serum-neutralizing antibodies against BoHV-1 in a rabbit model. Clin. Vaccine Immunol. 2006;13(11):1246-1254. [ Links ]
DONOFRIO G, SARTORI C, FRANCESCHI V, CAPOCEFALO A, CAVIRANI S, TADDEI S, et al. Double immunization strategy with a BoHV-4-vectorialized secreted chimeric peptide BVDV-E2/BoHV-1-gD. Vaccine. 2008;26(48):6031-6042. [ Links ]
EVGIN L, VÄHÄ-KOSKELA M, RINTOUL J, FALLS T, LE BOEUF F, BARRETT JW, et al. Potent oncolytic activity of raccoonpox virus in the absence of natural patho- genicity. Mol Ther. 2010;18(5):896-902. [ Links ]
ESPOSITO JJ, KNIGHT JC, SHADDOCK JH, NOVEMBRE FJ, BAER GM. Successful oral rabies vaccination of raccoons with raccoon poxvirus recombinants expressing rabies virus glycoprotein. Virology. 1988;165(1):313-316. [ Links ]
ESPOSITO JJ, SUMNER JW, BROWN DR, EBERT JW, SHADDOCK JH, BAI XH, et al. Raccoon poxvirus rabies-glycoprotein recombinant oral vaccine for wildlife: further efficacy and safety studies and serosurvey for raccoon poxvirus. In: Brown F, Chanock RM, Ginsberg H, Lerner RA (eds), Vaccines-91: modern approaches to new vaccines including prevention of AIDS. Cold Spring Harbor Laboratory. New York; 1992. p. 321-330. [ Links ]
FERRER MF, DEL MÉDICO ZAJAC MP, ZANETTI FA, VALERA AR, ZABAL O, CALAMANTE G. Recombinant MVA expressing secreted glycoprotein D of BoHV-1 induces systemic and mucosal immunity in animal models. Viral Immunol. 2011;24(4):331-339. [ Links ]
GOGEV S, VANDERHEIJDEN N, LEMAIRE M, SCHYNTS F, D'OFFAY J, DEPREZ I, et al. Induction of protective immunity to bovine herpesvirus type 1 in cattle by intranasal administration of replication-defective human adenovirus type 5 expressing glycoprotein gC or gD. Vaccine. 2002;20:1451-1465. [ Links ]
GUTHRIE AJ, QUAN M, LOURENS CW, AUDONNET JC, MINKE JM, YAO J, et al. Protective immunization of horses with a recombinant canarypox virus vectored vaccine co-expressing genes encoding the outer capsid proteins of African horse sickness virus. Vaccine. 2009;27(33):4434-4438. [ Links ]
HWA SH, IAMS KP, HALL JS, KINGSTAD BA, OSORIO JE. Characterization of recombinant raccoonpox vaccine vectors in chickens. Avian Dis. 2010;54(4):1157-1165. [ Links ]
IOANNOU XP, GRIEBEL P, HECKER R, BABIUK LA, VAN DRUNEN LITTEL-VAN DEN HURK S. The immunogenicity and protective efficacy of bovine herpesvirus 1 glycoprotein D plus Emulsigen are increased by formulation with CpGoligodeoxy- nucleotides. J Virol. 2002;76:9002-9010. [ Links ]
LARSON LJ, SCHULTZ RD. Three-year serologic immunity against canine par- vovirus type 2 and canine adenovirus type 2 in dogs vaccinated with a canine com- bination vaccine. Vet Ther. 2007;8(4):305-310. [ Links ]
MENCHER JS, SMITH SR, POWELL TD, STINCHCOMB DT, OSORIO JE, ROCKE TE. Protection of black-tailed prairie dogs (Cynomysludovicianus) against plague after voluntary consumption of baits containing recombinant raccoon poxvirus vaccine. Infect Immun. 2004;72(9):5502-5505. [ Links ]
MITTAL SK, PAPP Z, TIKOO SK, BACA-ESTRADA ME, YOO D, BENKO M, et al. Induction of systemic and mucosal immune responses in cotton rats immunized with human adenovirus type 5 recombinants expressing the full and truncated forms of bovine herpesvirus type 1 glycoprotein gD. Virology. 1996;222(2):299-309. [ Links ]
MOLANO D, RODRÍGUEZ JL, RAMÍREZ G, VILLAMIL LC. Caracterización electro- forética e inmunológica de una cepa de campo del virus de la rinotraqueitis infecciosa bovina y su comparación con cepas de referencia. Rev Med Vet Zootec. 1996;44(1):35-38. [ Links ]
ORGANIZACIÓN MUNDIAL DE SANIDAD ANIMAL (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 5a edición. OIE. París; 2004. p. 894-903. [ Links ]
OSORIO JE, FRANK RS, MOSS K, TARASKA T, POWELL T, STINCHCOMB DT. Raccoon poxvirus as a mucosal vaccine vector for domestic cats. J Drug Target. 2003;11(8-10):463-470. [ Links ]
OSORIO JE, POWELL TD, FRANK RS, MOSS K, HAANES EJ, SMITH SR, et al. Recombinant raccoon pox vaccine protects mice against lethal plague. Vaccine. 2003;21(11-12):1232-1238. [ Links ]
PERALTA A, MOLINARI MP, TABOGA O. Baculovirus recombinantes como inmunogenos. Rev Argent Microbiol. 2006;38(2):74. [ Links ]
PERALTA A, MOLINARI P, CONTE-GRAND D, CALAMANTE G, TABOGA O. A chimeric baculovirus displaying bovine herpesvirus-1 (BHV-1) glycoprotein D on its surface and their immunological properties. Appl Microbiol Biotechnol. 2007;75(2):407-414. [ Links ]
PIDONE CL, GALOSI CM, ETCHEVERRIGARAY EM. Herpesvirus Bovinos 1 y 5. Analecta Veterinaria. 1999;19(1/2):40-50 [ Links ]
REDDY PS, IDAMAKANTI N, PYNE C, ZAKHARTCHOUK AN, GODSON DL, PAPP Z, et al. The immunogenicity and efficacy of replication-defective and replication- competent bovine adenovirus-3 expressing bovine herpesvirus-1 glycoprotein gD in cattle. Vet Immunol Immunopathol. 2000;76(3-4):257-268. [ Links ]
REDDY PS, IDAMAKANTI N, ZAKHARTCHOUK AN, BAXI MK, LEE JB, PYNE C, et al. Nucleotide sequence, genome organization and transcription map of bovine adenovirus type 3. J Virol. 1999;72(2):1394-1402. [ Links ]
ROCKE TE, DEIN FJ, FUCHSBERGER M, FOX BC, STINCHCOMB DT, OSORIO JE. Limited infection upon human exposure to a recombinant raccoon pox vaccine vector. Vaccine. 2004;22(21-22):2757-2760. [ Links ]
ROCKE TE, SMITH SR, STINCHCOMB DT, OSORIO JE. Immunization of black- tailed prairie dog against plague through consumption of vaccine-laden baits. J Wild Dis. 2008;44(4):930-937. [ Links ]
ROCKE TE, IAMS KP, DAWE S, SMITH SR, WILLIAMSON JL, HEISEY DM, et al. Further development of raccoon poxvirus-vectored vaccines against plague (Yersinia pestis).Vaccine. 2009;28(2):338-344. [ Links ]
RUIZ SAENZ J, JAIME J, RAMIREZ G, VERA VJ. Molecular and in vitro characterization of field isolates of bovine herpesvirus-1. Virol Sin. 2012;27(1):26-37. [ Links ]
RUIZ SAENZ J, JAIME J, VERA V. Serological prevalence and isolation of Bovine herpesvirus-1 (BHV-1) in cattle herds from Antioquia and Valle del Cauca provinces. Rev Colomb Cienc Pecu. 2010;23(3):299-307. [ Links ]
RUIZ SÁENZ J, JAIME J, VERA V. Vacunas contra el BHV-1: Una mirada desde el pasado hacia el futuro de la inmunización. Acta biol Colomb. 2009;14(2):3-20 [ Links ]
SOBOLL G, HUSSEY SB, MINKE JM, LANDOLT GA, HUNTER JS, JAGANNATHA S, et al. Onset and duration of immunity to equine influenza virus resulting from canarypox- vectored (ALVAC) vaccination. Vet Immunol Immunopathol. 2010;135(1-2):100-107. [ Links ]
VAN DRUNEN LITTEL-VAN DEN HURK S, PARKER MD, MASSIE B, VAN DEN HURK JV, HARLAND R, BABIUK LA. Protection of cattle from BHV-1 infection by immunization with recombinant glycoprotein gIV. Vaccine. 1993;11(1):25-35. [ Links ]
VAN DRUNENLITTEL-VAN DEN HURK S, GIFFORD GA, BABIUK LA. Epitope specificity of the protective immune response induced by individual bovine herpesvirus- 1 glycoproteins. Vaccine. 1990;8(4):358-368. [ Links ]
VAN DRUNENLITTEL-VAN DEN HURK S, PARKER MD, MASSIE B, VAN DEN HURK JV, HARLAND R, BABIUK LA, et al. Protection of cattle from BHV-1 infection by immunization with recombinant glycoprotein gIV. Vaccine. 1993;11(1):25-35. [ Links ]
VAN DRUNENLITTEL-VAN DEN HURK S, VAN DONKERSGOED J, KOWALSKI J, VAN DEN HURK JV, HARLAND R, BABIUK LA, et al. A subunit gIV vaccine, produced by transfected mammalian cells in culture, induces mucosal immunity against bovine herpesvirus-1 in cattle. Vaccine. 1994;12(14):1295-1302. [ Links ]
VAN DRUNENLITTEL-VAN DEN HURK S. Rationale and perspectives on the success of vaccination against bovine herpesvirus-1.Vet Microbiol. 2006;113(3-4):275-282. [ Links ]
VANDERPLASSCHEN A, PASTORET PP. The uses of poxviruses as vectors. Curr Gene Ther. 2003;3(6):583-595. [ Links ]
WALSH SR, DOLIN R. Vaccinia viruses: vaccines against smallpox and vectors against infectious diseases and tumors. Expert Rev Vaccines. 2011;10(8):1221-1240. [ Links ]
WYLER R, ENGELS M, SCHWYZER M. Infectious Bovine Rhinotracheitis/Vulvovaginitis (BHV-1) Institute of Virology. University of Zurich; 1995. p. 1-55. [ Links ]
